
Abstract
Compsus n. sp. was recorded for the first time in 1939 associated to citrus crops in Colombia. Since 1995 populations of this insect has increased causing economic losses on citrus crops in Quindio and Tolima. To contribute to the knowledge of this species a study was conducted in citrus plantations in Montenegro, Quindío (1250 m.a.s.l., 21.9°C, 78% R. H., 2026 mm) and Armero Guayabal, Tolima (350 m.a.s.l., 28.5°C, 62% R. H., 1400 mm). The life cycle, behavior, natural enemies and population fluctuation through time, were studied at these sites. Compsus n. sp., completed its life cycle from 132 to 219 days. Adult females oviposit irregular egg masses on the aerial part of the trees; neonates larvae fall to the soil and bury into the ground were first instars feed on small adventitious roots of the tree. Later on bigger larvae start feeding on more developed roots. At the end of its developmental stage they build a pupal camera in the ground and pupation period takes about one to two months. Then adult emerges and the life cycle goes on. Twenty-five natural enemies were recorded, which confirms that this weevil has a good number of organisms regulating its populations.
Keywords
Introducción
En cultivos de cítricos de los departamentos del Quindío y el Tolima se detectó la presencia del picudo de los cítricos Compsus pos. n. sp. causando daños en el sistema radical y follaje (Osorio 1997). Actualmente se ha detectado su presencia en nueve departamentos, incluyendo la zona central cafetera (Sánchez 2000), y representa una amenaza potencial a la sanidad de la citricultura en Colombia, por la capacidad destructiva de esta especie y por el manejo inadecuado que se ha efectuado para reducir sus poblaciones
En diversas zonas citricolas de América se encuentran complejos de picudos (Coleoptera: Curculionidae) (Montejo y Montes s.f) que incluye los géneros: Artipus, Cleistolophus, Compsus, Diaprepes, Epicaerus, Exophalmus, Lachnopus, Litostylus, Pachnaeus, Pantomorus y Tanymecus, (Woodruff 1985), Otiorrhychus y Asynonychus (Davies y Albrigo 1994). Estas especies se comportan como plagas dobles ya que en sus estados de larva y adulto realizan daños.
El género Compsus es conocido comúnmente en Colombia como vaquita (Posada 1989) o burrita de San José (Cárdenas y Posada, 2001). Fue registrado en cítricos en Colombia desde 1936⁵; a pesar de ello no existe en la literatura antecedente de estudios sobre biología, comportamiento y enemigos nativos de este género en Colombia ni en el mundo.
La especie que está afectando los cítricos fue identificada como C. viridilineatus Jekel por el Instituto Colombiano Agropecuario (ICA) en Tibaitatá, por comparación con una especie clasificada en 1938 por L. L. Buchanam en el Departamento de Agricultura de los Estados Unidos (USDA) 5 . Posteriormente, la especie C. viridilineatus fue denominada como un "nomen nudum", es decir un nombre publicado sin la suficiente información descriptiva para satisfacer los criterios de disponibilidad o de publicación 6 , por esta razón se denomina pos. n. sp.
BENAVIDES, M. 1998. Colección taxonómica “Luis María Murillo” Tibaitatá, ICA.
PEÑA, J. E. 1998. Comunicación escrita de la nota aclaratoria de la identificación del Dr. Charles O'Brien.
El enfoque de manejo dado a este insecto en algunos huertos de la zona central cafetera ha sido inadecuado y ha obedecido al desconocimiento de aspectos básicos como biología, hábitos y enemigos nativos sin los cuales es imposible establecer cualquier medida racional de control. El objetivo de esta investigación fue contribuir con el conocimiento de esta especie, para la futura difusión de estrategias de manejo integrado del picudo de los cítricos.
Materiales y Métodos
La investigación se desarrolló en el Centro Nacional de Investigaciones de Café, Cenicafé y en tres fincas de producción comercial (Tabla 1). Los ensayos de laboratorio se realizaron en la finca La Concha, con condiciones de temperatura y humedad relativa promedio de 21.7° C y 89%, respectivamente.
Condiciones de los sitios donde se llevó a cabo la investigación
Ciclo de vida
Para el establecimiento de la cría artificial del insecto se colectaron adultos de ambos sexos en la parte aérea de los árboles, se confinaron en cajas transparentes de 25×30×10 cm en grupos de 200 individuos en una proporción de 1:1. Se alimentaron con hojas frescas de cítricos, como sitios de oviposición, se colocaron tiras dobles de plástico transparente en el interior de las cajas, de acuerdo con la metodología de Pérez 7 . Las posturas se recogieron diariamente y se llevaron a cajas plásticas de 18×12×7 cm con papel absorbente hasta evidenciar la eclosión (Modificaciones a la metodología de Beavers 1982). Se seleccionaron 10 cajas con posturas y se observaron diariamente para registrar la emergencia, tamaño y los cambios sufridos.
PÉREZ MYRIAM. 1998. Comunicación personal acerca de cría del picudo. Cenicafé, Chinchiná.
Las larvas se criaron utilizando dos metodologías: una de campo y otra de laboratorio que permitieran medir las variables de crecimiento durante el primer mes de desarrollo (Tabla 2). En la cría en laboratorio se emplearon seis cubetas de plástico de 57×35×5,5cm divididas en 84 celdas. En cada celda de 137 ml se colocó una porción pequeña de zanahoria y 50 larvas neonatas (0-96 horas obtenidas de los criaderos artificiales) como lo sugiere Peña (1998). A los cinco días de la infestación artificial se iniciaron las evaluaciones (muestreo destructivo), tomando el número de celdas necesarias para recuperar como mínimo 30 larvas; el suelo se revisó por medio de cedazos de cocina. Se midieron las variables longitud, ancho y diámetro de la cápsula cefálica (dcc) de las larvas.
