
Abstract
Adults of Periplaneta americana L. were evaluated under laboratory conditions to verify indirect poisoning (by cannibalism) through action of a toxic bait base on imidacloprid (2,15%). The mortality caused by direct and indirect ingestion (through cannibalism) of the bait varied between 97-100%. The average lethal time of specimens that consumed the bait directly was less (11,012 h) and significantly different than those that consumed it indirectly (65,88-96,70 h). Similarly, the average lethal knock down time of specimens that directly ingested the bait was less (0,07 h) and significantly different from average knock down times of those that acquired it indirectly, 7,98 h for those that ingested the toxin of an individual poisoned 4 h before the tests and 10,23 h from those that ingested the toxin of an individual poisoned 24 h before the tests. The results obtained support the hypothesis that the imidacloprid molecule is able to affect specimens in an indirect way (through cannibalism).
Introducción
Los Blattodea domésticos son considerados como unas de las plagas más antiguas e importantes asociadas a ambientes humanos alrededor del mundo, su éxito reproductivo se basa en aspectos biológicos y etológicos que les confieren características adaptativas especiales (Robinson 1996). Aunque sólo el 1% de las 4.000 especies de cucarachas existentes alrededor del mundo están asociadas con los humanos, son consideradas como una de las peores plagas debido a su olor repulsivo, sus hábitos poco higiénicos, la degradación de los ambientes donde se encuentran y la transmisión de enfermedades (Peterson y Shurdut 1999). Son capaces de transmitir muchos patógenos, incluyendo bacterias, virus, hongos, protozoos y helmintos que afectan la salud humana (Gordon 1996); actúan como transmisores potenciales de agentes de la diarrea bacterial (Bennett et al. 1996) e infecciones nosocomiales en hospitales (Tawatsin et al. 2001). Además, las excretas y otros desechos pueden contener gran número de alergenos que pueden ocasionar alergias respiratorias severas y urticarias, al igual que los adultos de la cucaracha americana y ninfas de primeros estadios (Bennet et al. 1996; Córdoba et al. 1999).
El hombre ha tratado durante siglos de controlar esta plaga utilizando diversos métodos, principalmente una gran diversidad de químicos sintéticos. Para que un insecticida sea aprovechado al máximo sin aplicaciones permanentes, es necesario que llegue a la mayor cantidad de especímenes en el menor tiempo posible, ya sea de forma directa o indirecta. Si se tiene en cuenta que los Blattodea son insectos potencialmente caníbales (lo cual les confiere ciertas ventajas adaptativas sobre todo en condiciones de escasez de alimento y altas poblaciones) y de alguna manera gregarias (Gordon 1996), se podría pensar que un individuo que muere cerca de sus congéneres después de haber ingerido un cebo tóxico, puede mantener la molécula del ingrediente activo y afectar a otros individuos conespecíficos que potencialmente se alimenten de ella. Esto se conoce como el efecto dominó™.
Por lo anterior, se planteó la hipótesis que probaría sí el comportamiento de canibalismo de Periplaneta americana L. ayuda en la transmisión indirecta de un insecticida, en este caso imidacloprid (ingrediente activo de Blattanex gel®).
Materiales y Métodos
Para forzar y garantizar la ingestión del cebo, los especímenes seleccionados para las pruebas (incluyendo los testigos) se dejaron en ayuno por aproximadamente 48 h antes de los ensayos.
El cebo utilizado fue la presentación comercial Blattanex Gel® (Bayer S. A.), cuyo ingrediente activo es imidacloprid (1-[(6-cloro-3-piridinil)metil]-=4,5-dihidro-N-nito-1H-imidazol-2-=amine) al 2,15%.
