
Abstract
This note reports observations on the pollen collecting behavior and foraging activity of the solitary bee, Anthophora walteri Gonzalez on Salvia bogotensis in the Eastern Andes of Colombia. Bees foraged from 7:00–17:00 h, or when the temperature exceeded 15°C. Peak visits occurred between 8:00-9:00, when the temperature was about 18°C and the humidity was 60%. On average, bees spent 3 seconds at each flower and collected pollen throughout the day, although pollen-collecting trips were twice as frequent in the morning as in the afternoon. The daily number and duration of foraging trips per bee ranged from 1-13 trips (x = 6.8±4.3) and 4-88 min (x = 21.7±23.8). Some possible morphological and behavioral adaptations for pollen collection on flowers of Salvia, as well as thermal constraints on the foraging activity of A. walteri in the Andes are also discussed.
Introduction
Anthophora (Mystacanthophora) walteri Gonzalez 2004 (Hymenoptera: Apidae, Anthophorini) is a solitary bee species, about the size of a honeybee worker (12– 15 mm long), endemic to isolated xeric highlands (2000–3000 m) in the Eastern Andes of Colombia (Gonzalez & Chavez 2004; Gonzalez & Engel 2004; Gonzalez et al. 2005). Nests are usually found in aggregations of 5–65 nests per square meter in flat ground or small banks in very hard soils in open, barren areas. As in other species of the subgenus Mystacanthophora, and certain bee genera in the families Andrenidae, Colletidae and Megachilidae, females of A. walteri have a face with a flat supraclypeal area and a clypeus covered with apically hooked hairs. Such modified hairs are apparently related to pollen collection from Salvia (Lamiaceae) (Michener 2000; Gonzalez & Chavez 2004). Palynological analysis of A. walteri brood provisions in Mondoñedo (Departamento of Cundinamarca, Colombia) showed that nearly all pollen within each nest cell was from Salvia bogotensis Bentham, 1824 (Gonzalez & Chavez 2004). Because little is known about the foraging ecology of A. walteri or any neotropical Anthophora, herein we report observations on its foraging and pollen-collecting behavior. Possible morphological and behavioral adaptations to pollen collection on flowers of Salvia, as well as the effect of climatic conditions on its foraging activity, are also discussed.
Materials and Methods
This study was conducted on December 18–20, 2004, in Mondoñedo (4° 39′ 52.9″ N; 74° 17’ 2″ W), about 10 km W of Bogotá city (Cundinamarca, Colombia), at 2720 m. Mondoñedo is a semiarid area that is highly disturbed by cattle ranching, gravel extraction and waste dumping. The rainy season is bimodal with a maximum in April-June and the other in September-October, never reaching above 100 mm of precipitation monthly. The median temperature is approximately 13 °C. The most predominant plant is Salvia bogotensis, a small shrub (< 1 m tall) that provides sparse but spatially aggregated shade in the area (Gonzalez & Chavez 2004).
The daily foraging activity of A. walteri was studied at a nest aggregation consisting of 21 nests per square meter, and on three patches (8–9 mβ) of S. bogotensis. Early in the morning, before bees started to forage, nests entrances were marked with small pieces of paper (~ 2x2 cm), which were attached to the ground with an insect pin. The departure and arrival times were recorded for each bee, as well as the presence or absence of pollen on the scopae. Pollen carrying females were easy to distinguish by the yellow pollen loads on their hind leg scopae. Unlike arrivals, bees left the nest very quickly and did not exhibit an orientation flight at the nest entrance; thus, it was difficult to observe the exact departure time in some cases. We therefore only used data on arrival times to build figure 3. Observations were made about 1.0–1.5 m away from the nest aggregation.
At the flowers, the numbers of A. walteri individuals were recorded every 30 minutes from 6:00 to 18:00 during a 10 min scan sampling (n = 2 h total) on Dec. 20, 2004. When possible, focal observations were taken from marked females (n = 34) that were uniquely painted on the mesosoma with quick-drying Decocolor ® paint markers 24 h before observations. Bees were captured on the selected patches with an insect net and placed into plastic vials and immobilized on ice for about 3–4 min. Once marked, bees were released on the same patch where they had been captured. Ambient temperature and relative humidity taken in shade were recorded using a digital hygrother-mometer (Extech®).
