
Abstract
The sustainability of butterflies farming for commercial purposes was determined at "Peña Roja" indigenous community (Middle Caquetá River, Colombian Amazons), based on biological, economical and social selection criteria. During August 1999 the economical interest butterflies was identified. Heraclides anchisiades anchisiades was selected for exploring its economical potential and the viability of its grow in captivity as a model. This species was selected based in the number of postures and its survival rate. Between February and July 2000 it was observed the host preferences, natural enemies, oviposition and herbivory behavior, the consumed foliar area, the survival and longevity of each stadium of development cycle, starting from eclosioned larvae of collected postures in Citrus sinensis and Citrus reticulata (Rutaceae). During February to April, the eclosioned larvae were individually kept in plastic glasses and the food supplies come from their hosts. The survival in C. reticulata was greater and there were no significant differences in the longevity of each stadium related to the host. Between May and July three larvae were maintained in only one plastic glass and the nutritive quality of Citrus species was tested; this resulted in a survival increase and the decrease of stadium longevity on C. sinensis, while with C. reticulata the survival rate was kept stable. Based on literature search about the demand of this butterfly and with the results founded in this research was carried out an economical analysis of cost-benefits (TIR). This analysis indicated that the farming of the studied specie is economical profitable. Additionally, the social characteristics of the region and the culture pattern at Peña Roja community, were analyzed in relationship with the butterflies, which pointed out that within the traditional knowledge of the community, the butterflies are not considered sacred animals, which explains the positive answer of the community to the proposal of farming these species in the community. We may conclude that the farming of this specie in the study area is biological, economical and social sustainable.
Introducción
La búsqueda de alternativas de explotación sostenible de los recursos naturales ha propiciado el desarrollo de estrategias de manejo productivo de la biodiversidad mediante trabajos de investigación y apropiación de tecnologías locales. Una de estas alternativas es la cría comercial de mariposas ornamentales, un recurso forestal no maderable promisorio que se puede implementar en Colombia debido a su alta biodiversidad (Constantino 1997), siendo el tercer país con mayor diversidad de mariposas diurnas, con unas 3019 especies descritas hasta el momento, de las cuales 300 son endémicas (Andrade-C 2000). Para iniciar un programa de cría de mariposas el conocimiento de su biología es fundamental. Sin embargo, existen muy pocos estudios en el país llevados a cabo sobre las poblaciones de mariposas con potencial económico, destacándose los trabajos de Luis Miguel Constantino en el Valle del Cauca (Fundación Herencia Verde, 1997). Adicionalmente, en el Jardín Botánico del Quindío y el Centro de Investigaciones de Café (Cenicafé), se están adelantando estudios sobre el ciclo de vida de lepidópteros diurnos con el fin de iniciar programas de conservación de los mismos en la zona cafetera Colombiana, dentro de los cuales se ha planteado la cría en cautiverio (Gil-Palacio et al. 2000). Para la Amazonía Colombiana sólo se cuenta con algunos inventarios de especies de mariposas para determinadas localidades, pero no hay estudios publicados que abarquen aspectos amplios sobre su biología e historia natural.
El ciclo de vida de una mariposa se inicia cuando las hembras seleccionan cuidadosamente una planta hospedera para poner sus huevos. Esta selección tiene una base química (Brower y Brower 1964), pero la identificación inicial del hospedero es visual, probablemente basada en el color y forma de la hoja. Una vez sobre la planta, la mariposa corrobora la identificación por percepción química (Gilbert 1975; Rausher 1980, 1983). En papiliónidos, la hembra deja sus huevos en respuesta a ingredientes químicos específicos contenidos en sus plantas hospederas, percibiendo los componentes esenciales a través de sus quimioreceptores tarsales; los estimulantes de oviposición de muchos papiliónidos consisten en múltiples componentes entre los cuales se incluyen glicósidos flavonoides, alcaloides y aromáticos (Nishida 1995). Si bien el adulto se alimenta exclusivamente de fluidos (néctar, exudados azucarados o fermentos de frutas), la mayoría de las larvas de mariposa son herbívoros especialistas (Ehrlich y Raven 1964) y para estos la supervivencia puede afectarse por la calidad del hospedero, dado que una baja calidad nutricional de la hoja reduce la sobrevivencia de la larva de manera indirecta, al incrementar el período de exposición a enemigos naturales (Loader y Damman 1991). Elementos como nitrógeno (determinante en la tasa de crecimiento y niveles de reproducción poblacional), fósforo, azufre y potasio se encuentran en mayor proporción en las zonas de crecimiento de la planta, siendo áreas generalmente preferidas (Feeny 1975; Medina 1977; Scriber y Feeny 1979). Varios papiliónidos se encuentran fuertemente asociados con Rutáceas, la mayoría pertenecientes a la tribu Papilionini, a la cual pertenecen las mariposas del género Heraclides.
Las mariposas están en constante riesgo a causa de diversos factores que influyen en su probabilidad de sobrevivencia; los huevos y orugas jóvenes están sujetos a una extensiva depredación por parte de artrópodos parasitoides (Chalcidae, Trichogrammatidae y Scelionidae, entre otros), e incluso por hongos patógenos (que pueden afectar a los adultos), mientras que otras familias pertenecientes a Hymenoptera, Diptera, Orthoptera, Heteroptera y Aranea (Ctenidae y Lycosiadae) son enemigos naturales de orugas o pupas (DeVries 1987). Por su parte los adultos son depredados por invertebrados (arañas, mántidos, hormigas, odonatos, entre otros) y vertebrados (aves, reptiles y mamíferos; De Vries 1987).
Desafortunadamente muchas especies de mariposas se encuentran en vía de extinción debido principalmente a la destrucción de sus hábitats y a su extracción masiva y selectiva, lo que hace imprescindible hacer aportes para contribuir a su conservación, mediante la investigación sobre su cría, mantenimiento y liberación (Gil-Palacio et al. 2000). La demanda internacional de mariposas es básicamente generada por cuatro sectores: coleccionistas, artesanías e industrias de adornos, museos y granjas o vivarios de mariposas. Los tres primeros sectores requieren adultos disecados y preservados, mientras que el cuarto requiere orugas vivas y pupas recién formadas. La venta de ejemplares poco comunes y/o raros es uno de los renglones más lucrativos. Los países lideres en producción de mariposas son Malasia, Filipinas, Corea, Hong Kong Tailandia, Taiwan, Kenya, Madagascar, Costa Rica, El Salvador y Papua Nueva Guinea. En este último, se ha observado que la producción de mariposas en parcelas de cultivo sirve para evitar la pérdida de diversidad genética (salvando a especies en vía de extinción), apoyando también la conservación de especies y sus hábitats naturales (Hutton 1985).
La cría de mariposas promueve un lazo fuerte entre la conservación y el desarrollo, disminuyendo la presión que sobre poblaciones naturales ejerce la captura, proveyendo así individuos de mayor calidad y en mayor cantidad a los mercados dado que las mariposas del bosque generalmente presentan alas desgastadas debido al ataque de depredadores o por el contacto con las ramas de los árboles (Constantino 1996). En la actualidad, se manejan tres sistemas de cría, los cuales son cría in situ o enriquecimiento del bosque; rancheo o asociación de cosecha sostenible en vida libre con formas de producción ex situ y la cría ex situ o cría intensiva en vivarios que generalmente están por fuera del hábitat de las especies, por lo cual no considera la conservación del hábitat (Constantino 1997). Otra alternativa para aprovechar el valor escénico de las especies tropicales, conjuntamente con la flora y el paisaje, es el turismo ecológico; siendo esta una actividad no destructiva del recurso para la cual Colombia tiene un gran potencial.
De acuerdo con Rudas (1999), para que el aprovechamiento comercial de un producto de la biodiversidad sea sostenible debe cumplir, al menos, con que la tasa de aprovechamiento del producto sea menor que su tasa de regeneración (sostenibilidad biológica); que los costos totales reales de producción sean menores que los beneficios que se obtengan (sostenibilidad económica) y que los beneficios netos del aprovechamiento sean repartidos equitativamente entre los distintos sectores de la población, generando niveles de ganancia adicional adecuados incluyendo los costos ambientales y los costos sociales de los impactos negativos presentes y futuros generados por el aprovechamiento- (sostenibilidad social).
En el presente estudio se generaron espacios para la integración de la comunidad en su desarrollo, de modo que ésta fue permanentemente consultada y participó en el proceso de toma de decisiones, disminuyendo así conflictos al interior de la comunidad. Dado lo anterior, se inicia con este trabajo un proceso de investigación participativa con los habitantes de la comunidad indígena de Peña Roja, con el fin de evaluar la sostenibilidad de la cría de mariposas como alternativa económica específicamente para esta zona, teniendo en cuenta, además, las características sociales y culturales de la comunidad para, a mediano plazo, iniciar el proceso de cría de mariposas con potencial económico dentro de la comunidad.