En el campo se utilizaron 150 plantas de naranja valencia sobre citrumelo 4475 y 20 de yuca sembradas en bolsas plásticas de vivero (30″ × 18″); en cada bolsa se realizó una infestación artificial en la superficie del suelo con 300 larvas neonatas.
Después de un mes de la infestación en campo se realizó la primera evaluación (muestreo destructivo), que consistió en tomar tres plantas de cítricos cada siete días y cada 15 una de yuca; el suelo de cada bolsa se pasó por cuatro zarandas (malla de 3×3, 5×5, 7×7, 8×8) y un tamiz estándar No.12, las raíces se revisaron minuciosamente para recuperar el mayor número de larvas posible. Los estados colectados se colocaron en cajas plásticas con suelo, luego se trataron con agua caliente y solución Kalhes, posteriormente se registró la edad y se midieron las mismas variables que en el laboratorio.
Las evaluaciones para determinar la duración de los estados subterráneos se realizaron hasta que se observó emergencia de los adultos y/o cuando se terminaron las unidades experimentales. Para determinar la longevidad de los adultos, periodo de preoviposión, capacidad y distribución de oviposición se colectaron adultos recién emergidos en campo por medio de la instalación de trampas de emergencia de 50x60x10 cm (0.3 m2 de suelo), hechas con madera, malla 3×3 y cuatro varillas, ya que en la cría artificial no se pudo obtener este estado.
Las hembras colectadas se llevaron al laboratorio donde se confinaron en cajas con tiras dobles de plástico transparente en el interior, hojas de cítricos y un macho colectado del campo. Las cajas se revisaron cada 24 horas para registrar la aparición de la primera postura y así se contabilizó el período de preoviposición, luego se revisaron dos veces por semana para llevar el registro de longevidad y el número de huevos por hembra determinando con ello la capacidad y distribución de la oviposición.
Los machos, encontrados en las trampas de emergencia, se confinaron en frascos de boca ancha y cada semana se contó el número de individuos vivos para determinar el tiempo total del estado adulto.
Las variables de tamaño de todos los estados se midieron bajo estereoscopio provisto de micrómetro ocular, los datos obtenidos se sometieron a análisis de estadística descriptiva que incluye: promedio, desviación estándar, rangos, coeficiente de variación y límites de confianza del promedio.
El periodo de incubación, además de evaluarlo en condiciones de laboratorio, se determinó en campo utilizando mangas entomológicas de 90 cm de longitud y 40 cm de diámetro forradas con muselina, las cuales se ubicaron en una rama de la parte inferior de 30 árboles de naranja valencia de dos años de edad. A la mitad de las ramas seleccionadas se les colocaron cuatro tiras plásticas dobles de 2.5 x 30 cm como sitios de oviposición.
En cada una de las ramas cubiertas con las mangas se infestó artificialmente con adultos colectados del campo en diferentes proporciones (♀:♂): (0:5, 0:10, 0:20, 1:1, 1:2, 1:5, 2:2, 2:3, 3:3, 5:5, 20:20; 40:40). 24 horas después de la infestación artificial se revisaron cada una de las hojas y las tiras plásticas contenidas en la manga para detectar posturas, se marcaron y se retiraron los adultos. Las posturas se revisaron diariamente hasta evidenciar eclosión.
Estudio del comportamiento
El comportamiento de la población de los adultos de Compsus pos n. sp. en el tiempo y su relación con las condiciones climáticas y la humedad del suelo, se realizó en un lote comercial de 200 árboles de naranja valencia (5600 m2) por 33 semanas. En el lote se marcaron los árboles con el número de surco y el número de fila, y semanalmente se seleccionó al azar el 10% de los árboles, se les realizó el monitoreo y se recogieron los adultos que caían al suelo. El monitoreo a los árboles se hizo sacudiendo ocho ramas durante 10 segundos sobre una superficie negra de plástico de 5 × 5 m con un vibrador mecánico accionado neumáticamente por un compresor portátil con el fin de obtener vibraciones constantes de las ramas en cada árbol.
Los datos meteorológicos de precipitación, humedad relativa, brillo solar y temperatura se obtuvieron de la estación climática de El Agrado del Comité de Cafeteros del Quindío. El balance hídrico se calculó con la precipitación y evapotranspiración de la zona, la capacidad de retención de humedad de la unidad Montenegro y la evapotranspiración potencial de Penman, por medio del programa elaborado en Cenicafé.
Las variables que se midieron con relación al insecto fueron: número de adultos hembras, machos y totales por árbol en cada semana de lectura; estas variables se analizaron por medio de estadística descriptiva. La asociación del número promedio semanal de adultos por árbol con los factores climáticos y el balance hídrico (exceso, déficit e índice de humedad del suelo) se analizó por medio de correlaciones.
Para determinar si el picudo de los cítricos era atraído a la luz artificial ó semioquímicos, se hizo un estudio exploratorio de atracción de los adultos de Compsus pos n. sp. a trampas de luz artificial y cebadas con diferentes materiales. En este ensayo se evaluaron trampas con lámparas de luz blanca y negra y una trampa tipo "Pitfall" o de caída, colocadas en un lote comercial de naranja valencia sembrada a una distancia de 5m × 7m. La trampa de caída se diseño con material de reciclaje (tarros con tapa de 12 cm de diámetro y 20 cm de profundidad) (Fig. 1), se ubicaron a 20 cm del tronco (Fig. 1c, d) cada cinco árboles dentro del surco (25 m) y separadas cada tres calles (21 m).

Trampa pitfall construida de material de reciclaje para evaluar atracción de Compsus pos. n. sp. a semioquímicos. a. Trampa utilizada cuando los atrayentes eran adultos del picudo. b. Trampa utilizada para el resto de atrayentes. c. d. Instalación de la trampa pitfall en campo.
