
Abstract
It has been found that ant biodiversity is reduced with the transition from traditional coffee plantations (shade) to intensified (sun) systems and also that the composition of ant species changes in coffee plantations according to management. It has been hypothesized that ants from sun coffee plantations have physiological adaptations that permits them to colonize these habitats in comparison with ants from shade plantations. Using temperature as an indicator of microclimatic fluctuation, the behavior of Tetramorium simillimum, exclusive to sun plantations, and of Paratrechina steinheili, abundant but not exclusive to shade plantations (in Apía, Risaralda, Colombia), were analyzed in two thermal fluctuation tests between 17°C and 32°C, and one thermal shock test (12°C and 35°C) in an environmental chamber. Control colonies were maintained at constant temperature (24 +/- 1°C). Ant activity outside the nest, foraging and mortality were evaluated. In the first two tests P. steinheili exhibited a relatively lower mortality than T. simillimum. In both cases the colonies submitted to stress in the environmental chamber lost more workers than controls. In the thermal shock test an increase in P. steinheili mortality was found, but it did not exceed that of T. simillimum. It is concluded that there is no evidence for physiological adaptation, at least detectable, of T. simillimum to thermal stress in comparison to P. steinheili and that perhaps other ecological factors, such as limitation of nesting resources, may restrict the presence of the latter to sun coffee plantations.
Introducción
En diferentes países de Latinoamérica se ha demostrado que la transición de sistemas de cultivo tradicional de café, con sombrío de árboles acompañantes, a sistemas intensificados en donde se practica el monocultivo sin sombra, ejerce una drástica reducción de la diversidad de hormigas asociadas (Perfecto et al. 1997; Moguel y Toledo 1999; Armbrecht y Perfecto 2003; Rivera y Armbrecht 2005).
Para explicar por qué se afecta negativamente la biodiversidad asociada a los cultivos, al ser simplificados e industrializados, se han planteado como posibles causas la limitación en el suministro alimenticio, la disponibilidad y estabilidad de los sitios de anidamiento (Philpott y Foster 2005) y las fluctuaciones microclimáticas (Andersen 2000). Estas últimas se relacionan con la respuesta fisiológica que diferentes especies puedan tener a los cambios microclimáticos (Perfecto y Vandermeer 1996) al igual que a los procesos evolutivos que la adaptaron a estas condiciones de bosque. La vulnerabilidad a las condiciones se puede explicar en términos de temperatura y humedad del microambiente, ya que las hormigas especializadas de hojarasca son altamente susceptibles a la pérdida de agua (Kaspari 1996; Andersen 2000; Ramírez 1998). En este estudio, se investigó la hipótesis que los cambios microclimáticos no afectan de igual forma a todas las especies de una comunidad de hormigas, sobreviviendo sólo aquellas que posean adaptaciones fisiológicas que les permitan soportar el estrés generado por las nuevas condiciones de hábitat abiertos en los cafetales de sol (a libre exposición).
Para las hormigas de hojarasca puede existir un intercambio de costo-beneficio entre ser altamente competitivo y ser resistente al estrés ambiental, es decir, en condiciones similares al bosque se verían favorecidas especies altamente competitivas, mientras que en agroecosistemas intensificados se favorecerían aquellas preadaptadas al estrés ambiental. Tilman et al. (1994) determinaron un modelo que muestra como la destrucción (incluso moderada) de hábitats naturales relictuales causa extinciones determinísticas (es decir no estocásticas, sino predecibles) de los competidores dominantes de los parches remanentes. Este modelo predice que las especies se extinguen en orden del mejor al peor competidor a medida que la destrucción del hábitat se incrementa. En el contexto de los cafetales, el empobrecimiento de la diversidad y estructura de las plantas de sombra durante la intensificación conlleva a una disminución de la abundancia de las especies de hormigas de sombra mientras que las especies resistentes a condiciones abiertas aumentan en abundancia hasta dominar en cultivos a plena exposición (Risch y Carroll 1982; Nestel y Dickschen 1990; Roth et al. 1994). Un posible mecanismo para que este intercambio de especies se lleve a cabo es que las especies de áreas abiertas, que son pobres competidores en condiciones de sombra, se vean favorecidas con la intensificación debida a que están bien adaptadas al estrés fisiológico de esos hábitats abiertos. Si este mecanismo está en acción, es posible pensar que la exclusión de la especie dominante en condiciones de sombra permitirá el establecimiento de la especie dominante de condiciones de sol pero no viceversa, pues la limitación para las especies de sombra sería más de tipo fisiológico (Perfecto y Vandermeer 1996).