Materiales y Métodos
La investigación se llevó a cabo en la Comunidad Indígena de Peña Roja, cuenca media del río Caquetá, ubicada entre los 0° 39' L.S. y los 72° 06' L.W. (Fig. 1). Esta comunidad se encuentra conformada principalmente por familias de la etnia Nonuya, aunque existen otras pertenecientes a las etnias Bora, Miraña, Paky, Matapí, Uitoto, Yukuna, Cubeo y Tukano. La zona tiene un clima ecuatorial superhúmedo (26° C de temperatura media y 3000 mm de precipitación media); diciembre a marzo representa el período con menor precipitación y los más lluviosos van de abril a junio y de septiembre a noviembre. La mayor parte del área alcanza una altitud de 200-270 m, correspondiendo su zona de vida a bosque húmedo tropical (bh-T, sensu Holdridge, 1963); en la zona de estudio se encuentran la mayoría de las unidades geomorfológicas descritas para el Medio Caquetá (Martínez y Galeano 1994).
Ubicación del área de estudio. a) Región de Araracuara. b) Detalle -Comunidad indígena de Peña Roja.
En agosto de 1999 se llevó a cabo la fase preliminar, en la cual se colectaron mariposas con el fin de identificar la lepidopterofauna y seleccionar las que presentaban potencial económico. El muestreo y colecta se realizó en un período de 26 días, utilizando captura mediante jama y observación visual a lo largo de transectos de longitud no definida y captura mediante trampas con cebo (Fagua 2001). Para cada uno de los sistemas de paisajes definidos en el área de estudio se instalaron 25 trampas Van Someren-Rydon, una cada 50 m, a lo largo de 3 transectos de 250 m y otro de 300 m. Las trampas se colocaron a diferentes alturas (1m, 3 m y más de 3 m), utilizando orina, excrementos, frutas y pescado en descomposición como cebo; éstos se depositaban en las trampas todos los días entre las 7 y 8 a.m. Las trampas se revisaron cada 3 horas entre las 8 a.m. y las 4 p.m. (a esta hora se cerraban, totalizando 200 horas de esfuerzo de captura por día y un total de 5200 horas para el muestreo). El jameo se realizó principalmente a lo largo de trochas y curso de quebradas durante el tiempo entre las revisiones de las trampas. Esta actividad se realizó de 7 a.m. a 12 m y de 1 p.m. a 4 p.m. En las localidades más cercanas a la sede de trabajo de TROPENBOS (en el área de Peña Roja) se realizó un muestreo adicional entre las 5 p.m. y las 6:30 p.m. con el fin de capturar especies de hábitos crepusculares. Para toda la actividad de captura se contó con la colaboración de Fabián Moreno (guía indígena de la comunidad). Dado lo anterior, el esfuerzo de captura total fue de 416 horas (208 por cada observador) más 39 horas de observación del muestreo de la tarde. El material colectado se encuentra depositado en las colecciones de Entomología del Instituto de Ciencias Naturales de la Universidad Nacional de Colombia y de la Pontificia Universidad Javeriana.
Para la fase formal de toma de datos, llevada a cabo desde febrero hasta julio del 2000, se seleccionó a Heraclides anchisiades anchisiades (Esper, 1788) por presentar un número de posturas y tasas de supervivencia comparables numéricamente. En campo se observaron las plantas utilizadas por las hembras para ovipositar y se identificó el mayor número posible de plantas registradas como hospederos para H. anchisiades; las plantas fueron monitoreadas diariamente en las horas de la mañana, para encontrar nuevas posturas. A las hospederas utilizadas y las potenciales se les midió altura, DAP (diámetro del tronco a la altura del pecho), número estimado de hojas, características de la corteza viva y muerta, y estado fenológico, realizando sus respectivas colecciones botánicas, las cuales se depositaron en el Herbario Amazónico (COAH) del Sinchi y Herbario de la Pontificia Universidad Javeriana (HPUJ). Se observó la ubicación, características y número de huevos por postura; adicionalmente se realizó el seguimiento de las hembras para observar y registrar el comportamiento de oviposición, contabilizando el tiempo total empleado por postura. Posteriormente, se colectaron posturas frescas de las plantas hospederas identificadas y se distribuyeron los huevos en vasos plásticos desechables. Inicialmente, durante los meses de febrero abril (época seca), se colocó un individuo (larva) por vaso plástico y se les suministró material vegetal fresco proveniente de su respectivo hospedero. Posteriormente, durante los meses de mayo a julio se realizó una prueba de dieta, y se colocaron tres individuos por vaso, con el fin de observar si así se optimizaba la tasa de supervivencia y por ende, el proceso de cría.
Se realizaron descripciones detalladas para cada estadio del ciclo de desarrollo, anotando color, longitud corporal, medida y coloración de la cápsula cefálica (las orugas se colectaron en alcohol etílico al 70%). Se observó si se encontraban parásitos, parasitoides, depredadores y patógenos de los diferentes estadios del ciclo de vida de la mariposa; de los tres primeros se realizaron colecciones en alcohol (al 70%) para su posterior identificación en el Laboratorio de Entomología de la Pontificia Universidad Javeriana. A partir del total de huevos colectados, se registró la cantidad de individuos eclosionados, los que pasaron a cada ínstar, a pupa y de ahí a adulto bajo condiciones de laboratorio; estos datos se trabajaron como probabilidades (Ix) con el fin de establecer comparaciones entre las tasas de supervivencia obtenidas para los diferentes hospederos y para cada época de estudio en particular. Se analizó la mortalidad específica por cada estadio de desarrollo (qx), para observar cual fué más crítico, y por ende, de mayor cuidado para la cría, y la mortalidad respecto a la cohorte (dx), para conocer el comportamiento de la población bajo condiciones de laboratorio. Se realizaron cálculos de la mortalidad específica por estadio (qx) y respecto de la cohorte (dx), los cuales también se compararon de acuerdo con el hospedero y la época de estudio. Se midió la duración de cada estadio de desarrollo a partir de las orugas provenientes de las posturas colectadas y llevadas a laboratorio; para los ínstares la duración se tomó basándose en el período en días entre muda y muda para cada individuo; estos datos se promediaron para compararlos de acuerdo con la dieta y el período de colecta. Los cambios de un estadio a otro se determinaron por la muda, dejando la cápsula cefálica como testigo conservada en alcohol al 70%.
Se realizaron observaciones de los hábitos de herbivoría de las orugas tanto en campo como en laboratorio. En laboratorio se midió el área foliar consumida por oruga (provenientes de las posturas colectadas) para conocer la cantidad de material vegetal necesario para alcanzar el estadio adulto, información necesaria para el proceso de cría; para este último se siguió la metodología de Fagua y Ruiz (1993) y Fagua et al. (1998). En campo se realizaron anotaciones sobre los hábitos de herbivoría de las orugas y se colectó material vegetal fresco para suministrarles a las orugas en laboratorio. Para determinar las tasas de herbivoría específicas se observó el consumo promedio por individuo durante su desarrollo larval y se analizó lo que este equivale en comparación a la planta. Para estimar la tasa de herbivoría diaria se realizaron dibujos en papel milimetrado del área foliar consumida por individuo (medida utilizando el equipo "Portable area meter", marca Li-COR, modelo Lf-3000). Las orugas próximas a empupar, o prepupas, se trasladaron a jaulas de madera forradas en tela tull para el empupado (aunque la mayoría empuparon dentro del mismo vaso plástico). Estas jaulas se ubicaron en la sombra, en sitios frescos resguardados de la lluvia y protegidas de la presencia de hormigas y arañas; las pupas se colocaron en soportes de alambre y para evitar la desecación se regaron con aspersión fina de agua 2 veces por semana (Constantino 1996).
El número de huevos por postura según el hospedero se comparó mediante una Prueba U de Wilcoxon-MannWithney. Las tasas de supervivencia (Ix) y de mortalidad (qx y dx) según el hospedero y según la época de estudio se compararon mediante una Prueba de rangos asociados a Wicolxon. La duración entre estadios según el hospedero y según la época de estudio se analizó por medio de una Prueba de Signos. Finalmente, los resultados obtenidos de área foliar consumida, de acuerdo con el hospedero y según la época, fueron comparados según pruebas de Kolmogorov-Smirnov para dos muestras.
Posteriormente, con el fin de analizar la factibilidad del proyecto de cría en esta comunidad indígena, se analizaron los tres criterios de selección para evaluar la sostenibilidad del proyecto tomando a H. a. anchisiades como referencia a para los parámetros biológicos del modelo de cría. Desde el punto de vista económico se realizó un análisis costo beneficio (análisis de rentabilidad) a partir de los resultados obtenidos del experimento de cría, la producción anual estimada de mariposas y los costos de los materiales requeridos durante el curso de la investigación para la cría de las orugas. El análisis se proyectó a 5 años, agrupándose los datos obtenidos de forma anual. Finalmente, como criterios sociales, se tuvieron en cuenta los patrones culturales de la Comunidad de Peña Roja para entender las acciones sobre ella con el fin de evaluar los posibles impactos culturales que generaría el proyecto en esta comunidad; para esto se llevaron a cabo reuniones con los miembros de la comunidad, en las cuales se dio a conocer el objetivo del trabajo y las expectativas de ellos referentes al mismo, y los planes futuros del proyecto. Adicionalmente, se consideraron las actuales condiciones sociales y de orden público de la región que pueden afectar de manera directa el proceso de cría y la comercialización de las mariposas para así proponer el sistema más adecuado para la zona de estudio.