Los atrayentes utilizados fueron: hojas tiernas de naranja valencia, frutos de naranja valencia descompuestos, proteína hidrolizada al 10%, excremento sólido del picudo obtenidos de la cría en laboratorio, alcohol, alcohol-etanol; hembras, machos y parejas del picudo. Se cebaron individualmente cada 14 días y se evaluaron semanalmente durante un mes. Cuando las trampas se colocaron por primera vez o cuando se cebaron, se sacudían todos los árboles del lote para que los adultos del picudo cayeran al suelo. En cada evaluación se registraron las capturas de insectos en las trampas semanalmente.
Las trampas de luz se ubicaron en una rama cerca del tronco a 1 m del suelo, distanciadas 50 m entre árboles y 28 m entre surcos; en la parte inferior de las trampas se colocó una bolsa plástica con agua para que los insectos atrapados no pudieran escaparse, la bolsa se revisó dos veces por semana y los insectos atrapados se recuperaron por medio de un tamiz (Fig. 2).

Trampas de luz utilizadas para evaluar atracción del picudo de los cítricos.
Detección e identificación de enemigos nativos
Se realizaron visitas mensuales a las fincas comerciales, colectando los estados del picudo que se observaban con síntomas de enfermedad o parasitismo. Además se estuvo atento a detectar algún tipo de predación.
Las observaciones se hicieron arrancando árboles desde la raíz posiblemente afectados por el picudo y con observaciones en la parte aérea con altas poblaciones del insecto. Los estados del insecto enfermos se ubicaron en cámaras húmedas, luego se realizó el aislamiento y posteriormente la identificación de los entomapatógenos. Los individuos con síntomas de parasitismo se colocaron en frascos de emergencia, los parasitoides encontrados se identificaron hasta género por expertos. Los depredadores colectados se identificaron hasta familia.
Resultados y Discusión
Establecimiento de la cría artificial
La metodología utilizada para obtener huevos y larvas de primer ínstar fue satisfactoria y resulta útil para el aumento del pie de cría de los parasitoides de huevos y la obtención de larvas neonatas con el fin de evaluar entomopatógenos. Se obtuvieron 105.000 larvas necesarias para realizar las infestaciones artificiales para el estudio del ciclo de vida. Con 15.200 parejas se lograron 148.860 huevos en 12 días, en promedio cada pareja por día ovipositó 9.0 ± 2.7 (X ± de) huevos.
La cría de larvas no fue exitosa ya que se desarrolló en condiciones muy artesanales y en un sitio no adecuado (cuarto en finca comercial) que estaba cerca a aspersiones periódicas de Beauveria bassiana. Entre los inconvenientes se señalan: el escape de las larvas que se encontraban confinadas, alta mortalidad causada por B. bassiana y presencia de hongos, bacterias y ácaros contaminantes.. Sólo llegó al estado de pupa el 0.4% de la población inicial y no fue posible obtener adultos.
Descripción de los estados del insecto
Los adultos de Compsus pos. n. sp. son generalmente de color blanco aperlado, presentan élitros esculpidos con costas y franjas longitudinales que varían desde verdes, azules hasta marrón iridiscentes; en las patas, placas ventrales, rostrum y pronoto se observan manchas de igual color a las franjas de los élitros (Fig. 3a). Los adultos recién emergidos del suelo presentan élitros blandos y mandíbulas grandes (Fig. 4d). El dimorfismo sexual en el adulto es marcado en el último uroesternito. También el tamaño de la hembra (11.76 ± 0.73 mm de longitud) sobre el macho (9.81 ± 0.5) (Fig. 3).
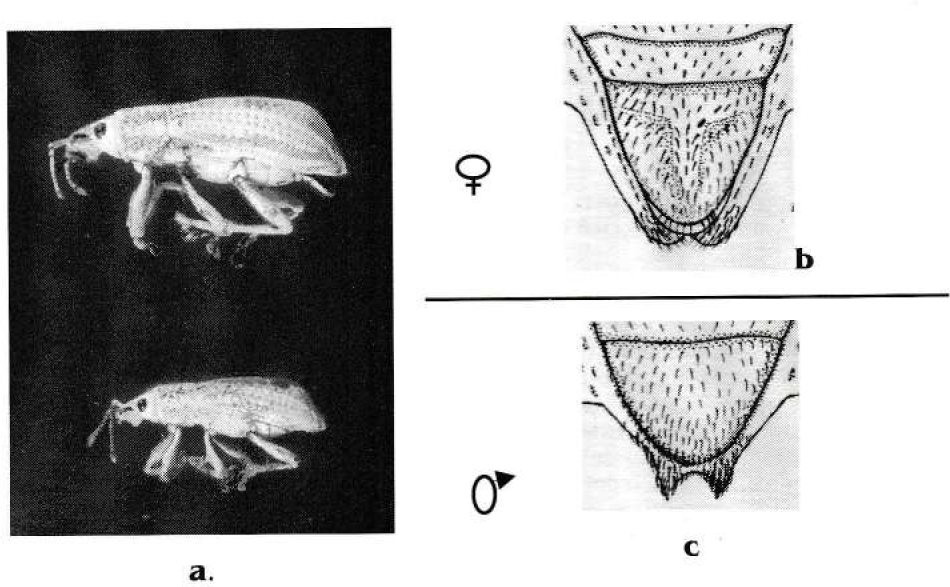
Dimorfismo sexual de Compsus pos n. sp. a Diferencias en el tamaño. b - c Diferencias en el último esternito, en las hembras el número de setas es mayor y su ápice es menos ancho que en los machos.
Los huevos son de forma ovoide, alargados, lisos (Fig. 4a) y miden 1.2 ± 0.05 mm por 0.37 ± 0.26 mm; son ovipositados en masas irregulares de una sola capa (Fig. 4b), el número de huevos por postura es muy variable (Tabla 3). Las larvas son ápodas y eucéfalas de color crema con cabeza muy esclerosada y mandíbulas bien desarrolladas, de color carmelita, su cuerpo es subcilíndrico y ligeramente arqueado. Las de primer ínstar miden 0.84 ± 0.24 mm y tienen 0.2 ± 0.01 mm de diámetro de la cápsula cefálica (Fig. 4c), durante su desarrollo el color es sub-hipodermal y alcanzan a medir hasta 15.35 mm de longitud y 2.4 mm de dcc.