Las observaciones se realizaron cada 15 minutos durante la primera hora de exposición al producto, debido a que la acción del imidacloprid es muy rápida (LT50=1,7 h para hembras de B. germanica - Appel y Tanley 2000), luego cada 30 minutos durante la hora siguiente y después cada 60 minutos hasta completar ocho horas de observación. Si después de ésto quedaba algún individuo vivo, se continuaban las observaciones cada 24 h. En cada examen se anotaba si el individuo estaba en “knock down” o muerto (cuando el individuo ya no presentaba movimientos en respuesta a estímulos externos como tocarle las patas o antenas). Después de la muerte de cada espécimen, éste era disecado con la ayuda de un estereoscopio para contemplar el recorrido del producto en el tracto digestivo.
Al grupo control se le ofreció únicamente pequeños trozos de pan blando humedecidos con el colorante rojo congo.
Para esta fase se realizaron dos tratamientos: Ay B, cada uno de ellos con un total de cinco repeticiones. Para cada tratamiento se evaluaron 30 especímenes: 15 hembras y 15 machos.
Para la preparación de los especímenes que serían ofrecidos como alimento se los inducía a ingerir dos gotas del gel cuatro h (tratamiento A) y 24 h (tratamiento B) antes de los ensayos. Esto se hizo sujetando cada espécimen en una mano y con la otra se le acercaba al aparato bucal un mondadientes impregnado con una gota estándar de gel (0,2 ml). Antes de ser ofrecido a los especímenes de la unidad experimental, éstos eran limpiados con un algodón humedecido para no dejar rastros del producto sobre su cuerpo que interfiriera con los resultados de los ensayos. Luego, cada espécimen era colocado en un recipiente con las mismas especificaciones que para los ensayos de ingestión directa, con una caja de Petri que contenía un algodón humedecido para proveerles de agua; por último se introducían los seis especímenes de cada repetición.
Las observaciones se realizaron de la misma forma que para la ingestión directa. Cada 24 h se retiraban los restos del individuo consumido y se cambiaba por uno muerto en iguales circunstancias. El espécimen que moría era retirado de la caja para evitar que los demás se alimentaran de él. Al grupo control se le ofrecía una cucaracha muerta por enfriamiento.
Resultados y Discusión
De las 31 disecciones realizadas a los especímenes que murieron por ingestión directa del cebo, los lugares más frecuentes donde se encontraron rastros del producto fue en el intestino medio (80,64%) y en los ciegos gástricos (71%). Debido posiblemente al relativo corto tiempo de muerte se encontró que el 39% de los especímenes todavía presentaban una cantidad considerable del cebo en el buche. Estos resultados corroboran el hecho de que los ciegos gástricos e intestino medio son puntos importantes en la absorción (Bell 1981; Chapman 1998), lo cual sugiere que puede ser igualmente un área de absorción de la molécula de imidacloprid para su paso hacia el sistema nervioso.
Observaciones sobre el comportamiento de P. americana han mostrado que el canibalismo es posible (Gahlhof et al. 1999). Una vez colocado el individuo muerto en la caja, inmediatamente sus conespecíficos se acercaban y comenzaban a mordisquearlo en la zona de la cervix hasta arrancarle la cabeza, o mordían sus coxas hasta arrancarle las patas (la zona del cuello y las coxas son las más blandas del cuerpo del insecto). Luego comenzaban a ingerir los órganos internos hasta sólo dejar el exoesqueleto, esto también se ha observado en colonias donde el número de cadáveres encontrados es relativamente muy bajo (observaciones personales).
Como consecuencia de la ingestión directa o indirecta (vía canibalismo) del imidacloprid se presentó una alta mortalidad. La mortalidad en el grupo de cucarachas que ingirieron el cebo directamente y de aquellos que lo ingirieron vía canibalismo - 24 h (tratamiento B) fue del 100%. El porcentaje de mortalidad en el grupo que ingirió el cebo vía canibalismo - 4 h (tratamiento A) fue del 96,7%. En los grupos evaluados como controles para cada tratamiento la mortalidad presentada fue del 0%. Durier y Rivault (2000) también encontraron altos porcentajes de mortalidad en ensayos similares (51,0% para hydramethylnon y el 99,0% para fipronil).