We used a linear regression analysis to test the effect of ambient temperature and relative humidity on the frequency of bee visits. Mean values are given with standard deviations.
Results
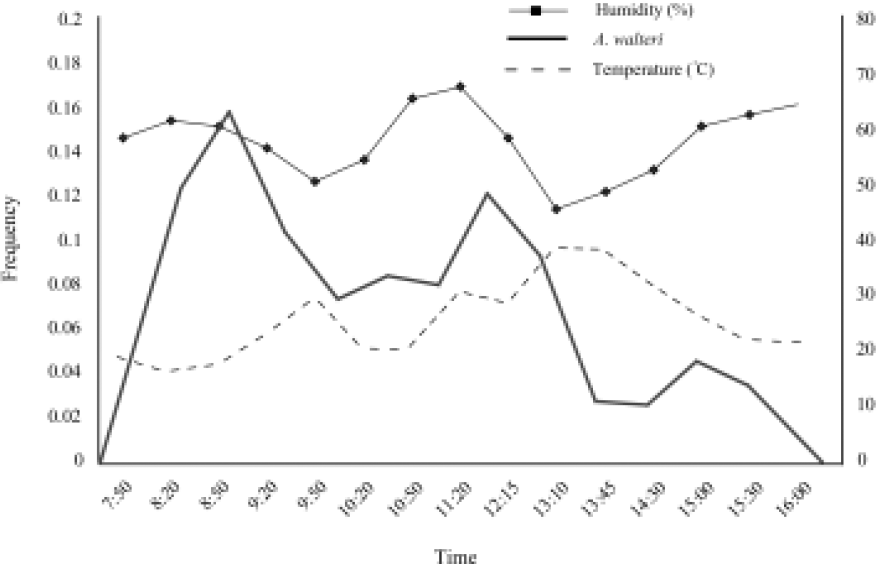
Foraging activity of Anthophora walteri on a flower patch of Salvia bogotensis in Mondoñedo (Colombia). Frequency of individuals observed is shown on left y-axis; temperature and humidity on right y-axis.
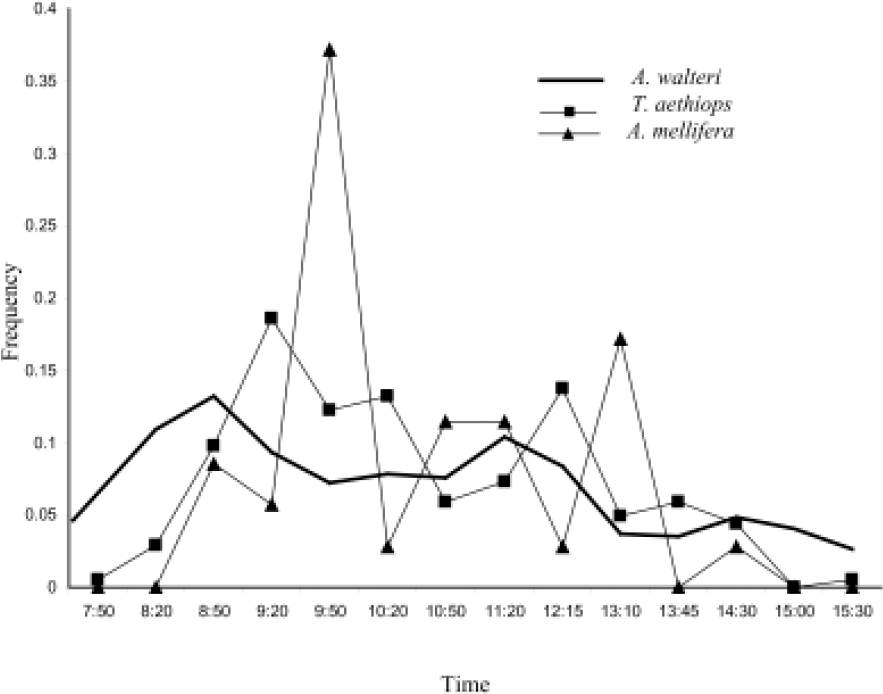
Comparative foraging activities of Anthophora walteri, Thygater aethiops, and Apis mellifera on flowers of Salvia bogotensis.