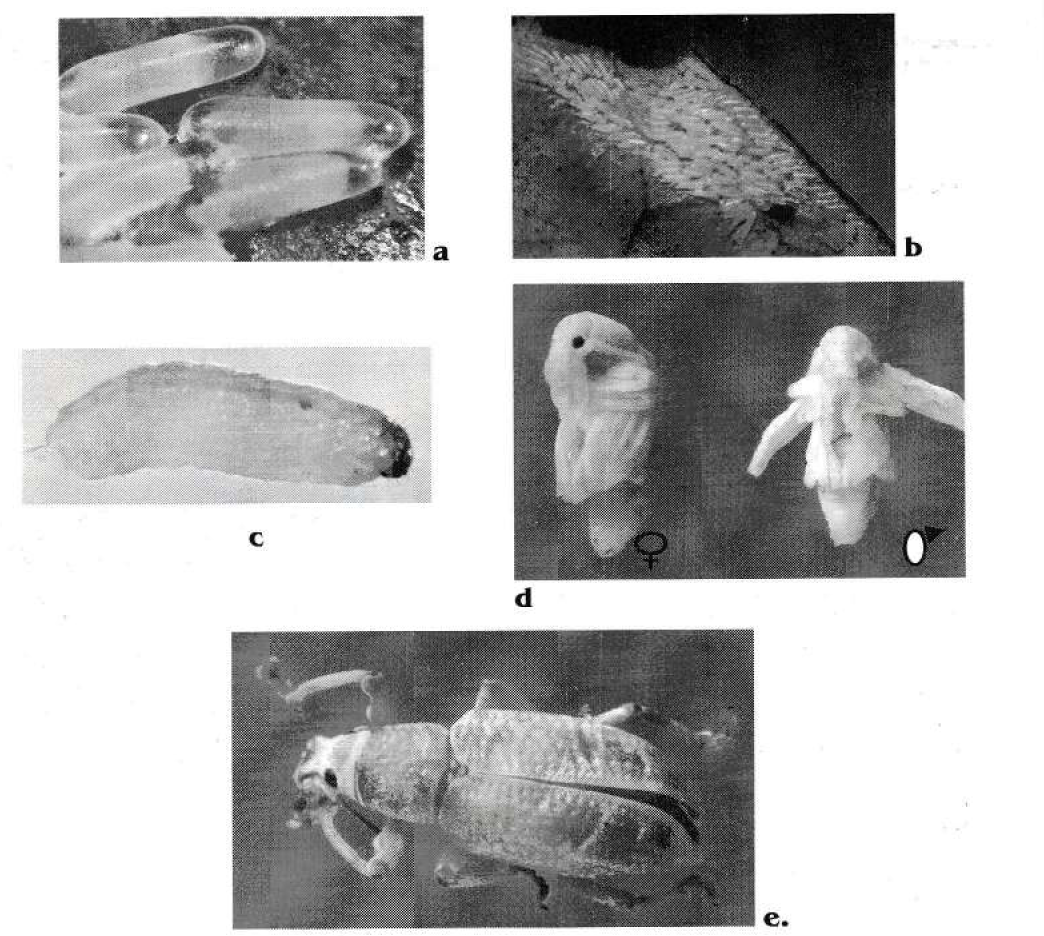
Estados de Compsus pos. n. sp. a. Huevos b. Postura c. Larva de primer ínstar d. Dimorfismo sexual en pupas e. Adulto sin emerger del suelo.
Las pupas son del tipo exarata, de color crema con ojos negros al final del desarrollo, al igual que los adultos se diferencian macho y hembra en el tamaño y en el último segmento abdominal las hembras miden 11±1.1 mm de longitud y presentan gonoporo en forma triangular, mientras que los machos miden 9.4±0.9 mm (Fig. 4d) y no posen este orificio, sólo apertura anal en forma transversal.
Periodo de preoviposición
En promedio la hembra de Compsus tiene un periodo de preoviposición de 21.42 ±13.82 días. Esta información se obtuvo de 21 hembras y fue muy variable (CV 76.1%), desde un día hasta un máximo de 49 días.
Duración del estado de huevo
La eclosión de los huevos se inició al noveno día en el laboratorio y en el décimo se presentó la emergencia de todas las larvas en observación. En muy bajo porcentaje de las posturas se observó deshidratación y no ocurrió eclosión. En el campo la duración del estado de huevo fue ligeramente más prolongada tomando dentro de las mangas entomológicas 11 a 13 y de 11 a 14 días cuando se colocaron en tiras dobles de plásticos y en hojas, respectivamente. El tiempo de la emergencia de las larvas en laboratorio y campo para cada postura fue muy uniforme, probablemente este aspecto es el más constante de una vida extremadamente variable como las de los picudos de los cítricos (Woodruff 1968).
En el campo no hubo oviposición en todas las ramas con mangas en que se colocaron los adultos. Las proporciones de sexos (♀:♂) en las que se observó posturas en hojas a las 24 horas fueron: 0:10, 0:20, 1:5, 2:3, 3:3, 5:5.
En las 15 mangas que tenían tiras de plástico, las hembras prefirieron el plástico en vez de las hojas, resultados similares se registran para el picudo de los cítricos de la Florida donde prefirieron ovipositar en papel encerado (Adair et al. 1998). Este hecho se debe seguir investigando para utilizarlo en el futuro como medio de monitoreo y/o como práctica física para reducir las poblaciones de los sitios de alta prevalencia, ya que la viabilidad del huevo en la tiras plásticas es menor, pues se presenta ataque de hongos.