Se predijo, por tanto, que dos especies de hormigas, una proveniente de cafetales de sol y una proveniente de cafetales de sombra responderían diferencialmente a condiciones controladas de estrés por calor (amplias fluctuaciones de temperatura). Para examinar la hipótesis descrita anteriormente, se realizaron experimentos bajo condiciones de laboratorio, con la temperatura como indicador de fluctuación microclimática, para comparar su efecto sobre el comportamiento y mortalidad de dos especies de hormigas. Estas son: la primera Tetramorium simillimum (F. Smith), abundante en cafetales a libre exposición de la región de Apía, y la segunda Paratrechina steinheili (Forel, 1893), que se encuentra tanto en cafetales de sol como de sombra, con preferencia a estos últimos (Armbrecht et al. 2005).
Materiales y métodos
Sitios de recolección de colonias
Con base en las experiencias dejadas por el proyecto de estudios de la biodiversidad y su relación con el manejo de los cafetales (Armbrecht 2003; Rivera y Armbrecht 2005), las colonias de hormigas fueron recolectadas en el municipio de Apía, Departamento de Risaralda (5°08′N; 75°56′W), en las fincas La María y El Convenio. El área de trabajo se encuentra entre los 1400 y 1700 msnm. Según Holdridge (Espinal 1967) el sitio se define como bosque húmedo premontano (bh-Pm), con una topografía montañosa, una temperatura promedio anual de 18-20 °C, humedad relativa del 80% y precipitación promedio anual de 2320 mm.
Treinta nidos de P. steinheili fueron recolectados en la finca El Convenio, en donde esta especie, es abundante especialmente en cafetales de sombra monogenéricos (con Inga edulis Mart.). Otros 30 nidos de T. simillimum fueron recolectados en la finca La María, en este sitio esta especie es la más abundante en cafetales de sol o libre exposición. Las colonias se trasladaron al laboratorio de entomología de la Universidad del Valle, en donde se adecuaron, en nidos artificiales según método adaptado de Passera et al. (1988) y se mantuvieron en un cuarto de cría con temperaturas entre los 23 y 25°C y una humedad relativa entre 63 y 67%.
Ambas especies de hormigas fueron sometidas a tratamientos que buscaron simular las fluctuaciones de temperatura encontradas en el monocultivo a libre exposición, mientras que se mantenían colonias testigo (control) a la temperatura del cuarto de cría.
Fluctuación horaria de la temperatura de acuerdo al tipo de experimento.
Fluctuación térmica
Para establecer las fluctuaciones de temperatura presentes en los cafetales bajo libre exposición y con sombrío, en la finca La María (cafetal de sol o sin árboles de sombra) y en la finca El Convenio (cafetal con sombrío de Inga spp.), se ubicaron termómetros de registros máximos y mínimos. Este registro se realizó entre el 11 y 18 de marzo de 2003. Además, en la finca La María, con un termómetro adicional de lectura instantánea (digital), se tomaron registros de temperatura y humedad relativa a distintas horas del día durante cinco días, esto para conocer la fluctuación horaria de estas variables y establecer la simulación bajo condiciones de laboratorio. Ya que sobre estos datos se realizaron los experimentos de laboratorio, es necesario destacar que en La María se presentó una mayor amplitud térmica que en El Convenio (con temperaturas máximas mayores en promedio y menor promedio para las mínimas). Como consecuencia, el gradiente de fluctuación en la temperatura es mayor en el cafetal de sol que en el cafetal de sombra. Las variaciones de temperatura fueron de 17-31°C en el cafetal de sol y 18-28°C en el cafetal de sombra. Además, la humedad relativa fue mayor en El Convenio (75.6%) que en La María (65.0%).
Para la simulación de las fluctuaciones de temperatura de un monocultivo bajo libre exposición, se trabajó con una cámara ambiental (Industrias IFV, Minitron I, Florez-Vélez CIA. Ltda), que permitió controlar temperaturas entre los 5 y 40°C y que se programó para brindar 12 horas luz entre las 7 AM y 7 PM. La humedad relativa se mantuvo entre 62 y 69%.
Experimentos
Se ubicaron simultáneamente seis colonias de cada especie en la cámara ambiental, mientras que otras seis colonias de cada especie eran mantenidas como testigo (control) bajo las condiciones del cuarto de cría. Para el montaje de los diferentes experimentos, el número de obreras de cada colonia fluctuó entre 105 y 135 para el caso de Paratrechina y entre 230 y 276 para Tetramorium.
