
Abstract
During 1997 a study about natural parasitism on the different biological stages of the fruit borer of solanaceae Neoleucinodes elegantalis Guenée (Lep: Pyralidae) was carried out. Eggs, worms and pupae in crops of Solanum quitoense Lamark (lulo), Ciphomandra betacea Sent (tomate de árbol) and Lycopersicum esculentum Mill (tomate de mesa), were collected in the departments of Cauca and Valle del Cauca. The results suggest that eggs, larvae and pupae of the fruit borer insect have a great variety of natural control. Trichogramma exiguum and T. pretiosum (Hymenoptera: Trichogrammatidae) were detected parasiting eggs (4.2 and 54%). The species Copidosoma sp. (Hymenoptera: Encyrtidae) was detected parasiting worms with 16.6 to 60%. Lixophaga sp. (Diptera: Tachinidae) was also observed parasiting worms in the last instar with 0.8 to 22% of parasitism. Different parasites such as Brachymeria sp., Conura sp. (Hymenoptera: Chalcididae), Aprostocetus sp. (Hymenoptera: Eulophidae) and 3 Hymenopteros of Ichneumonidae emerged from N. elegantalis pupae. Parasitism on pupae reached 18%, 4.7%, 25% and 20%, respectively.
Introducción
El pasador del fruto Neoleucinodes elegantalis Guenée es considerada una de las plagas limitantes de los cultivos solanáceos como lulo (Solanum quitoense Lamark), tomate de árbol (Cyphomandra betacea Sendt) y tomate de mesa (Lycopersicum esculentum Mill). Las pérdidas ocasionadas por esta plaga, en frutos de estas solanáceas, pueden ser superiores al 70% (ICA, 1981a; 1981b; 1981c; 1982; 1988a; 1993).
Una de las medidas culturales recomendadas para reducir el daño causado por la plaga y su dispersión en los cultivos infestados ha sido la recolección y entierro de frutos que presenten daño. Costa Lima (1949) recomienda el uso de soluciones arsénicales, asperjando los frutos pequeños, como método preventivo.
Cross (1996) encontró que las liberaciones de Trichogramma pretiosum Riley, en dosis de 162 pulgadas/ha/semana, cada cuatro días, reducen en un 22% el daño ocasionado por N. elegantalis en tomate de mesa, causando parasitismo en huevo hasta de 84%.
Según estudios realizados por Silveira (1976), citado por Varela (1997), muestran un 76.6 % de eficiencia en la captura de adultos de N. elegantalis utilizando trampas de luz del tipo "Luiz de Queiroz". Souza (1985) recomienda plantar tagetes (Tagetes sp.) junto a los cultivos para evitar la llegada del pasador. También aconseja realizar aspersiones de polvo de derris y azufre, mezclados en proporción 1:1.
En Brasil se está investigando otros métodos de manejo entre ellos el uso de insecticidas fisiológicos y feromonas. Los insecticidas fisiológicos como la abamectina, clorfluazuron y triflumuron para el control del pasador Neoleucinodes elegantalis son sustancias promisorias que muestran más del 80% de eficiencia al ser aplicadas contra la plaga, reduciendo tanto el número de frutos atacados por el insecto como el número de larvas por fruto (Souza y Reis 1996).
De acuerdo con Fernández et al. (1982) la feromona sexual utilizando hembras de la plaga como atrayentes para la captura de Neoleucinodes fue más eficiente que la atracción ejercida por la luz producida por mecheros empleando gasoil como combustible.
En Colombia, específicamente en el Valle del Cauca, el pasador es controlado por los agricultores con aplicaciones de insecticidas, los cuales resultan ineficientes debido al hábito de vida de la plaga. La larva del pasador cuando nace se introduce rápidamente dentro del fruto. completa su desarrollo dentro de éste y sólo sale cuando está listo para empupar. De esta forma el fruto protege a la larva contra la acción de las sustancias químicas.
El control biológico se presenta como una alternativa dentro de la estructuración de programas de manejo integrado de esta plaga; con el fin de conocer y utilizar el potencial biológico que podría actuar como regulador de Neoleucinodes elegantalis a nivel de campo se realizó este estudio con los siguientes objetivos: Reconocer e identificar los agentes naturales de control biológico natural del pasador del fruto Neoleucinodes elegantalis en plantas hospedantes de la plaga como lulo, tomate de mesa y tomate de árbol, cultivados en algunos municipios del Cauca y Valle del Cauca. Determinar los niveles de parasitismo natural sobre Neoleucinodes elegantalis después de muestrear huevos, larvas y pupas en cada uno de los cultivos de solanáceas hospedantes.