Estadios larvales
Según Montejo y Montes (s.f.) éste presenta cinco estadios de desarrollo. Los huevos recién ovipositados se observan de color amarillo claro en toda su longitud y esto constituye el primer estadio (E1), el segundo (E2) comienza aproximadamente a las 24 horas, el huevo se torna de un color menos intenso (más opaco) y en uno de los extremos la membrana vitelina se empieza a separar del corion haciendo que se vea hialino; transcurridas 48 horas, el huevo es de color crema y los dos extremos son translúcidos (Fig. 4a). Cuando se observan cambios en el color y la desaparición paulatina de los extremos translúcidos es la transición del E2 al estadio tres (E3). Al séptimo u octavo día el color de los huevos es blanco lechoso y en algunos se puede distinguir un pequeño punto rojo. Uno o dos días antes de eclosionar se observan claramente las mandíbulas de las larvas y esta etapa de desarrollo corresponde al estadio 4 (E4). El último estadio se presenta cuando la larva está completamente formada y se mueve rompiendo el corion por el extremo donde estaba ubicada la cabeza, sale el citoplasma y posteriormente la larva.
Número de larvas de Compsus pos. n. sp. evaluadas para determinar el desarrollo
Número de huevos por postura encontrados en diferentes localidades y obtenidos en laboratorio
Huevos/postura.
Determinación de la duración de los estados subterráneos (larva y pupa)
Las larvas que se alimentaron en raíces de yuca se desarrollaron más rápido que las alimentadas en raíces del patrón citrumelo 4475 y estas últimas alcanzan su desarrollo más precozmente que las criadas en laboratorio en arena y zanahoria. La duración de la larva depende de las condiciones de alimentación (Quintela et al. 1998) y tipo de suelo (Rogers et al 2000). Probablemente el tipo de alimentación está directamente relacionada con el desarrollo y esto sea una constante en los picudos asociados a cítricos.
El tiempo en el que se cumplió el desarrollo del estado larval fue entre 91 y 136 días en raíces de yuca; en raíces de cítricos tuvo una duración de 109 a 143 días. En cada evaluación se evidenció la mortalidad alta de las larvas; es decir, fue тиy bajo el número de pupas obtenidas, alrededor del 0.1% de las larvas infestadas (a los 91días (d) el número de pupas (n) recuperadas fue de 2; 108d, n=1; 109d, n=4; 118d, n=2; 136d, n= 2; 138d, n =3; 140d, n=5; 143d, n=3).
En laboratorio las unidades experimentales se acabaron a los 113 días de evaluación y no se encontraron pupas. Quintela et al. (1998) afirman que cuando las larvas de Diaprepes abbreviatus se alimentan con dietas artificiales deficientes, desarrollan ínstares supernumerarios y muy pocas llegan hasta pupas.
El comportamiento de las variables de crecimiento: longitud, ancho y diámetro de la cápsula céfalica (
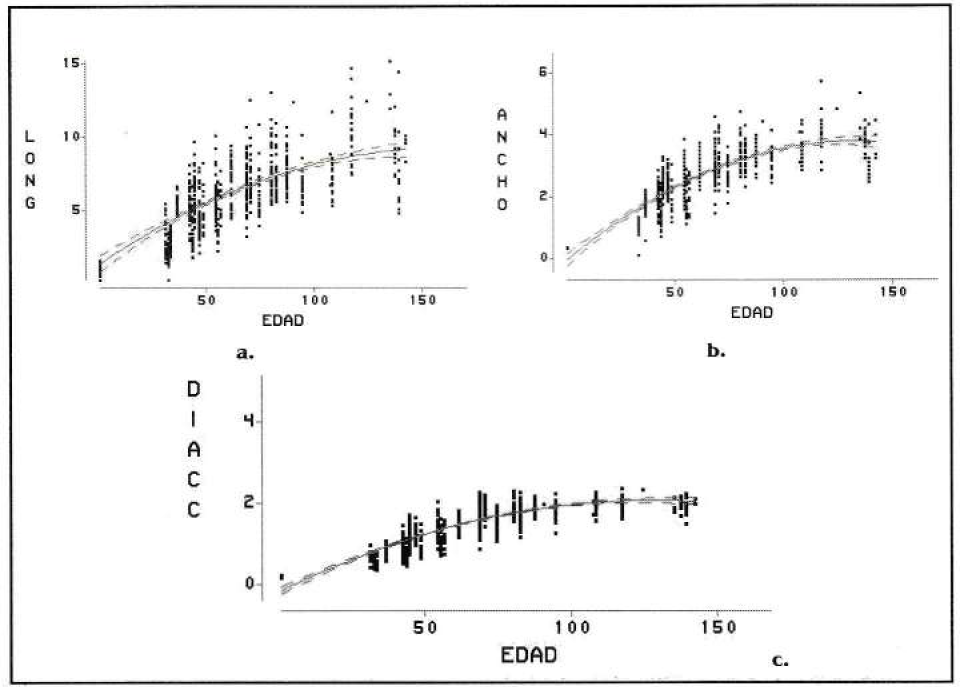
Comportamiento de las variables de crecimiento de las larvas en raíces de cítricos. a. Variable longitud vs edad b. Variable ancho vs edad c. Variable diámetro de cápsula céfalica vs edad.
Resultados de análisis de correlación de las variables de crecimiento en las larvas criadas en raíces de cítricos
Los adultos se recuperaron de las bolsas que contenían al patrón citrumelo 4475 a los 140, 143 y 171 días después de la infestación y 31, 34 y 62 días luego de la evidencia de la primera pupa indicando una duración de uno a dos meses (31 a 62 días) para este estado. La proporción entre machos y hembras encontrada fue de 0.9:1.