Para la observación del comportamiento de las dos especies de hormigas se realizaron dos tipos de ensayos, uno de fluctuación térmica gradual, comenzando desde 17°C hasta 32°C (17, 20, 24 y 32°C), éste se realizó dos veces; y el otro ensayo consistió en un choque térmico manejando temperaturas de 17 y 35°C (Tabla 1).
El primer experimento fue llevado a cabo entre el 16 y 31 de julio de 2003, el segundo entre el 8 y el 15 de agosto de 2003 y el tercero, entre el 22 de agosto y el 3 de septiembre de 2003.
Evaluación del comportamiento
Las variables evaluadas en el comportamiento de las hormigas fueron: 1. Forrajeo: número de individuos participantes de la búsqueda de alimento; 2. Actividad fuera del nido: número de individuos en la superficie del nido y 3. Proporción de mortalidad. Se realizaron dos observaciones en cinco momentos del día (7AM, 9AM, 11AM, 2PM, 4PM), para contar con registros a diferentes temperaturas durante su fluctuación diaria. En el caso del tercer experimento las variables se evaluaron sólo a dos horas del día, 7AM y 1PM (Tabla 1).
Análisis de datos
Para dos variables de respuesta: actividad fuera del nido y forrajeo, se estimó el promedio de individuos observados por día. Mientras tanto, la proporción de individuos muertos se estimó relacionando la cantidad de individuos muertos registrados durante el experimento con respecto al número inicial de individuos de la colonia. Para comparar los resultados obtenidos entre las colonias utilizadas como testigo y aquellas sometidas al tratamiento térmico, se realizaron pruebas de t mediante las herramientas del programa Excel (XP) versión 2001.
Resultados y discusión
Fluctuación térmica gradual
En el primer experimento (Tabla 2), la actividad de las obreras de T. simillimum fuera del nido no presentó diferencias entre lo observado para las colonias testigo y las sometidas al tratamiento. Sin embargo, en estas últimas hubo un menor número de obreras forrajeando. Por otro lado, hubo una mayor cantidad de individuos muertos en las colonias sometidas al tratamiento con respecto a las colonias testigo. Para P. steinheili no existió diferencia para ninguna de las tres variables consideradas (Tabla 2) y en lo referente a la mortalidad observada entre ambas especies, ésta fue mucho mayor para T. simillimum que para P. steinheili (t = 6.15; gl= 6; p<0.01) (Fig. 1).
En el segundo experimento (Tabla 2), en cuanto a T. simillimum no se corroboraron las diferencias encontradas en el primer experimento, ya que no hubo diferencias en ninguna de las variables evaluadas. Mientras tanto, P. steinheili presentó una mayor actividad de obreras en las colonias sometidas a la cámara (Tabla 2), pero no presentó diferencias en cuanto a forrajeo y mortalidad, lo que no concuerda con lo encontrado en el primer experimento. Como hecho destacable, una vez más se presentó una mayor proporción en la mortalidad de las colonias de T. simillimum sometidas a la cámara ambiental con respecto a las colonias de P. steinheili (t =4.41; gl = 6; p<0.01) (Fig. 2).
Estos dos experimentos de fluctuación térmica gradual, no fueron consistentes respecto al efecto que tiene la temperatura en los comportamientos actividad fuera del nido y forrajeo de las hormigas. Sin embargo, las colonias sometidas a la cámara evidencian una menor mortalidad relativa de P. steinheili que la especie propia de cafetal de sol, T. simillimum.
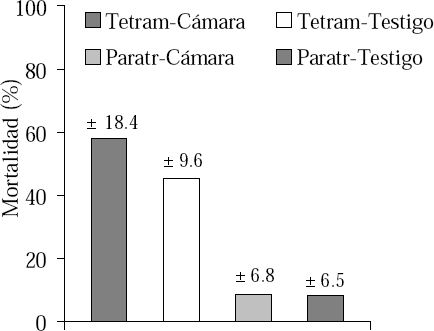
Primer experimento de fluctuación térmica gradual, comportamiento de las hormigas en mortalidad (promedio ± desviación estándar).