Materiales y Métodos
Los estudios sobre reconocimiento e identificación de agentes de control biológico del pasador del fruto de solanáceas Neoleucinodes elegantalis, fueron realizados bajo condiciones de campo y laboratorio.
Durante 1997 se visitaron cultivos solanáceos de diferentes municipios del Cauca y Valle del Cauca realizando muestreos de huevos, larvas y pupas de N. elegan-talis. Se visitaron 22 municipios en el departamento del Valle del Cauca y 3 en el departamento del Cauca. En estos municipios se visitaron 78 fincas; 29 de ellas productoras de tomate de mesa, 12 de tomate de árbol y 37 de lulo. El número de fincas visitadas por municipio fluctuó entre 1 y 6 explotaciones (Tabla 1).
Fincas productoras de lulo, tomate de arbol y tomate de mesa, visitadas por municipio para el estudio de la fauna benéfica natural de Neoleucinodes elegantalis. 1997-1998.
Los cultivos de lulo y tomate de árbol se encuentran ubicados en zona de ladera entre los 1500 y 2000 msnm (zona marginal cafetera) con temperatura promedio de 17.5 °C y los cultivos de tomate se localizan entre los 1000 y 1200 msnm, con temperatura promedio de 24 °C. En estos cultivos, los agricultores hacían control químico y control mecánico, recogiendo y enterrando los frutos infestados.
El muestreo de los diferentes estados biológicos de Neoleucinodes se realizó teniendo en cuenta los hábitos de oviposición, daño y empupamiento. Para el caso de los huevos, se examinaron inflorescencias y frutos pequeños de las plantas hospedantes. Estos se retiraron. colocaron en cápsulas de gelatina y se llevaron al laboratorio para determinar su parasitismo. Las larvas se obtuvieron de la recolección de frutos infestados con perforaciones de entrada (espinillas) y orificios de salida. El número de frutos recolectados dependió del nivel de infestación de cada finca, pero se buscó que en cada visita se tomara un número superior a 40 frutos. El muestreo de prepupas y pupas de N. elegantalis se realizó sobre las hojas secas de la planta hospedante. Se inspeccionó también la hojarasca del suelo. El material se guardó en frascos de boca ancha y se llevó al laboratorio.
Los estudios en laboratorio se realizaron en el C.I. Palmira Programa MIP, Corpoi-ca, situado a 1000 msnm, temperatura promedio de 25 Cº y humedad relativa del 70%.
Con la ayuda de un estereoscopio se determinó el parasitismo por Trichogramma en huevos de Neoleucinodes traídos del campo después de diez días de su recolección. Para determinar el porcentaje de parasitismo por Trichogramma, se relacionaron los huevos parasitados, cuya coloración es negra, con el total de huevos muestrados en cada fecha. Las larvas recién nacidas fueron retiradas de la cápsula y con ayuda de un pincel se colocaron en un vaso con dieta (propuesta por Muñoz y Serrano (1989)) para lograr su desarrollo y determinar su parasitismo. Los frutos recolectados en el campo se abrieron, realizando 2 cortes longitudinales, con el fin de extraer las larvas, las cuales fueron colocadas individualmente en vasos plásticos con dieta artificial. Las pupas procedentes de campo se colocaron en frascos de vidrio de boca ancha con bagacillo humedecido y previamente tratado con hipoclorito al 0.5 %. Se hicieron observaciones periódicas para detectar la presencia de parasitoides en larvas y pupas; se anotaron los signos y síntomas de parasitismo observado. Una vez emergidos los adultos de los parasitoides se contabilizaron y se introdujeron en viales con alcohol al 70% o se montaron en alfileres entomológicos para su identificación.
Se realizó un muestreo por finca, llevando un registro detallado del número de huevos, larvas y pupas recolectadas. Las larvas y pupas colectadas se manejaron individualmente hasta finalizar el desarrollo biológico del insecto, determinando de esta forma la presencia o ausencia de parasitoides.
La identificación de los agentes benéficos se hizo a través de comparación con especímenes de la Colección Económica de plagas y benéficos del C.I. Palmira, CORPOICA y mediante el envió al Departamento de Agricultura de los Estados Unidos, USDA (Systematic Entomology Laboratory). Para el caso de Trichogramma se procedió a identificar las especies a través de la técnica de extracción de genitalia en machos; técnica propuesta por Nagarkatti y Nagaraja (1971).