El tiempo de estado de huevo a adulto fue de 4.4 a 7.3 meses (Tabla 5). El tiempo generacional de Compsus pos. n. sp. fue de 134 a 268 días, un rango muy amplio en el que se pueden presentar de 1.36 a 2.72 generaciones de adultos al año; es decir, dos en promedio, este resultado se confirma en el estudio de comportamiento de la población en el que se registraron dos picos altos a lo largo del año. Se puede concluir que es una especie bivoltina.
Ciclo de vida del picudo de los cítricos en la zona central cafetera
Longevidad del adulto, capacidad y distribución de oviposición de las hembras
Se obtuvieron muy pocos adultos a lo largo del año en las trampas de emergencia instaladas de 0.3 m2. Whitwell (1990) utilizó cámaras de emergencia de 0.25 m2 en 20 semanas y colectó solo 5 picudos. La longevidad de los adultos y la capacidad de oviposición de las hembras fue muy alta, los adultos machos fueron más longevos que las hembras (Tabla 6). La capacidad de oviposición registró un máximo de 4264 huevos en 173 posturas durante un año (367 días, 12.23 meses); el promedio de número de huevos por día fue de 6.85±4.1, con un máximo de 18.46 y un mínimo de 2.69. La distribución de la oviposición, tuvo picos y descensos y ninguna relación directa con la edad de las hembras, no siguió ningún comportamiento que se pudiera explicar con funciones y no declinó al pasar el tiempo.
Longevidad de adultos y capacidad de oviposición del picudo de los cítricos en laboratorio
Comportamiento
En las evaluaciones de campo y del ciclo de vida se observó que las hembras de Compsus ovipositan en la parte aérea de los árboles (Fig. 6-1) pegando dos hojas o los pliegues de una con sus propias secreciones; dentro del pliegue dejan masas irregulares de una sola capa, muy pocas veces de dos o tres, el número de huevos por masa o postura en campo es muy variable (Tabla 3).
Las larvas neonatas caen a la superficie del suelo (Fig. 6-2), se entierran rápidamente e inician su vida subterránea (Fig. 6-3). En los primeros ínstares se alimentan de raícillas y pelos absorbentes (Fig. 6-4), posteriormente consumen la corteza de raíces secundarias y de la pivotante (Fig. 6-5,6). La larva en su último ínstar construye una cámara pupal de tierra y allí se alberga la pupa durante uno o dos meses (Fig. 6-7). El adulto emerge e inicia nuevamente su ciclo (Fig. 6-8).
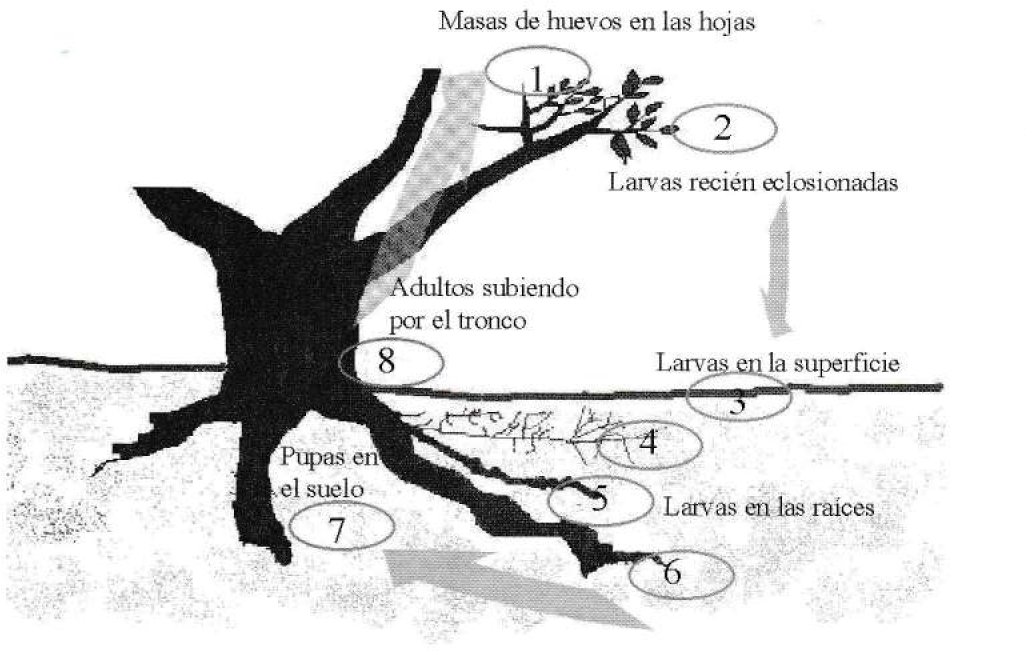
Ciclo de vida del picudo de los cítricos Compsus pos. n. sp.
La profundidad máxima a la que se encontraron estados subterráneos del picudo fue 94 cm en el Quindío y 60 cm en el Tolima; los estados más superficiales estaban a los 3 cm en las dos localidades. El mayor número de estados encontrados en los raíces de los árboles arrancados de siete años de edad fue de 54 para el Quindío y 35 para el Tolima.
La actividad alimenticia de los adultos ocurre tanto en el día como en la noche, la preferencia de hojas para alimentación y oviposición parecen ser las hojas del tercio medio de la rama; eventualmente se encontraron posturas y daños por alimentación en hojas muy viejas o muy nuevas. Los adultos se refugian en el envés de las hojas o en lugares sombreados; al ser disturbados o percibir movimientos toman como hábito de protección la tanotosis muy usada en curculionidos (Metcalf y Flint 1984).
El picudo de los cítricos es un mal volador, se ha observado que la mayoría de veces los adultos suben a los árboles caminando por el tronco, al sacudir los árboles algunos individuos abren los élitros antes de caer al suelo pero no alcanzan a volar; en la única ocasión que hay evidencia de vuelo es cuando se realizan aspersiones de insecticidas en los lotes. Este comportamiento genera indicios para la implementación de prácticas físicas de control (Cintas con pegante en el tronco).