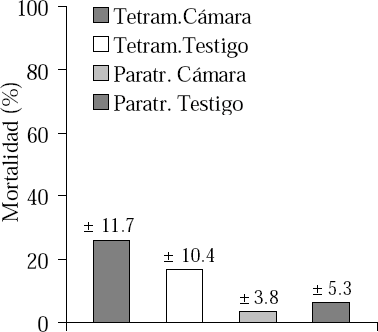
Segundo experimento de fluctuación térmica gradual, comportamiento de las hormigas en mortalidad (promedio ± desviación estándar)
Choque térmico
La actividad fuera del nido en colonias de T. simillimum no presentó diferencia estadística, aunque sí un menor forrajeo en las colonias sometidas al tratamiento con respecto a las colonias testigo (Tabla 3). Además, existió una mayor mortalidad en las colonias sometidas al choque térmico respecto a las testigo. P. steinheili tampoco mostró diferencia en la actividad y forrajeo fuera del nido (Tabla 3), aunque si se observó una alta mortalidad en las colonias sometidas a choque térmico con respecto a las colonias testigo (Tabla 3). Cabe señalar que la proporción de obreras muertas encontradas en P. steinheili no presenta diferencias con la proporción de individuos muertos observada en T. simillimum. (t= -0.46; gl=10; p>0.05) (Fig. 3).
Nuestros resultados no sugieren la existencia de una adaptación fisiológica, al menos detectable, de T. simillimum al estrés térmico en comparación con P. steinheili. Se plantea la posibilidad de que otros factores sean los que determinan la preferencia de P. steinheili a los cafetales de sombra frente a los cafetales de sol. Al respecto, cabe señalar que la temperatura (como medida de la cantidad de calor) es muy importante en la regulación de las poblaciones de hormigas, particularmente de zonas frías (Holldobler y Wilson 1990). Teniendo en cuenta que estudios anteriores indican una alta influencia de la temperatura en la distribución de las especies de hormigas (Torres 1984; Bustos y Chacón 1996; Perfecto y Vandermeer 1996), quizá la mejor forma de entender el efecto de la temperatura, en el caso de las hormigas, sea desde una perspectiva más amplia, tal y como lo sugiere Andersen (2000), como el producto combinado del clima y de la estructura del hábitat, que determina el grado de radiación solar (insolación) de la superficie de forrajeo.
Dentro del conjunto de factores climáticos, la humedad relativa ha sido sugerida como un factor de gran importancia que determina la distribución de las hormigas de suelo y hojarasca (Kaspari y Weiser 2000). De igual forma la cobertura y diversidad de árboles mostró una alta relación con la riqueza del gremio de hormigas crípticas de hojarasca en agroecosistemas cafeteros de Risaralda, de donde se extrajeron las hormigas para el presente estudio, y este hecho estuvo altamente ligado a la temperatura que presenta un sitio (Rivera y Armbrecht 2005). En el presente estudio sólo se analizó la fluctuación térmica manteniendo una humedad relativa entre 62 y 69%, que es semejante al promedio obtenido en el cafetal de sol La María (65%).
Es evidente que las fluctuaciones de temperatura pueden ser mayores en un cafetal sin sombrío respecto a uno con árboles acompañantes (en nuestro caso se encontraron casi 4 grados centígrados, en promedio, de diferencia entre las fluctuaciones máximas y mínimas de la temperatura entre el cafetal de sol, La María, y el cafetal con sombrío, El Convenio), pero este factor no explicaría, por sí solo, la reducción en la diversidad de especies. Ramírez et al. (2004) en cultivos de caña de azúcar, desprovistos de todo tipo de sombrío, encontraron que la biodiversidad de hormigas estuvo asociada directamente con el manejo que recibe cada cañaduzal y que este manejo está relacionado básicamente con la acumulación de hojarasca (previa eliminación de la quema) y, aunque estos autores no lo sugieren, podría deberse también a la disminución de las prácticas de preparación del suelo (Robertson et al. 1994), Acorde con esto, ha sido demostrado que las prácticas de labranza afectan notablemente la diversidad de especies de hormigas y una labranza de conservación o una práctica de cero labranza, como es el caso de los cafetales bajo sombrío, permitirían una mayor disponibilidad y estabilidad de los sitios de anidamiento y por ende una mayor diversidad (Peck et al. 1998).