The daily number and duration of foraging trips per bee female ranged 1–13 trips per day (x = 6.8 ± 4.3, n = 10 bees) and 4–88 min per trip (x= 21.7 min ± 23.8, n = 24 trips). Bees collected pollen throughout the day, although the percentages of pollen-collecting trips were twice as frequent in the morning as in the afternoon (Fig. 3). Pollen-collecting trips were observed in 42 % of the total observed foraging trips (n = 125). Bees may have carried nectar or water in their crops during the remaining trips but we did not test this.
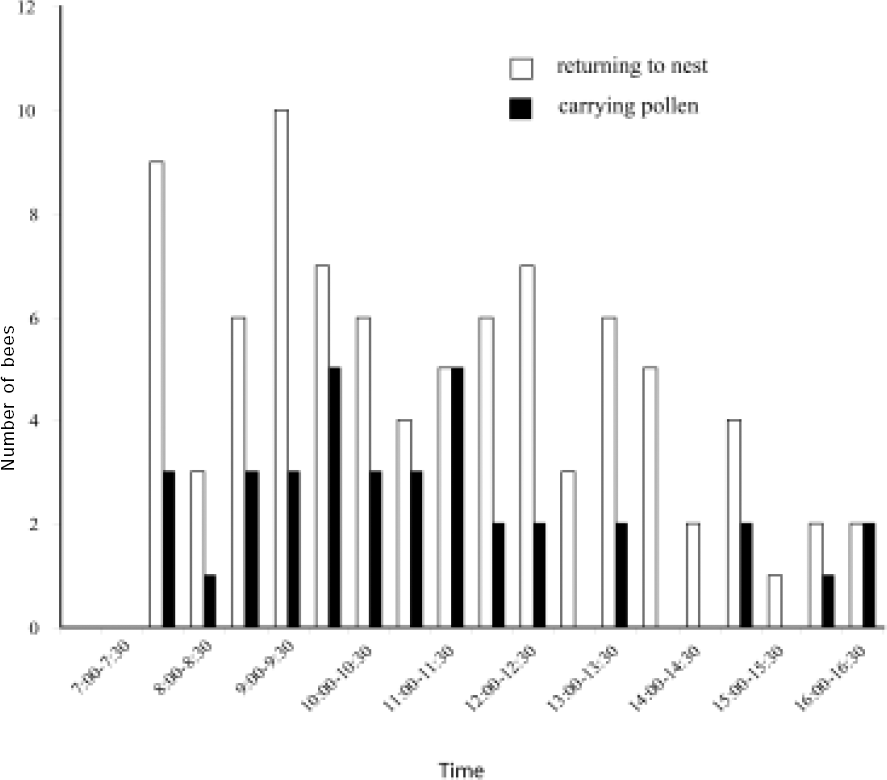
Number of Anthophora walteri females observed on Dec. 20, 2004, returning and carrying pollen of Salvia bogotensis to the nest.
Discussion
All bee species that visited S. bogotensis, including A. walteri, seemed to be attracted to flowers primarily for nectar as indicated by the extended tongue on approach to the flower. Although we do not have data, it is likely that pollen is accidentally transferred to the face in all species during a visit. However, the modified hairs on the faces of A. walteri female may increase the amount of pollen transferred. Most bees, especially hairy bees like B. atratus and T. aethiops, comb off the pollen from the face using the forelegs during grooming (Thorp 1979; 2000; Roubik 1989; Michener 2000) but the additional repertory of behaviors (i.e., moving the abdomen while holding onto a stem with the mandibles) were only observed in A. walteri. In addition, some of the marked females (n = 8 out of 34) returned several times during the day (1-4) to forage on the same patch where they had been captured. Anthophora walteri females also seemed to skip flowers that had previously been visited by other bees, suggesting they are capable of recognizing them. All these observations indicate morphological (modified hairs on face) and behavioral adaptations for collecting pollen from Salvia.