Compsus pos n. sp. tiene una marcada preferencia por las hojas de naranja valencia, además se alimenta de una gran variedad de cítricos, como: swinglea, mandarina oneco, mandarina arrayana, lima ácida tahití, tangelo mineola, toronja, de una gran diversidad de plantas y de un amplio grupo de malezas (Tabla 7), lo cual permite ubicar a este picudo dentro del grupo de insectos polífagos. El daño de los adultos consiste en cortes irregulares en los márgenes de las hojas. Las larvas en la raíz consumen la epidermis en línea recta o en espiral siempre en forma ascendente.
Comportamiento de adultos de Compsus en el tiempo y su relación con las condiciones climáticas y la humedad del suelo
La población del picudo de los cítricos a través del tiempo fue muy variable (C.V. 68.5%). En general el número de adultos por árbol disminuyó a lo largo de las evaluaciones; al inició el número de adultos promedio por árbol fue de 15.71 ± 4.82 y para la última evaluación fue de 5.19 ± 3.11 (Fig. 7). Es importante aclarar que el lote donde se desarrolló la actividad no fue objeto de control contra esta plaga. Estos resultados hacen pensar que con la practica de recolección manual al 10% de los árboles de un lote y la regulación natural que ocurre se reduce el número de adultos de picudo por árbol.
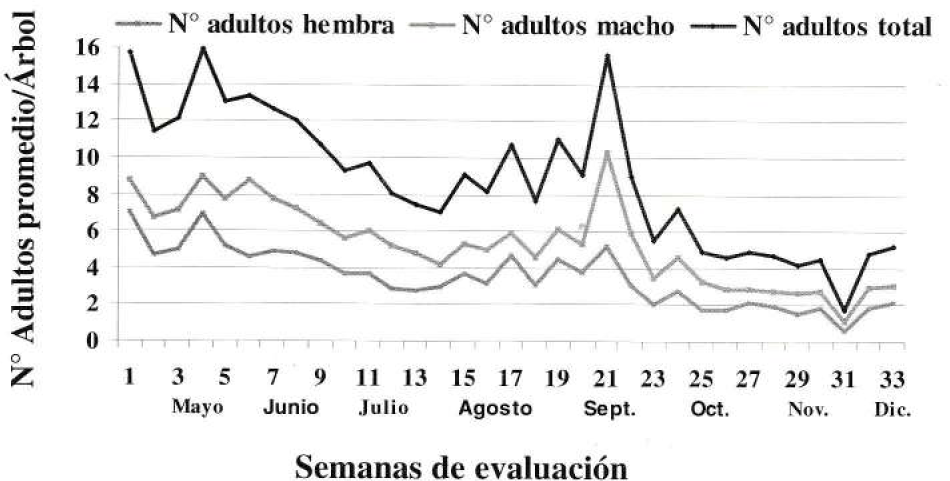
Comportamiento de la población de Compsus pos n. sp. en el tiempo.
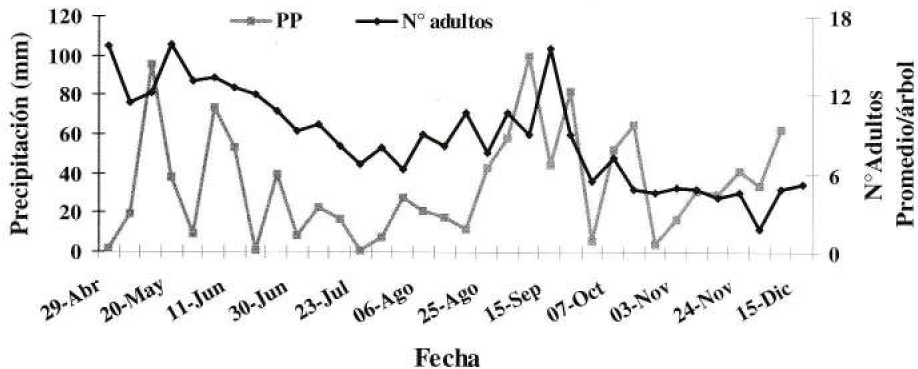
Relación de la precipitación con el comportamiento de la población de Compsus pos n. sp. en el tiempo.
También en la Figura 7, se observa que el número de hembras siempre fue menor con relación al número de machos. La proporción entre sexos encontrada en este ensayo fue de ♀:♂ 1:0.64, diferente a lo encontrado en los pocos adultos obtenidos de la cría en campo en bolsa de vivero e infestaciones artificiales.
En los meses de mayo y septiembre se observaron picos máximos de población que indican las generaciones de este insecto en el año. En la semana 31 de evaluación (principios de diciembre) se presentó el promedio mínimo de población 1.76 adultos/árbol (C.V 84.0%) posiblemente por que previo a la evaluación se realizó la recolección de naranja que causa la tanotosis ó caída de los adultos. Basados en esta observación se vislumbra el momento adecuado para colocar las cintas con pegante en el tronco de los árboles.
Ei número de adultos de Compsus por árbol presentó un intervalo muy amplio, el máximo fue de 32 que se obtuvo en la semana 21 de evaluación correspondiente a septiembre y el mínimo de cero presentándose en casi todas las evaluaciones, lo que hace pensar que es un insecto de hábitos gregarios o con distribución de contagio. Schroeder y Beavers (1985) en trabajos con el picudo de los cítricos, Diaprepes abbreviatus, afirman que posiblemente ellos se congregan por una feromona de agregación.