Resultados
Se colectaron 102 especímenes pertenecientes a cinco familias, 14 subfamilias (Fig. 2), 47 géneros y 61 especies, 43 de las cuales son de interés comercial (70%; Tabla 1). En cuanto a la especie seleccionada como modelo exploratorio de cría (H. a. anchisiades), todos los hospederos registrados pertenecen a la familia Rutacea (DeVries 1987; Tyler et al. 1994; Brown et al. 1995; Nishida 1995; Constantino 1998). Para la zona se colectaron posturas en Citrus reticulata y Citrus sinensis. De C. reticulata se monitorearon 6 árboles (sólo se encontraron posturas en 2, para un total de 1002 huevos) y para C. sinensis 11 árboles (4 con posturas y 1127 huevos en total); adicionalmente se monitorearon otras tres especies de Rutáceas presentes en la zona: Citrus limon (12 árboles, dos con orugas gregarias de 3er. y 5º. ínstar), Citrus limetta (2 árboles, sin huevos ni orugas) y Zanthoxylum sprucei (1 árbol, sin huevos ni orugas). Todas estas especies, salvo Z. sprucei, son introducidas y cultivadas por los indígenas de la comunidad alrededor de sus viviendas. El monitoreo a los hospederos efectivos y potenciales se realizó entre el 17 de febrero y el 12 de julio. Adicionalmente, se destaca que se observó a la hembra libando en flores de Inga sp. (Mimosaceae).
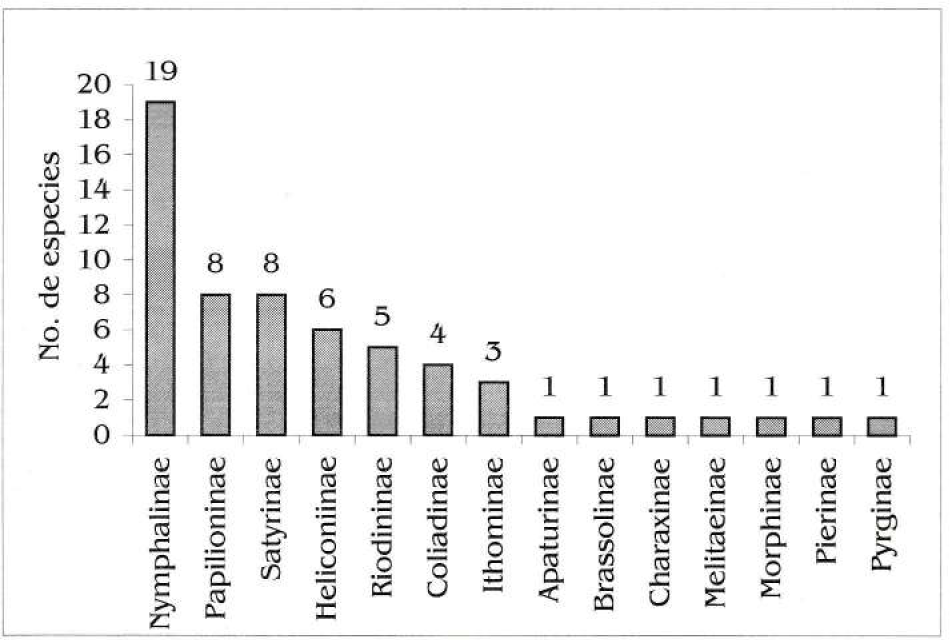
Composición por subfamilias de lepidópteros diurnos colectados entre agosto de 1999 y julio del 2000 en la Comunidad de Peña Roja.
Respecto a los parámetros biológicos de Heraclides anchisiades anchisiades, se encontró que sus huevos son esféricos, de color amarillo, cubiertos con ornamentaciones irregulares más oscuras y diámetro de 0.9 mm. El tiempo promedio de eclosión fue de 6 días (n= 431, S= 1.83, Rangos 5-7). Los huevos son colocados de manera gregaria y, a medida que se acerca la eclosión, se van tornando más oscuros. La oruga de primer ínstar es marrón, en segundo ínstar es anaranjada y en tercero es verde; a partir del cuarto se oscurece y aparecen manchas dorsales blancas; en quinto ínstar es negra aterciopelada, con las manchas blancas más grandes y notorias hacia los lados y centro del cuerpo. La cápsula cefálica en todos los ínstares es naranja, tornándose más oscura con cada ínstar (en la Tabla 2 se registran las dimensiones corporales y duración del período de desarrollo promedio de acuerdo a la dieta y época de estudio). Las orugas se alimentan en todos sus ínstares de manera gregaria y en la noche; durante el día se encontraron individuos de quinto ínstar descansando en la base del tronco de los árboles, mientras que las orugas de tercero y cuarto ínstar se observaron agrupadas en racimo en el ápice de las ramas.
En el estado de larva duraron 41 días en promedio durante febrero abril y 31 días durante mayo julio bajo dieta de Citrus sinensis y 43 días en febrero - abril y 39 días durante mayo julio con Citrus reticulata. En prepupa la duración fue siempre de 1 día. La pupa es grisácea con regiones verdosas y apariencia de liquen; su longitud promedio es de 3,8 cm (n=25, S= 0,17, Rangos 3,6-4,1) y permaneció en este estadío 15 días en promedio. El ciclo de desarrollo total alcanzó los 62 días durante febrero - abril (n=3, S= 4.93, Rangos= 57-66) y 51 días durante mayo - julio (n=7, S= 2.79, Rangos= 47-55) con dieta de C. sinensis y de 65 días (n=11, S= 3.11, Rangos= 57-67) y 59 días (n=4, S= 2.16, Rangos= 53-58) durante los mismos períodos para C. reticulata. Del total de posturas colectadas durante el estudio, (15 posturas colectadas en C. sinensis -1127 huevos en total- y 17 en C. reticulata -1002 huevos en total), la mayoría de las no eclosionadas para C. sinensis fue por parasitoides (Chalci-dioidae), en tanto que para C. reticulata la mayor causa de mortalidad fueron los hongos; además de esto se encontraron mirmicinos depredando huevos (Fig. 3a). En campo no se observaron enemigos naturales de orugas, sin embargo, en condiciones de laboratorio se tuvo una mortalidad alta de las mismas causada por bacterias.
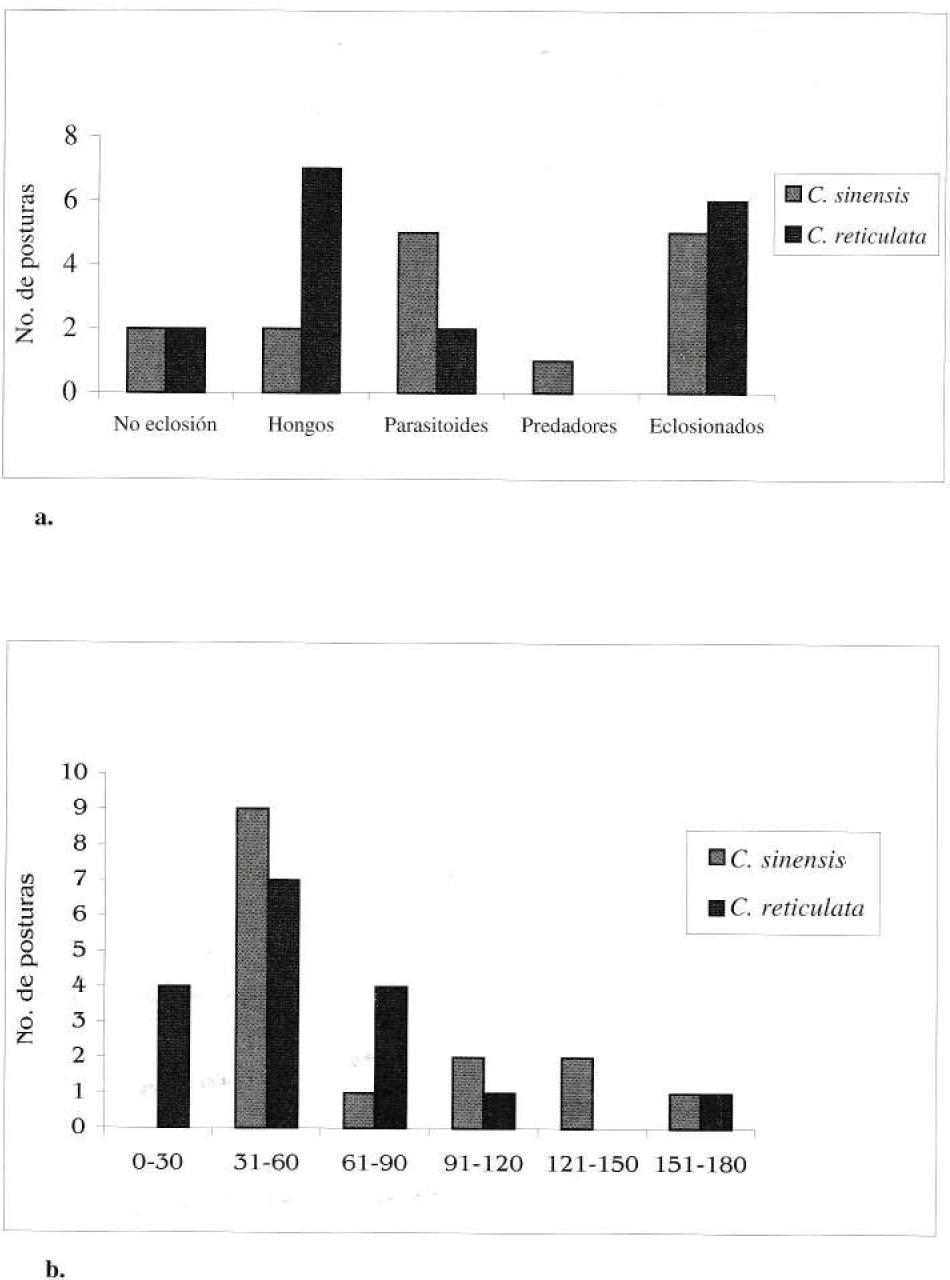
Características de las posturas de Heraclides anchisiades anchisiades. a) Número de posturas eclosionadas, no eclosionadas, parasitadas o depredadas de acuerdo con el hospedero. b) Número de huevos por postura de acuerdo con el hospedero.