Por otro lado, aunque en bosques primarios neotropicales no parece haber una limitación marcada por recursos de nidificación para hormigas de suelo (Kaspari 1996), en experimentos realizados en la misma zona cafetera de Apía (Risaralda, Colombia) se encontró evidencia de limitación de recursos de nidificación para Paratrechina steinheili (Armbrecht y Perfecto 2002). Esto podría explicar la disminución de esta especie en cafetales de sol, en donde la capa de hojarasca y ramitas secas es menor que en cafetales de sombra. En un estudio simultáneo de campo en la misma zona, Rivera et al. (2004) hicieron experimentos de exclusión y adición de colonias de ambas especies. Se adicionaron P. steinheili provenientes de cafetal de sombra a cafetal de sol y T. simillimum de cafetal de sombra a cafetal de sol. El estudio encontró que los trasplantes de colonias de P. steinheili eran infructuosos cuando se llevaban a cabo en cafetal de sol y exitosos si se realizaban en cafetales de sombra (control), mientras que T. simillimum sobrevivía exitosamente en ambos tipos de cafetales. Los resultados del presente estudio apoyan la idea de que otros factores ecológicos, diferentes del estrés fisiológico, influencian negativamente la sobrevivencia de P. steinheili en el cafetal de sol. Esta especie anida en hojarasca y ramitas en el suelo, mientras que Tetramorium simillimum anida en el suelo y por tanto no estaría limitada por este recurso en cafetales sin sombrío.
Esta investigación fue realizada con dos especies de hormigas que eran fáciles de manipular en el laboratorio y se asumió que el comportamiento que éstas presentaron no se puede generalizar al resto de especies integrantes de estas comunidades. Este estudio no encontró evidencia de que la desaparición de alguna de estas especies de hormigas en la transformación de cafetales bajo sombrío a cafetales con libre exposición se origine ante una imposibilidad fisiológica para tolerar las mayores fluctuaciones de temperatura resultantes. Se propone en este caso, que algunas especies de hormigas no logran adaptarse a una menor complejidad del hábitat, en donde el área y el número de sustratos aprovechables por las hormigas (troncos en descomposición, hojarasca, entre otros) para anidamiento o forrajeo es limitada (Bustos y Chacón 1996). Sin embargo, cabe considerar que otros factores pueden estar actuando en el establecimiento de las poblaciones de estas hormigas como competencia interespecífica, mortalidad por causada por enemigos naturales y las características propias del ensamblaje de las comunidades de hormigas tanto en cafetales de sol como de sombra. (Vargas et al. 2004; Rivera y Armbrecht 2005; Philpott 2005).
Experimentos de fluctuación térmica gradual y su efecto en la actividad de las hormigas fuera del nido, forrajeo y mortalidad (promedio desviación estándar)
T0.05(2), 10 = 2.28
T0.01(2), 10 = 3.16
Experimento de choque térmico y su efecto en la actividad de las hormigas fuera del nido, forrajeo y mortalidad (promedio ± desviación estándar).
T0.05(2), 10 = 2.28
T0.01(2), 10 = 3.16

Experimento de choque térmico, comportamiento de las hormigas en mortalidad (promedio desviación estándar).
Conclusiones
No se encontró evidencia que el estrés por fluctuaciones térmicas sea el factor que provoca la disminución en las poblaciones de una hormiga propia de cafetal de sombra en un agroecosistema que ha cambiado a cafetal de sol. En las dos especies de hormigas estudiadas no se detectaron efectos de fluctuación de la temperatura y choque térmico en las variables actividad fuera del nido y forrajeo.
En la fluctuación gradual de temperatura Tetramorium simillimum presentó una mayor susceptibilidad que Paratrechina steinheili en cuanto a la proporción de obreras muertas, mientras que en condiciones de choque térmico no hubo diferencias.
El efecto de la fluctuación térmica no explica por sí mismo el comportamiento de estas dos hormigas y la distribución en su espacio natural (agroecosistemas cafeteros) debe analizarse considerando otros factores como la estructura del hábitat, en términos de la mayor o menor complejidad del mismo y su oferta de sustratos de anidamiento disponible y otras variables ecológicas, que este estudio no abordó, como la competición interespecífica, la regulación por enemigos naturales y el ensamblaje de las comunidades de hormigas.
Footnotes
Agradecimientos
Los autores manifiestan sus agradecimientos a Patricia Chacón, Miguel A. Márquez, María C. Gallego, James Montoya, Patricia Rodas, Carmen E. Posso y a Loló. J. Longino identificó las especies de hormigas. Este proyecto fue financiado por Universidad del ValleCOLCIENCIAS, programa del Medio Ambiente y del Hábitat (Proyecto código1106-12-11693 a I.A.).