The genus Salvia comprises nearly 1000 species worldwide, and they are well known by their modified lever-like stamens, which play a key role in pollen transfer (Claßen-Bockhoff et al. 2003). A bee or bird (the main pollinators of Salvia) searching for nectar pushes one of the arms of the modified stamens that blocks the access to nectar, causing pollen to be loaded onto its head, bill or back. As a result, the pollen may be transferred to the stigma of another flower during a subsequent visit. This type of mechanism is often referred as nototribic pollination (Claßen-Bockhoff et al. 2004). Is A. walteri the only pollinator of S. bogotensis? We do not have data to answer this and related questions regarding pollen-transfer mechanisms involved in S. bogotensis. However, given that pollinator availability is very low for plants in high tropical altitudes in comparison with lower elevations due to climatic conditions (Primack 1985), specialization on a particular type of pollinator does not seem like a “best” strategy. Instead, even small contributions from a wide range of pollinators may be advantageous to high Andean plants such as S. bogotensis (Fagua & Gonzalez, in press). Undoubtedly, the pollination ecology of S. bogotensis needs to be studied in order to address such questions. Similarly, we do not know if A. walteri switches plant host when S. bogotensis is temporally unavailable as has been observed in others pollen specialist bees (Wcislo & Cane 1996).
Tropical high altitude ecosystems such as in Mondoñedo represent climatically hostile environments. Native species must tolerate drastic diurnal temperature changes, freezing temperatures during the night and avoid desiccation during the day (Sarmiento 1986; Lüttge 1997). Low temperatures and inclement weather strongly influence flight activity of heterothermic animals such as bees. Therefore, foraging is primarily restricted to warmer daily periods and to those animals that can efficiently thermoregulate their body temperature (Bishop & Armbruster 1999). We did not find a significant effect of the ambient temperature and relative humidity on the foraging activity of A. walteri though we only have data from a single day. Nonetheless, A. walteri did not fly at temperatures below 15 °C, during light rain or even before sunrise as frequently reported for some temperate Anthophora species [e.g., A. plumipes (Pallas, 1772), and A. bomboi-des stanfordiana Cockerell, 1904] (Brook 1983; Batra 1994; Stone 1994). This agrees with the idea that unlike arthropods from temperate areas, tropical alpine arthropods apparently do not have time for physiological preparation before the onset of low temperatures (Sømme 1989; Sømme et al. 1996). Furthermore, diurnal changes in quality and quantity of available floral nectar and pollen in S. bogotensis may also influence A. walteri activity as noted for other oligolectic Anthophora species (Stone et al. 1999).
In addition to climatic conditions, the number and duration of foraging trips per day of bees may also depend on individual status in social species, nest activities (cell construction, provisioning), type of material collected (mud, pollen, nectar), resource distance, etc. The daily number and length of trips observed in A. walteri are within the range of trips reported for other social and solitary species (e.g., Michener 1974). Spending time away from the nest while foraging for food could increase the risk of brood parasitization, especially in solitary species. Anthophora walteri females spent as much as 7 min foraging on 130 flowers before loading pollen in the scopae; that is, they could visit more than 500 flowers to complete a full pollen load on each trip, and spend on average, at least, 3.5 h per day (~200 min) away from the nest. Unlike habitats containing Anthophora species in the Northern Hemisphere, no nest parasites (including parasitic bee species) are known to occur in Mondoñedo (Gonzalez & Chavez 2004). Although we did not mark the bees from the nest aggregation under study, it was clear that sometimes a non-resident female entered a nest when it was already occupied. Buzzing and aggressive encounters usually occurred, and presumably those non-resident bees, then left the nest. Such intra-specific competition has also been reported in other Anthophora species (Batra 1994).
Footnotes
Acknowledgments
This paper is dedicated to Mr. Mauricio Palacios and Mrs. Rita Morillo, beloved parents of E. Palacios, for their encouragement, patient and unconditional support during our fieldwork in Mondoñedo. We thank Petra Wester, Daniel Bennett, Carlos Sarmiento, Marisol Amaya, and three anonymous reviewers for comments on the manuscript. Finally, we gratefully acknowledge the financial support of Idea Wild (to VHG). This is a contribution of the Division of Entomology, Natural History Museum and Biodiversity Research Center, University of Kansas.