Lista de las especies de lepidópteros diurnos colectados en la Comunidad de Peña Roja entre agosto de 1999 y julio del 2000
Especie de interés comercial y sus hospederos registrados en literatura.
Dimensiones promedio de la cápsula cefálica, longitud de la oruga y duración del período de desarrollo de Heraclides anchisiades anchisiades. (F-A= Febrero-Abril, M-J= Mayo-Julio). a) Con dieta de Citrus sinensis. b) Con dieta de Citrus reticulata
Para la oviposición la hembra escogía dos a tres árboles cercanos de las dos especies hospedero y dentro de un mismo árbol llegaba siempre a las mismas hojas (hasta 3 hojas por cada árbol); sobre éstas volaba alternadamente y se posaba pocos segundos explorando la superficie con sus antenas, mientras que con su abdomen tocaba el envés, se decidía por una hoja y comenzaba a ovipositar. Todas las posturas fueron colocadas en el envés de hojas basales jóvenes (.C reticulata) y en hojas medias jóvenes (C. sinensis) de la parte apical de las ramas, siempre a la derecha de la nervadura principal, en la zona de sombra permanente. El rango más frecuente de número de huevos por postura estuvo entre 51 y 60 huevos; C. sinensis, presentó un número de huevos por postura más alto en comparación con C. reticulata (Fig. 3b), aunque no hubo diferencias significativas (U de Wilcoxon-MannWithney, U' = 105, p>0.05).
En febrero - abril (época seca, cuando se colocó un individuo por vaso), se observó una mayor tasa de supervivencia (Ix) por estadio en C. reticulata, llegando a adulto un 6% de la población inicial; en tanto que para C. sinensis, el Ix fue menor y sólo un 2% llegó a adulto (Fig. 4a); sin embargo estas diferencias no fueron significativas (rangos asociados a Wicolxon, T=0.031, N=6, p>0,05). En mayo a julio (época húmeda, con tres individuos por vaso) se realizó una prueba de dieta a partir de individuos de una postura colectada en C. sinensis, suministrando a una parte del grupo C. sinensis y a la otra C. reticulata (para observar si la tasa baja de supervivencia era debida a un error en la selección del hospedero por parte de la hembra, asumiendo que C. reticulata es el hospedero óptimo en la zona de estudio); se decidió también colocar tres individuos por vaso con el fin de optimizar el consumo del material vegetal. Durante esta prueba, el número de adultos obtenidos en C. sinensis aumentó a un 19%, aunque para C. reticulata se mantuvo estable (6%), siendo el Ix por estadío significativamente diferente en C. sinensis (rangos asociados a Wicolxon, T=0.0312, N=6, p<0,05, Fig. 4b). También en C. sinensis se observó una diferencia significativa entre los lx por época (rangos asociados a Wicolxon, T=0.0312, N=6, p<0,05), mientras que en C. reticulata no las hubo (rangos asociados a Wicolxon, T=2, N=6, p>0,05).
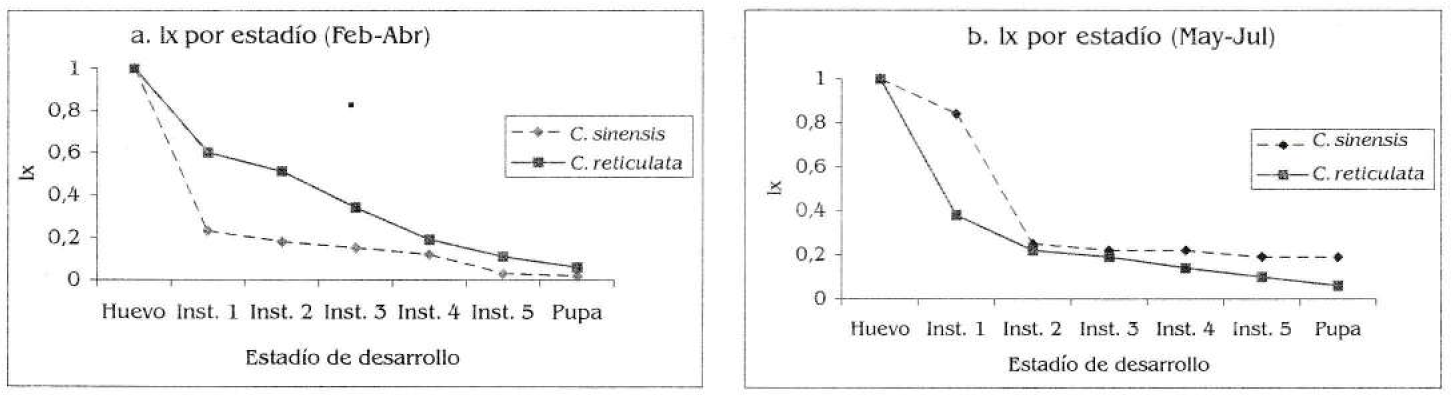
Probabilidad de supervivencia (lx) por estadío de desarrollo de Heraclides anchisiades anchisiades de acuerdo con la dieta. a) lx durante febrero-abril, (N= 122 para C. sinensis y N=183 para C. reticulata). b) lx durante mayo-julio (N= 63 para ambos hospederos).
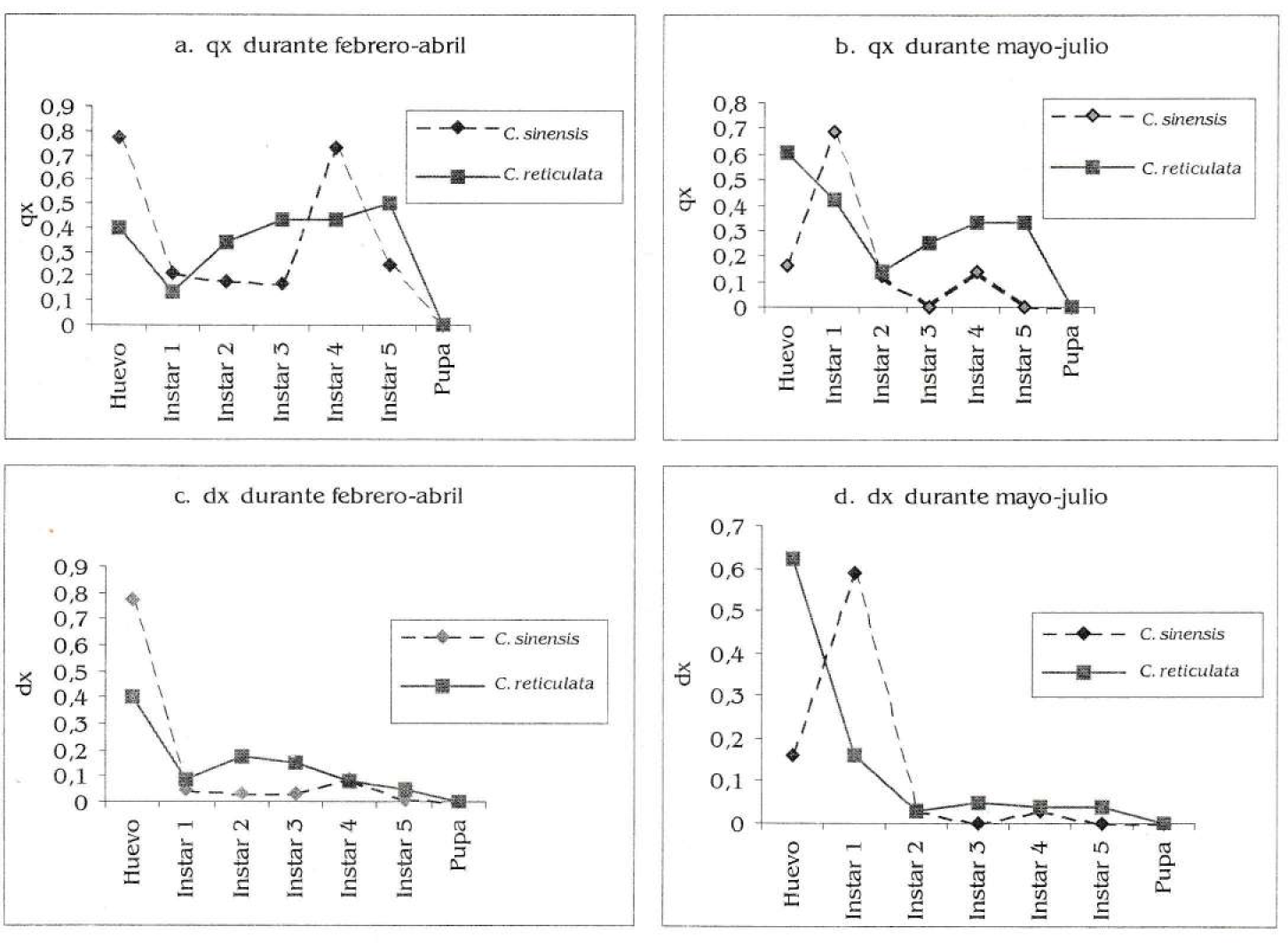
Mortalidad de Heraclides anchisiades anchisiades según hospedero. a) Específica por estadío (qx) durante febrero-abril. b). Específica por estadío (qx) durante mayo-julio. c). Respecto a la cohorte (dx) durante febrero-abril. d). Respecto a la cohorte (dx) durante mayo-julio.