El tiempo promedio de muerte más bajo (11,01 h) fue presentado en especímenes que consumieron el cebo de manera directa, seguido de aquellos especímenes que ingirieron el cebo de forma indirecta vía canibalismo (65,88 h para los que ingirieron un individuo preparado 24 h antes de los ensayos y 96,70 h para aquellos que ingirieron un individuo preparado cuatro h antes de los ensayos) (Fig. 1).
Entre los tiempos promedio de muerte de los especímenes que ingirieron el cebo de manera directa y de aquellos que ingirieron el cebo de manera indirecta (vía canibalismo, tratamiento A y tratamiento B), se presentaron diferencias estadísticas significativas (P<0,005).
Sin embargo, el tiempo transcurrido desde la ingestión del cebo hasta su contacto vía canibalismo con otros especímenes no parece importante; la molécula del insecticida sigue activa y es capaz de afectar a otros especímenes debido a que también causa mortalidad. La prueba de comparación múltiple de Tukey (α = 0,05%) mostró que el tiempo promedio de muerte (en horas) de los especímenes que ingirieron directamente el cebo tóxico fue significativamente diferente a los tiempos promedio de muerte de los especímenes que ingirieron el cebo indirectamente (vía canibalismo), tanto los del tratamiento A como del tratamiento B, pero no se encontraron diferencias estadísticas significativas entre los tratamientos por ingestión indirecta del mismo (Ay B).
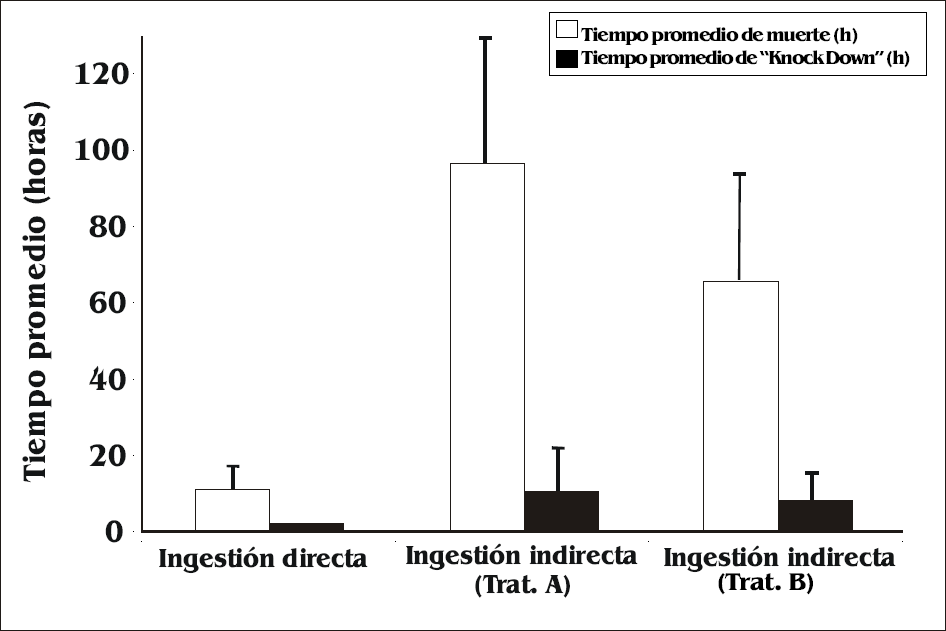
Tiempos promedios de muerte y “knock down” (± SEM) de P. americana que ingirieron imidacloprid de manera directa e indirecta (vía canibalismo) (Tratamientos A y B).