En análisis de correlación entre el número promedio de adultos por árbol semanalmente y los factores climáticos no mostró significancia (Tabla 8). La precipitación se relacionó con la población, coincidiendo las épocas de mayor precipitación con las de mayor población del picudo. En la Figura 8 se observa la influencia de las lluvias en el número de adultos, ya que la mayor población se presentó en mayo y septiembre después de semanas muy lluviosas. En los meses de junio y julio, donde se presenta el periodo seco para esta zona, ocurre una disminución paulatina en la población hasta llegar a una reducción del 50%. Después de septiembre la población disminuye en un 75% sin tener una correlación muy clara con la precipitación, probablemente por que no existía población del insecto en el suelo. Resultados similares han sido registrados por Whithwell (1990), aunque Méndez (1997) y Peña (1998) no han logrado establecer una correlación clara al analizar información relacionada con otros picudos que afectan también cítricos.
Plantas hospedantes de Compsus pos. n. sp.
Se encontraron dos larvas de Compsus n. sp. en el sistema radical de esta especie.
Análisis de correlación Población-Condiciones climáticas y el balance hídrico
T°, temperatura; H R, humedad relativa; PP, precipitación; Bs brillo solar; Exceso, déficit e índice de humedad en el suelo.
Según Méndez (1997), los picudos de los cítricos pueden reducir o ampliar su ciclo de vida dependiendo de condiciones climáticas; se presenta mayor emergencia de adultos en períodos con una humedad relativa alta, días largos y frecuentes y abundantes lluvias. En forma gráfica tampoco se pudo reiterar la influencia del brillo solar y la humedad relativa en la emergencia de adultos.
Por medio del balance hídrico de la zona se evidenció que en la mayor parte del año 1999 hubo exceso de agua en el suelo y que el índice de humedad fue casi siempre igual a uno o valores muy cercanos (0.95-1.1). Relacionando estos valores con los datos de la población, uno, dos y tres meses después se encontró una asociación positiva entre el número de adultos y el índice de humedad tres meses antes a la evaluación, pero con una variación explicada muy baja (r = 0.36); de forma gráfica no se encontró una relación consistente a lo largo del tiempo de ninguna de las variables.
Atracción de los adultos de Compsus a trampas de luz artificial y pitfall
En las trampas de luz instaladas sólo se recuperaron 2 individuos en un mes, la de luz blanca registró mayor número de adultos capturados pero sigue siendo un método inefectivo de detección de picudos. Beavers (1982) y Schoroeder (1984) evaluaron el uso de trampas de luz y determinaron que no eran eficaces para el picudo de los cítricos de la Florida. En las trampas de caída o "pitfall", se obtuvo solo captura de adultos en las cebadas con machos y parejas, Schoroder y Beavers (1985) demostraron la existencia de atrayentes semioquímicos, al parecer producidos por ambos sexos.
Para otras exploraciones es necesario mezclar el excremento con cloroformo, metanol y agua en una solución monofásica y no solo mirar en el recipiente trampa, sino en una área a la redonda (Whithwell 1990).
Enemigos nativos
En este estudio se reconoció la acción de los entomopatógenos B. bassiana aislados de adultos y larvas en Quindío y Tolima (Codificados como Bb 9903, Bb 9806, Bb 9807 en el cepario de Cenicafé) y Metarhizium anisopliae (Ma 9901) en larvas (Ma 0001, Ma 0002) y aislado de pupas y adultos sin emerger del suelo (Tabla 9).
En un adulto sin emerger del suelo, colectado en Tolima, se aislaron nemátodos entomopatógenos (M 653); los nemátodos emergidos (Ji3) se colocaron en prueba de patogenicidad con la polilla de los apiarios, Galleria melonella. La coloración obtenida en este hospedante fue marrón oscuro hasta vino tinto lo que coincide con la sintomatología mostrada por nemátodos de la familia Heterorhabditidae.
El uso de nemátodos entomopatógenos para la disminución de las poblaciones en el suelo es promisorio. En ensayos hechos por Schroeder (1990) hubo reducción de emergencia de adultos hasta en un 70%, en estudios en Cuba con H. heliothidis en vivero y en campo se observaron mortalidades entre un 63% al 100% de las larvas de P. litus, con el nemátodo Steirnenema riobravis se redujeron poblaciones de larvas en un 80-90% (Montejo y Montes sf).
En el estado de huevo se recuperaron ocho parasitoides identificados por el Dr. John LaSalle (Unit of Parasitoid Systematics, CABI Bioscience Centre, Reino Unido), Dra. Rita Duncan (Universidad de la Florida) y el Dr. John Pinto (Universidad de California) como Haeckeliana sp. (Hymenoptera: Trichogrammatidae) (cuatro especies), Hadrotrichodes sp. (dos especies), Oomyzus sp. (Hymenoptera: Eulophidae) y el hiperparásito Horismenus sp. (Hymenoptera: Eulophidae) (Tabla 9). También se encontraron en hojas evidencia de posturas o posturas destruidas por ácaros de la familia Phytoseiidae.
Como depredadores se han colectado diferentes especies de hormigas llevando en sus mandíbulas larvas, pupas y adultos. Las hormigas se identificaron hasta género en el Museo Entomológico "Francisco Luis Gallego" (Tabla 9). También se encontraron, con frecuencia, adultos inmovilizados por telarañas o destruidos por arañas.
Nemátodos del género Hexamermis, (Mermithidae) se visualizaron emergiendo de larvas del picudo en el suelo, así como especies de insectos depredadores de las familias Carabidae, Reduviidae, Cicindellidae, Hidrophilidae, Belostomatidae, Gelastocoridae, Staphilinidae y Dermaptera.
Los hallazgos anteriores indican que el picudo de los cítricos posee una buena diversidad de enemigos que pueden regular sus poblaciones cuando el agroecosistema está en equilibrio. Además, las experiencias en otros países como Cuba muestran que a pesar de luchar contra los picudos con grandes cantidades de productos químicos no se obtuvieron resultados satisfactorios, ya que, el daño se mantiene estable en áreas tratadas y sin tratamiento (Montejo y Montes sf).
Lista de enemigos naturales del picudo de los cítricos Compsus pos. n. sp.
Q: Quindío y T: Tolima.