No hubo diferencias significativas en los qx de cada dieta al compararse las dos épocas (Wicolxon para febrero abril, T=0.8438 y T=0.2188 para mayo julio; N=6 y p>0,05 en ambas pruebas; Figs. 5a y 5 b). En Citrus sinensis la mortalidad mayor fue en huevo (febrero - abril) e ínstar uno (mayo julio), mientras que en C. reticulata fue en el quinto ínstar (febrero - abril) y huevo (mayo - julio); aunque tampoco hubo diferencias significativas en los dx por época (Wicolxon, T=0.5626, N=6, p>0,05 para febrero abril y T=0.4376, N=5, p>0,05 para mayo julio; Figs. 5c y 5d) ni por dieta (Wicolxon, T=0.4376, N=5, p>0,05 para C. sinensis y T=0.8438, N=6, p>0,05 para C. reticulata).
Se encontró una disminución en la duración total del ciclo de desarrollo en ambas dietas de acuerdo con la época; en C. sinensis fue de 62 días en febrero abril y de 51 días en mayo julio; mientras que en C. reticulata fue de 66 días inicialmente y de 59 días en mayo julio (Figs. 6a, 6b, 6c y 6d); sin embargo, estas diferencias no fueron significativas al comparar las dietas por época (prueba de signos, n=5, p= 0.376, p>0,05 para ambas épocas) ni al comparar para cada dieta en particular según la época (n=5, p=2, p>0,05 para C. sinensis y n=4, p2, p>0,05 para C. reticulata). Se observaron diferencias significativas en el área foliar consumida entre dietas por estadio de desarrollo, y para cada dieta en particular según la época (Kolmogorov-Smirnov para dos muestras, Dmax=5, para todas las pruebas, Figs. 7a, 7b, 7c y 7d). El consumo promedio total de C. sinensis fue de 321,94 cm2/oruga en febrero-abril y 604,54 cm2/oruga en mayo - julio; para C. reticulata fue inicialmente de 309.76 cm2/oruga y 416.4 cm2/oruga en mayo julio.
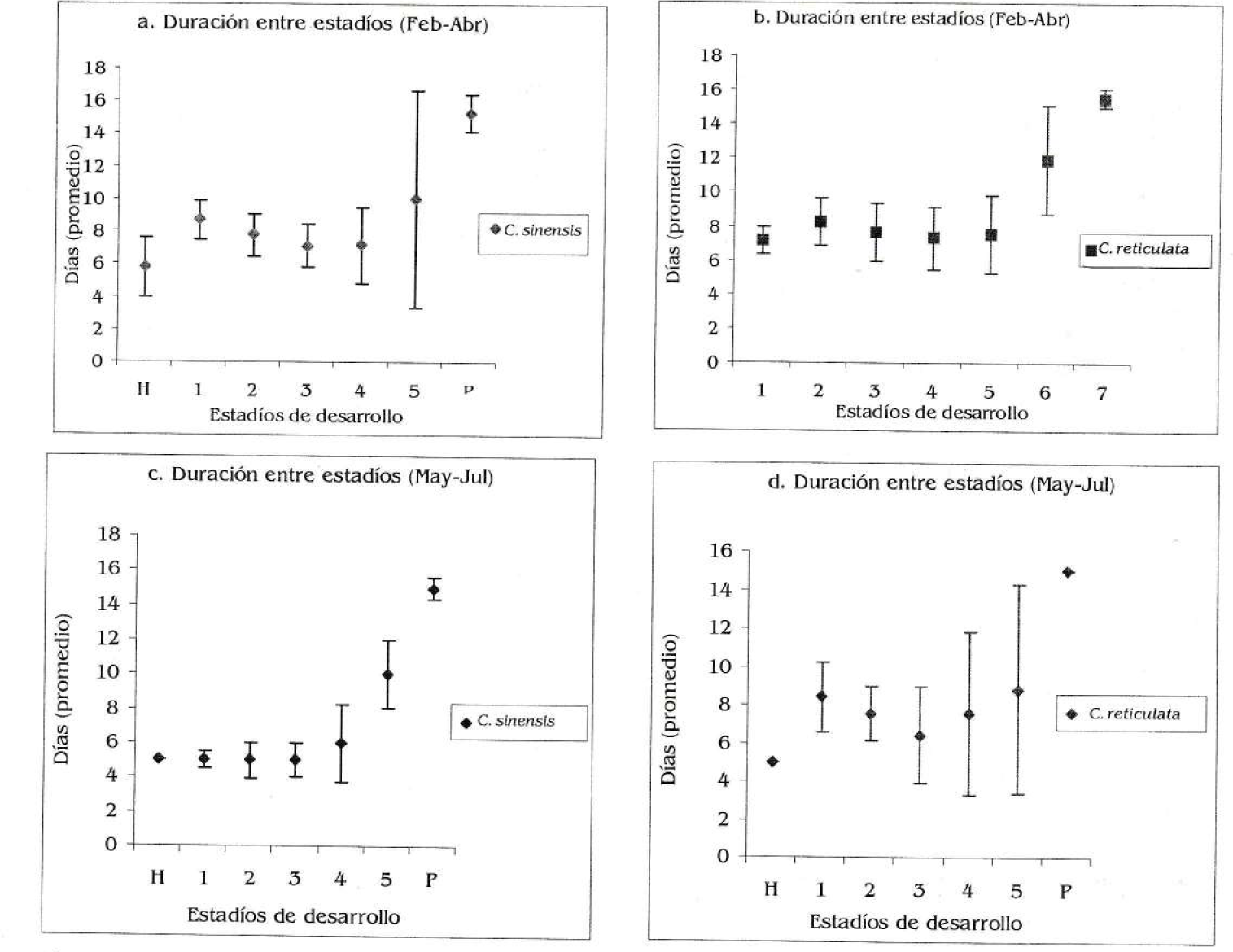
Tiempo de duración entre cada estadío de desarrollo de Heraclides anchisiades anchisiades de acuerdo con el hospedero (H=huevo, 1= Ínstar 1, 2= Ínstar 2, 3= Ínstar 3, 4= Ínstar 4, 5= Ínstar 5, P= Pupa). a) En Citrus sinensis durante febrero-abril. b) En Citrus reticulata durante febrero-abril. c) En Citrus sinensis durante mayo-julio. d) En Citrus reticulata durante mayo-julio.