Sin embargo el tiempo promedio de muerte de los especímenes que ingirieron el cebo de forma indirecta a partir de especímenes de 24 h de muertos, fue menor que el tiempo del tratamiento con especímenes de cuatro h de muertos. Esto puede deberse a que como el tóxico entra vía intestino, y tiene que pasar por los puntos de absorción del mismo para llegar al sitio de acción en el sistema nervioso, entre más tiempo pase antes que el individuo sea consumido mayor cantidad de tóxico logrará pasar al hemocele y cubrir una mayor área del interior del cuerpo del insecto; por lo tanto habrá mayor cantidad de ingrediente activo atacando el órgano blanco. Con estos resultados se apoya la teoría de muerte secundaria por ingestión indirecta de un cebo tóxico (vía canibalismo) y difiere de Durier y Rivault (2000) quienes sugieren que la muerte secundaria real se produce por trazas del tóxico que quedan impregnadas en el cuerpo del espécimen o en el ambiente, dado que en los ensayos realizados se eliminó esa fuente de variación al limpiar muy bien los especímenes antes de introducirlos a las cajas de prueba para que fueran consumidos. Además, si la ingestión fuera directa, entonces el tiempo promedio de muerte observado en la ingestión indirecta del cebo no debería ser relativamente diferente al tiempo promedio de muerte observado por ingestión directa del mismo.
De igual manera, el tiempo promedio de “knock down” de aquellos especímenes que ingirieron el cebo tóxico de manera directa (0,07 h) fue significativamente menor que el de aquellos que adquirieron el cebo de forma indirecta (vía canibalismo) (Fig. 1), 10,23 h para aquellos que adquirieron el cebo de un individuo envenenado cuatro horas antes y 7,98 h para aquellos que lo adquirieron de un individuo envenenado 24 h antes de los ensayos. Sin embargo, al realizar una prueba de T-student, no se encontraron diferencias estadísticas entre ellas (T0.05 = -0,13/P = 0,79). También se observó una tendencia similar a que el tiempo promedio de “knock down” de aquellos especímenes que adquirieron el tóxico por un individuo envenenado 24 h antes de los ensayos fuera menor al de aquel de los especímenes que adquirieron el tóxico de un individuo envenenado cuatro h antes de los ensayos; la explicación a esta tendencia es similar a la expuesta anteriormente para los tiempos promedio de muerte.
El hecho de que el tiempo promedio de “knock down” por ingestión directa del cebo fuera tan bajo, aunque es un indicio del buen desempeño del producto, puede llegar a ser un problema para el control de esta plaga; pues si un individuo cae muy rápido en “knock down” y no alcanza a llegar hasta el nido o a un lugar resguardado donde no sea fácilmente descubierta por el “ama de casa”, no podrá ser encontrado fácilmente por sus conespecíficos para ser consumido.
Los tiempos promedio de “knock down” para machos y hembras por ingestión directa del producto no fueron estadísticamente diferentes (T= -0,21/df= 19/P= 0,8364); de igual forma ocurrió para ambos sexos por ingestión indirecta del imidacloprid en el tratamiento A (T= 0,94/df= 14/P= 0,3609) y para machos y hembras por ingestión indirecta en el tratamiento B (T= 1,62/df= 14/P= 0,12). Tampoco se encontraron diferencias estadísticas significativas entre los tiempos promedio de muerte entre machos y hembras que ingirieron el cebo de forma directa (T= 2,03/df= 19/P= 0,0569); y entre machos y hembras que ingirieron imidacloprid de forma indirecta (vía canibalismo) en el tratamiento A (T= 1,17/df= 13/P= 0,2617). Por el contrario, el tiempo promedio de muerte de machos fue significativamente menor (21,90 h) que el de las hembras (81,53 h), cuando se presentó ingestión indirecta del producto en el tratamiento B (T= 3,24/df= 14/P= 0,0059). Los resultados del LT50 (en h) por ingestión directa e indirecta de imidacloprid se aprecian en la tabla 1.
LT50 (en horas) observado en especímenes de P. americana que ingirieron imidacloprid directa e indirectamente (vía canibalismo)
Los resultados del LT50 variaron desde las 2,52 h para el tratamiento B de ingestión indirecta, hasta 35,19 h para el tratamiento A por ingestión indirecta. El LT50 por ingestión indirecta del cebo en el tratamiento A fue el más alto (21,69 h), seguida por el LT50 de los especímenes que ingirieron el cebo indirectamente en el tratamiento B (8,47 h) y por último el LT50 de los especímenes que ingirieron el cebo de manera directa (7,66 h). No hubo diferencias significativas entre los valores del LT50 para los dos tratamientos por ingestión indirecta de imidacloprid, tampoco entre el LT50 por ingestión indirecta del producto en el tratamiento B y el LT50 por ingestión directa del producto. El LT50 por ingestión indirecta del producto en el tratamiento A fue significativamente diferente del LT50 por ingestión directa de imidacloprid (Tabla 1).