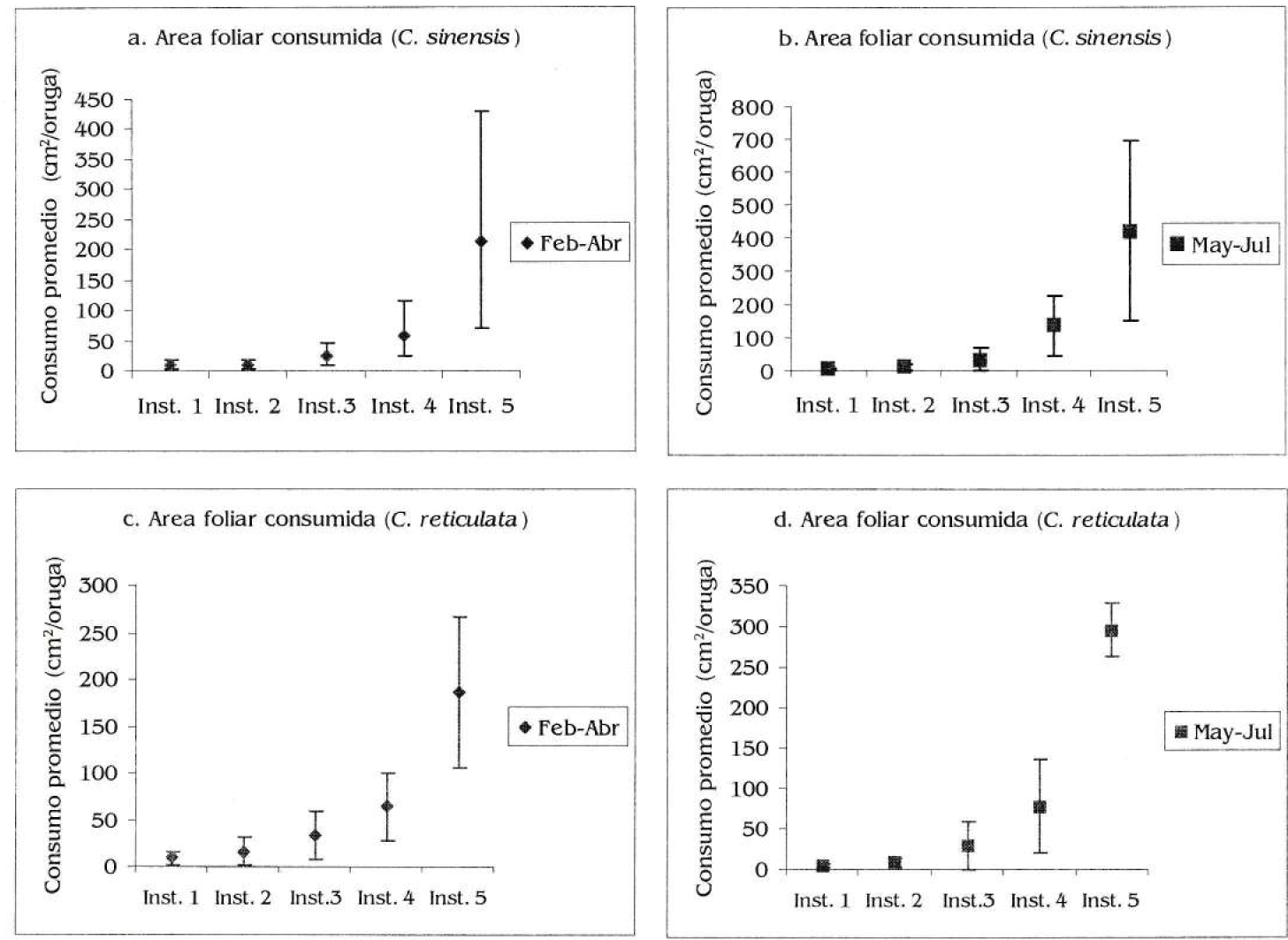
Area foliar consumida (cm2/oruga) por cada estadio de desarrollo de Heraclides anchisiades anchisiades. a) Consumo de Citrus sinensis durante febrero-abril. b) Consumo de Citrus sinensis durante mayo-julio. c) Consumo de Citrus reticulata durante febrero-abril. a) Consumo de Citrus reticulata durante mayo-julio.
Para evaluar si el proceso de cría de la mariposa tomada como modelo es sostenible, se realizó un análisis de factibilidad de acuerdo con criterios biológicos, económicos y sociales. Por una parte, el conocimiento requerido en la biología de las especies para que el proceso de cría sea exitoso, fue estudiado y analizado en su totalidad para la mariposa de estudio. Con la información obtenida fue posible generar propuestas para la optimización del proceso de cría. De otro lado, la comunidad, al estar directamente involucrada con el proceso de cría adquirió experiencia en el manejo de los huevos, orugas y adultos y participó activamente aportando ideas, muchas de las cuales fueron planteadas buscando directamente solucionar la alta mortalidad que se obtuvo en laboratorio.
Adicionalmente, se realizó un análisis TIR (tasa interna de retorno) para determinar la rentabilidad económica del proceso de cría de H. a. anchisiades en la zona de estudio. A partir del número promedio de huevos por postura, que fue de 60, y de la tasa de supervivencia optimizada al final del estudio (19%), por postura se obtendrían 11 adultos. Con base en este número y en la cantidad de posturas viables encontradas a partir de febrero hasta julio (11 posturas), se tiene que la producción sería de 121 mariposas por semestre, es decir, 242 mariposas anuales.
Para la cría de H. a. anchisiades en al zona de estudio no se requiere de la siembra de hospederos ni de la construcción de una sala de cría, por lo cual una familia de la comunidad fácilmente puede mantener dentro de su casa en un pequeño espacio los 80 vasos plásticos desechables que se necesitan para criar las 242 orugas y 5 jaulas de empupado, lo que constituye una inversión de $125.000. Los egresos están dados por la mano de obra de una persona, la cual es la encargada de manejar los huevos, las orugas, las pupas y los adultos, lo que se estima en $576.000 al año. Finalmente se estiman dos envíos anuales de mariposas desde la comunidad hasta la agencia compradora, lo cual totalizaría $648.936. De acuerdo con un análisis de mercado de esta especie el promedio en el precio de venta es de $5.335 por individuo.
Según el análisis económico del Humboldt (Moreno 1998), se estima que la agencia compra al productor cada mariposa por un 80% del valor final de venta (como es el caso de IFTA en Papua Nueva Guinea y el CRES en Costa Rica). Parsons (1995) calcula en un 75% este valor. Para este análisis se trabajó con el 75%, por lo que el precio de venta para el productor sería de $4000 por mariposa, que representarían $968.000 de ingresos para una producción anual de 242. La utilidad de la cría, representada por los ingresos que se obtienen menos los egresos, sería de $319.064 anual, sobre la cual se realiza un descuento del 12% efectivo anual (Tasa de interés), obteniendo así una utilidad neta de $280.776 (utilidad menos tasa de descuento). Con base en los resultados obtenidos de utilidad neta y la inversión, se realizó el análisis de rentabilidad (TIR) proyectado a 5 años (Tabla 3), obteniendo como resultado un 22.4%. Al ser el resultado obtenido en TIR mayor respecto a la inflación del país (estimada en 11.3% para diciembre del 2000, Banco de la República 1999) indica que la cría de esta especie es económicamente rentable.
Es importante resaltar que este resultado bueno se obtuvo con una tasa de supervivencia muy baja (19%) respecto a la óptima de cría (90%), y con una especie que tiene un precio bajo en comparación con las otras especies colectadas. Sin embargo, un proyecto de cría con fines comerciales no se puede sostener únicamente con una especie, por lo que en un futuro se debe incluir la construcción de una sala de cría en la cual se puedan mantener cómodamente los individuos de varias especies.
Análisis de tasa interna de retorno (TIR) calculada para 5 años de la cría de H. anchisiades anchisiades
En cuanto al análisis social, se encontró que los Nonuya tienen relaciones míticas ancestrales con las mariposas, y poseen historias para explicar su origen donde se ilustran las relaciones biológicas entre algunas especies con sus plantas nutricias y hospederas, sin embargo, no les consideran animales sagrados. Gracias a esto, no fue grande el impacto cultural; sin embargo, la aparición de las orugas, principalmente en agosto, corresponde a un período que ellos denominan como "época de gusanos", que relacionan como "enfermedad", razón por la cual al comienzo del proyecto se presentó un choque cultural fuerte, al considerar la cría de orugas como "cría de enfermedades". Dentro de esta concepción, el proceso de cría se fue asimilando paulatinamente con el transcurrir de la investigación, gracias a la participación activa de los indígenas en el proyecto, con lo que se generó un importante proceso de educación y sensibilización. Por otra parte, los indígenas conocen el uso potencial de las mariposas ornamentales, razón por la cual han manifestado su interés en estos proyectos, dada la necesidad de obtener fuentes de ingreso económicos especialmente para ayudar a solucionar en parte sus problemas de educación y salud.
Los dirigentes de la comunidad desean darle continuidad al proyecto de mariposas, desde un punto de vista cultural y occidental con fines comerciales. Para el trabajo de recuperación del conocimiento tradicional de las especies de mariposas, especialmente sus relaciones con plantas hospederas y nutricias, y la recolección de los nombres de las mariposas y orugas en idioma indígena, la comunidad ha planteado la ayuda de los conocedores Arturo Rodríguez y Rafael Mucutui (Baco). Esta recuperación del conocimiento es muy importante, no sólo a nivel cultural para la comunidad, sino también para el proyecto de cría de mariposas al contar con las ventajas que da el poseer un mayor conocimiento de la biología de las especies de interés.