Gahlhoff et al. (1999) al trabajar con la cucaracha alemana (B. germanica) encontraron que por ingestión directa de varios cebos tóxicos (abamectin, ácido bórico, chlorpyrifos, fipronil e hydramethylnon), los resultados de los LT50 fueron más altos que los presentados en este trabajo (Tablas 1 y 2). Estos resultados sugieren que el imidacloprid, al ser ingerido de manera directa por las cucarachas, es de acción más rápida que otros cebos tóxicos con características similares de presentación comercial pero con otros ingredientes activos. Mientras que el LT50 para ingestión indirecta de imidacloprid fue de 84,14 h, los LT50 para muerte secundaria por otros insecticidas puede llegar hasta las 257,52 h (Chlorpyrifos) (Gahlhoff et al. 1999).
Los valores del LT50 para machos y hembras variaron desde 3,48 h para los machos del tratamiento B (ingestión indirecta) hasta 84,14 h para hembras del tratamiento A (ingestión indirecta). El LT50 para hembras del tratamiento A (ingestión indirecta) presentó diferencias significativas con el LT50 de hembras y machos para ingestión directa de imidacloprid (Tabla 2). Los demás grupos no presentaron diferencias significativas entre ellos.
LT50 (en horas) observado en hembras y machos de P. americana en ingestión directa e indirecta de imidacloprid
Appel y Tanley (2000), al trabajar con un cebo experimental a base de Imidacloprid al 2,15%, encontraron diferencias significativas entre el LT50 de hembras de B. germanica (1,72 h (0,001-7,33)) y el LT50 de machos (30,65 h (24,23-36,37)). Los resultados presentados en este trabajo además de demostrar que no hay diferencias significativas entre los LT50 para machos y hembras por ingestión directa del producto, también muestran un LT50 menor para machos. El LT50 para hembras de P. americana por ingestión directa de imidacloprid aunque es un poco mayor al LT50 para hembras de B. germanica, no presenta diferencias significativas entre ellas. Esto sugiere que el metabolismo del imidacloprid en hembras y machos es posiblemente similar en P. americana, no pareciendo ser así en B. germanica, cuyas hembras son más susceptibles que los machos y mueren más rápido.
Los resultados del presente trabajo prueban la hipótesis de que la molécula de imidacloprid es capaz de afectar a especímenes (tanto hembras como machos) que la han adquirido de forma indirecta vía canibalismo, lo cual puede ser de importancia para optimizar el uso del producto.
Conclusiones
El canibalismo es una opción de alimentación en P. americana, cuando se presentan condiciones de escasez o disponibilidad de alimento. Lo anterior es una de las formas cómo la molécula de imidacloprid, vía canibalismo, llega hasta el órgano blanco y afecta el metabolismo de hembras y machos de P. americana, ocasionando mortalidades mayores que el 90%, lo cual puede ser de importancia para optimizar el uso de este producto.
A pesar de que la muerte secundaria por ingestión indirecta de este cebo tóxico (vía canibalismo) está ocurriendo, el tiempo letal medio de especímenes que lo ingirieren directamente es menor que para aquellos que lo consumen indirectamente, vía canibalismo; es decir, que el tiempo de saturación es superior al de la dosis por ingestión indirecta, indicando con esto que la cantidad del ingrediente activo disponible para la ingestión es menor que la que se obtendría de manera directa.
Footnotes
Agradecimientos
Los autores agradecen la colaboración brindada por el biólogo Darío Correa en la realización de los ensayos, la ingeniera Fanny Rivera y Gonzalo Fernández de la empresa BAYER S. A por su apoyo. Y a la empresa BAYER S. A por la financiación del mismo.