Uno de los resultados más significativos del proyecto fue el hecho de que algunos indígenas, por iniciativa propia, se encuentran criando en cautiverio las orugas que colectan en campo, como es el caso de Ezequiel Ayarce quien actualmente está criando orugas encontradas en Caimo (Pouteria sp., Sapotaceae), Limón (Citrus limonia, Rutaceae), Guama (Inga sp., Mimosaceae), entre otras, además de continuar colectando mariposas.
Discusión
Los resultados obtenidos en cuanto a la presencia de especies de interés económico en la zona de estudio, soportan la elaboración de propuestas de cría de mariposas con fines comerciales. A pesar de que la colecta de mariposas no fue el énfasis de este trabajo, se obtuvo un buen resultado al comparar el número de especies con potencial económico (43 spp.) con el total de especies colectadas (61 spp.).
Lo encontrado para el ciclo de desarrollo y hospederos de H. a. anchisiades ratifican lo citado en la literatura (DeVries 1987; Tyler et al. 1994; Nishida 1995; Brown et al. 1995; Constantino 1998). Se confirma a las rutáceas C. sinensis y C. reticulata como hospederos de H. anchisiades (Brown et al. 1995). Estos hospederos se encuentran sembrados alrededor de las casas de los indígenas, en sitios abiertos, por lo cual es muy probable que la identificación de la hembra de estos hospederos sea visual, de acuerdo con lo planteado por Rausher (1978); además, estas plantas son cultivadas junto a varias especies de Inga (Mimosaceae), en las cuales se observó libando a las mariposas. Aunque en campo se identificaron y se realizó el seguimiento a tres especies más de rutáceas, Citrus limon, C. limetta y Zanthoxylum sprucei, solo en C. limon, registrada también como uno de los hospederos principales (Brown et al. 1995; Constantino 1998), se encontraron orugas gregarias de tercer y quinto ínstar. Esta cantidad baja de orugas puede indicar que no es un hospedero frecuentemente utilizado por las mariposas de esta zona.
Por otra parte, la ubicación de las posturas en hojas jóvenes y medias jóvenes pueden deberse a que este tipo de hojas poseen una mayor concentración de nutrientes en sus tejidos (nitrógeno, potasio, fósforo, azucares) y una menor cantidad de celulosa, por lo cual la calidad nutricional sería mejor (Rausher 1980). Según Loader y Damman (1991), una baja calidad nutricional de la hoja reduce la sobrevivencia de herbívoros de manera indirecta al incrementar el período de exposición a enemigos naturales, por lo cual, para incrementar la supervivencia de las orugas, resulta más ventajoso colocar la postura sobre tejidos jóvenes. Es importante destacar que también la composición química de las plantas es un factor determinante en el uso del hospedero, como se ha encontrado en las especies de Papilio (Berembaum y Feeny 1981; Lindroth et al. 1988, citados por Nitao et al. 1991).
La ubicación de las posturas en el envés de las hojas puede deberse a la gran presión de parasitoides Chalcidioideos (para los huevos) y depredación por Mirmicinos (Fig. 3a); colocar los huevos en esta posición puede reducir la posibilidad de detección de los depredadores. Las diferencias en el número de huevos por postura (Fig. 3b), mayor en C. sinensis, pueden atribuirse a que los árboles de esta especie son menos frondosos, lo que facilitaría la detección a los parasitoides; tener un mayor número de huevos por postura incrementaría el éxito de eclosión de, al menos, unos pocos individuos (Singer 1984). Los árboles de C. reticulata, por el contrario, poseen un mayor número de hojas lo cual reduce la probabilidad de encuentro con parasitoides, pero esta misma característica pudo influir en el alto número de posturas encontradas con hongos (Fig. 3a), los cuales estarían favorecidos al ser el árbol tan denso, y por ende, tener unas condiciones de mayor humedad.
Dada la alta mortalidad de posturas por enemigos naturales se recomienda, para optimizar la cría, colectarlas una vez sean depositadas en el hospedero, reduciendo así la probabilidad de ataque de parasito ides, patógenos y depredadores y aumentando el número de mariposas que se obtendrán para la venta. En campo no se observó depredación sobre las orugas, probablemente debido a compuestos tóxicos (como furanocoumarinas, limonoides, enzimas antioxidantes y alcaloides del grupo benzil-isoquilonina, presentes en los tejidos foliares de las rutáceas y que, según Berembaum en 1983, son acumulados por las orugas de papiliónidos) lo que les puede conferir protección química y por consiguiente, ser rechazados como alimento por parte de depredadores como aves, mamíferos, reptiles o insectos. Además, se observó que cuando las orugas se sienten atacadas, expelen por su boca un líquido verdoso de olor desagradable y proyectan osmaterios, también de olor desagradable; dada la condición gregaria de las orugas, el olor se hace más intenso, lo que puede disuadir del ataque a sus posibles depredadores (Ford 1957; citado por Amarillo 1993).
Se encontró una gran variación en la sobrevivencia de la mariposa de acuerdo con el hospedero, lo cual puede estar incidiendo en su selección y utilización. Inicialmente, la tasa de supervivencia fue mucho menor en C. sinensis (Fig. 4a), por lo que se consideró que su utilización como hospedero era un probable error de selección por parte de la hembra, razón por la cual se realizó la prueba de dieta. Sin embargo, en ésta la supervivencia en C. sinensis aumentó, mientras que en C. reticulata se mantuvo igual (Fig. 4b) (es importante tener en cuenta que las larvas sometidas a esta prueba se obtuvieron a partir de una postura encontrada sobre C. sinensis, lo cual pudo favorecer que bajo esta misma dieta tuvieran una supervivencia mayor que los alimentados con C. reticulata).
Dados estos resultados, se puede descartar la probabilidad de error en la selección de hospedero, debido a que las dos rutáceas son igualmente óptimas para la especie de acuerdo con el fenotipo de la hembra; uno estaría mejor adaptado al recurso C. sinensis y otro a C. reticulata, por lo cual se recomienda, para la cría de esta especie, alimentar a las orugas con material vegetal proveniente de los hospederos en donde se encontró la postura. Sin embargo, este aumento en la supervivencia de las orugas en C. sinensis (su hospedero) y la reducción del período de desarrollo que se obtuvo durante la prueba, pudo deberse también a que inicialmente se colocaron las orugas solitarias, mientras que durante la prueba se colocaron tres individuos por vaso, lo cual les habría permitido optimizar el consumo de las hojas de C. sinensis, las cuales son más coriáceas que las de C. reticulata. Ghent (1960) encontró que los estadios más tempranos de Neodiprion pratti tenían dificultad al tratar de masticar las hojas de los pinos individualmente mientras que fueron mucho más exitosas cuando se alimentaron en grupo; esto podría aplicarse también, de acuerdo con lo observado, a las orugas de H. a. anchisiades.
La decisión de colocar tres individuos por vaso se tomó basándose en las observaciones de campo, donde la condición de ser gregario permanece hasta 5º ínstar, por lo que se consideró que el hecho de colocar un individuo por vaso pudo influir en la mortalidad alta encontrada inicialmente. Aunque lo ideal para realizar la prueba de dieta hubiera sido someter a las orugas a esta misma condición de individualidad, el objetivo principal del estudio del ciclo de desarrollo con fines comerciales está dirigido a aumentar la supervivencia de la población, para obtener una mayor cantidad de adultos como producción del sistema de cría. Dado lo anterior, se buscó la forma de aumentar esta supervivencia de acuerdo con las observaciones comportamentales de la especie en campo, por lo que se estimó necesario mantener el "gregarismo" durante el estadio larval.
De acuerdo con Chew y Robbins (1984), el comportamiento gregario de las orugas está determinado en gran parte por el tipo de posturas, y en este caso, siendo las posturas y larvas de H. a. anchisiades agregadas, es probable que esta condición le represente ventajas. Según Stamp (1980) hay ventajas tanto para las hembras, como para los huevos y las orugas. Para las hembras, el colocar posturas gregarias puede ser ventajoso cuando los recursos alimenticios de los adultos se encuentran lejos de los hospederos, o si estos se encuentran distribuidos en parches, o si las poblaciones de adultos están muy dispersas; en cualquier caso, las hembras necesitan gastar mayor tiempo localizando el hospedero o buscando pareja. Sin embargo, los hospederos de H. a. anchisiades se encontraban a corta distancia y junto a especies de Inga en las cuales se observaron libando, por lo cual se descartan las dos primeras posibilidades, sugiriendo que la condición de gregarismo en las posturas de esta especie estaría asociada más a disminuir el ataque por parasitoides, ya que los grupos de huevos alcanzan tasas de parasitoidismo más bajas que los huevos solitarios (Emlen 1973). Otra ventaja para los huevos en grupos es que así se reduce la cantidad de superficie del huevo expuesta a las condiciones ambientales, disminuyendo la posibilidad de desecación. Es importante anotar que las posturas gregarias no son frecuentes dentro de la familia Papilionidae (Stamp 1980).
La supervivencia de las orugas gregarias, especialmente recién eclosionadas, puede incrementarse debido al comportamiento termoregulatorio conjunto (Seymour 1974). Además, la tasa de crecimiento por individuo puede ser más rápida en las orugas que se alimentan, descansan y mudan sincrónicamente en comparación con las solitarias (Long 1953, 1955). Long (1953) encontró que las orugas gregarias son 4 veces más activas, gastan un 25% más de tiempo alimentándose, poseen un contenido más alto de grasa, y empupan más pronto que una oruga solitaria. Además, también les puede facilitar su alimentación y consecución de alimento. Al comparar la duración del ciclo de desarrollo en larvas criadas individualmente y en grupo (Figs. 6a, 6b, 6c y 6d), se observó una reducción tanto para los conjuntos de orugas alimentadas con C. sinensis como en C. reticulata; esta reducción en las orugas agrupadas puede relacionarse con su comportamiento gregario. También se observó que la tasa de herbivoría total por individuo fue mucho mayor para ambos hospederos cuando se dejaron tres orugas por vaso (Figs. 7a, 7b, 7c y 7d). Es importante destacar que, pese a la mayor coriacidad de las hojas de C. sinensis, el consumo de las mismas siempre fue mayor, tanto en orugas solitarias como gregarias, en comparación con el consumo de C. reticulata. Esto podría asociarse con una con menor calidad nutricional de sinensis; en donde las orugas se verían obligadas a consumir una mayor cantidad de biomasa para completar sus requerimientos.
Dados estos resultados, que apoyan las ventajas de mantener las orugas gregarias, se recomienda para el proceso de cría mantener esta condición, con el fin de reducir el tiempo de su ciclo de desarrollo, optimizar su consumo y aumentar la tasa de supervivencia. Otra ventaja muy importante desde el punto de vista económico, que se obtiene al colocar más orugas por vaso, es que se reduce el tiempo y materiales invertidos en el cuidado de las mismas, minimizando los costos.
De acuerdo con los análisis de mortalidad (qx y dx), los estadios que presentaron una mayor mortalidad son los más tempranos (Figs. 5a, 5b, 5c y 5d), por lo que en éstos se debe tener mayor precaución en el manejo y cuidado, ya que es este estadio los individuos son más frágiles y susceptibles a bacterias, que fueron la única causa de mortalidad de las orugas en condiciones de laboratorio. Dado lo anterior, se recomienda realizar la limpieza diaria del vaso donde permanecen, el cual debe ser cambiado periódicamente, siendo esto aún más indispensable cuando una de las orugas muere, con el fin de evitar el contagio de las demás. Se sugiere aislar las orugas enfermas de los cuartos de cría.
Con base en la información generada de la biología de H. a. anchisiades se pueden realizar otras pruebas en cuanto a calidad nutricional del hospedero (análisis de esclerofilia, de contenido de nitrógeno y metabolitos secundarios) y número de individuos que se pueden mantener por vaso plástico, con el fin de optimizar aún más la cría desde el punto de vista económico al obtener mayor cantidad de individuos para la comercialización.
Por otra parte, de acuerdo con las características de la zona de estudio, de la Comunidad y los resultados obtenidos en la biología de la especie, el sistema de cría recomendado para Peña Roja es el de rancheo. Según Ramírez (1994), las estrategias para el manejo sostenible de poblaciones silvestres deben estar basadas en la preservación de la máxima biodiversidad ecológica y ecosistémica en la región, lo cual se cumple con este sistema de cría. Además, con el rancheo se tiene la ventaja de reducir la mortalidad en las posturas al colectar las mismas y protegerlas del ataque de los enemigos naturales, lo cual fue la mayor causa de mortalidad poblacional. Se estima que en la naturaleza entre el 2% y el 5% de los huevos llegan a adultos debido a enemigos naturales (Tyler et al. 1994), pero con rancheo se puede lograr que entre 85 y 95% lleguen a la madurez (Constantino 1996), generando suficientes individuos para ser liberados al ecosistema. Otra ventaja adicional para la cría de esta mariposa en la zona es que no se necesita sembrar hospederos ya que estos se encuentran cultivados alrededor de las casas de los indígenas, por lo cual la recolección de posturas puede hacerse en menor tiempo.
Adicionalmente, el tiempo y esfuerzo invertido en la cría de las orugas es comparativamente menor respecto a cualquiera de los otras ofertas de trabajo remunerado presentes en la zona de estudio, como lo son la extracción de oro y la tala de bosque para cultivos ilícitos, obteniéndose beneficios superiores a nivel económico y para el ecosistema.
El producto a comercializar sería el de mariposas preservadas, ya que un mercado de pupas vivas es insostenible en la actualidad, dada la irregularidad de los vuelos en la zona, entre otros factores. Este hecho no permite tener ninguna seguridad de que las pupas lleguen a tiempo a su destino antes de la emergencia del adulto. Adicionalmente, la comunidad no cuenta con energía eléctrica, necesaria si se pretende establecer el mercado de pupas vivas para mantenerlas a bajas temperaturas en diapausa. Además de esto, en la zona se presentan otros problemas que deben tenerse en cuenta al momento de iniciar el proceso de comercialización, como lo es la presencia de guerrilla e incluso factores ambientales como la lluvia o la baja en el nivel del río, los cuales afectan el transporte de la comunidad a donde opera la pista de aterrizaje (distante tres horas en bote de Peña Roja).
Es importante destacar que, aunque las comunidades indígenas consideran que la valoración de los recursos en la economía de mercado subestima el verdadero valor de los mismos, la Comunidad de Peña Roja ha estado más abierta y receptiva a la información de los planes de manejo de la vida silvestre con fines económicos, por lo cual conocen los usos actuales o potenciales de los recursos provenientes de la biodiversidad en general (Instituto Humboldt 1999).
Adicionalmente, se resalta la importancia de tener en cuenta los patrones culturales de la comunidad indígena respecto a sus relaciones y acciones sobre los animales para evitar el riesgo de generar impactos culturales desde el momento de la formulación de un proyecto productivo como el de mariposas. Durante el trabajo con esta comunidad, se lograron algunos acercamientos, comenzando desde un cambio en la concepción de los indígenas que consideraban la cría de orugas como cría de enfermedades. En un comienzo esta visión se presentó como un obstáculo a nivel cultural para el desarrollo del proyecto; otro inconveniente fue la tradición de no sacrificar a ninguno de los animales criados; sin embargo, a lo largo de la investigación la comunidad siguió muy atenta el desarrollo de la misma, con lo que se logró involucrarla, teniendo así que en la actualidad son los indígenas de Peña Roja quienes, por iniciativa propia, se encuentran pendientes de los sitios de oviposición de otras especies de mariposas, llegando incluso a comenzar crías experimentales con las orugas que encuentran en campo durante sus actividades diarias.
Para iniciar el proyecto de cría con fines comerciales se deberá estudiar la biología de otras especies de mariposas de interés económico, de las cuales, en el presente proyecto, se obtuvo una lista preliminar. Dentro de estas se colectaron orugas de Colobura dirce en yarumo blanco (Cecropia ficifolia), una planta común en la zona. Algunas mariposas pueden ser estacionales, por eso es conveniente estudiar y producir mariposas de otras especies para que a lo largo del año se tenga una oferta constante. Esto último ha sido planteado por la comunidad, que está interesada en continuar con el proyecto, y seguir estudiando la biología de otras especies, para lo cual los indígenas ya conocen la metodología a seguir y esto, sumado a sus conocimientos tradicionales de los eventos de las relaciones hospedero-mariposas, hace más factible continuar el proceso de cría. Para esto se debe considerar la construcción de una sala de cría para mantener y manipular cómodamente varios individuos y especies.
Finalmente, dado el interés de la comunidad en estos proyectos productivos, puede plantearse el estudio de ciclos de vida de otras especies animales con fines económicos, como coleópteros, que tienen una gran demanda, en especial de las familias Carabidae, Buprestidae, Elateridae, Curculionidae, Scarabidae, Cerambycidae, Cincindelidae, Tenebrionidae, Nitidulidae, Sylphidae, Meloiidae, Cleridae, Passalidae, Lymexiloniidae y Melyridae, todas bien representadas en la Amazonía Colombiana.
Footnotes
Agradecimientos
Damos agradecimientos a la Comunidad Indígena de Peña Roja por su hospitalidad, colaboración e interés aportado desde siempre al proyecto, en especial a Ezequiel Ayarce (Chaqué). A la Fundación Tropenbos por el apoyo logístico y financiero, especialmente a su director Carlos Rodríguez. A Jaime Alberto Álvarez, Gonzalo Andrade-C. y Luis Miguel Constantino por su valiosa ayuda en la determinación de algunas mariposas. Al Herbario Amazónico COAH del SINCHI, especialmente a su director Dairon Cárdenas, por su colaboración en la determinación y montaje del material vegetal colectado.