Resultados y Discusión
Los huevos del pasador son colocados en diferentes sitios del fruto y sobre las inflorescencias de las plantas hospedantes, en forma aislada o en grupos fuertemente apilados. En lulo, las posturas se encontraron sobre la parte media de la superficie del fruto, específicamente en la base de los tricomas. Muñoz y Serrano (1989) encontraron que el sitio de mayor oviposición del pasador sobre frutos de esta solanácea ocurre en la parte basal del fruto cerca al cáliz. Según los mismos autores, los huevos se localizan en los tricomas en su parte basal, media y apical, siendo este último lugar el más frecuente para la oviposición. En frutos de tomate de árbol las posturas del pasador son expuestas, ya que el fruto no tiene tricomas ni sépalos largos que protejan los huevos. Las posturas se localizaron entre la unión del fruto con el pedúnculo. Al parecer la hembra del pasador inyecta los huevos en estos sitios. También se hallaron posturas indistintamente sobre la superficie del fruto. Las posturas sobre frutos de tomate de mesa se encontraron individuales y en grupos, preferiblemente, debajo de los sépalos y muy pocas veces sobre la superficie del fruto. Aunque es común encontrarlas en hojas y tallos cuando las poblaciones de la plaga son altas. Los resultados coinciden con las anotaciones realizadas por Fernández et al. (1982), Muñoz y Serrano (1989) y García (1991) quienes señalan que el sitio preferido por el adulto para realizar su oviposición es el espacio comprendido entre el cáliz y el fruto. Sin embargo, Marcano (1991) observó que las hembras de N. elegantalis ovipositaron sobre el fruto y eventualmente debajo de los sépalos.
El lugar de empupamiento del pasador estuvo relacionado con el tipo de planta hospedante. En lulo el pasador empupa en botones florales y en las hojas secas que se encuentran en las axilas de las plantas. Este lugar de empupamiento fue descrito por Muñoz y Serrano (1989). En tomate de árbol se localiza en las hojas secas que se encuentran sobre la superficie del suelo; la larva corta una porción de la hoja, la dobla y la pega con hilos de seda formando un capullo. En tomate de mesa, forma un capullo con las hojas verdes o secas de la planta (cercanas a los frutos con orificios de salida). Lo contrario fue observado por Salas et al. (1991) quienes anotan que el pasador empupa en el suelo.
Parasitoides de huevos
Se evaluaron 1506 huevos del pasador de los cuales sólo el 1.3% (21) se recolectaron en lulo, el 1.3% (22) en tomate de árbol y el 97.4% (1463) en tomate de mesa, lo cual indica la presencia de altas poblaciones de la plaga en esta solanácea.
Como parasitoides de huevos de Neoleucinodes elegantalis se presentaron dos avispas de la familia Trichogrammatidae (Fig. 1). Estos fueron reconocidos por la Dra. Ana E. Díaz 4 como: Trichogramma exiguum Pinto y Platner y Trichogramma pretiosum Riley. Los parasitoides emergieron de huevos encontrados sobre frutos de tomate de mesa y tomate de árbol. Los huevos de N. elegantalis parasitados por Trichogramma se caracterizan porque el corion toma una coloración negra. Se observó además que cada huevo parasitado de Neoleucinodes recolectado en campo puede albergar entre 1 y 2 adultos de Trichogramma. Muñoz y Serrano (1989) encontraron que, bajo condiciones de laboratorio, emergen hasta 3 adultos de Trichogramma.
Programa MIP CORPOICA. Pnlmira.
J.A. CORPOICA. Programa MIP.

Trichogramma sp. (Hymenoptera: Trichogrammatidae) parasitando un huevo del pasador Neoleucinodes elegantalis. (Foto: Hery F. Viáfara M.)
Hassan (1991), citado por Berti y Marcano (1995), afirma que uno de los criterios para la selección de especies de Trichogramma, a ser usadas en las liberaciones masivas, es aquel concerniente a la preferencia que presenta el benéfico por sus huéspedes naturales. Por esta razón, la presencia de las dos especies de Trichogrammatidae sobre posturas de Neoleucinodes deja una camino abierto para posteriores investigaciones que permitan conocer cual de las dos especies presenta mayor preferencia por las posturas de la plaga.
En forma natural se hallaron ambas especies de Trichogramma en socas o cultivos abandonados de tomate de mesa. T. exiguum fue encontrado en el municipio de la Unión y el parasitismo fue del 4.2%. En el municipio de Palmira se detectó a Trichogramma pretiosum con un parasitismo del 4.7% (Tabla 2). Muñoz y Serrano (1989) registraron la presencia de Trichogramma cerca pretiosum, identificado por Pinto y Platner, taxónomos de la Universidad de California, parasitando huevos de Neoleucinodes. Estos autores encontraron un parasitismo de 81.93% al examinar huevos de la plaga recolectados en un cultivo abandonado de tomate ubicado en el corregimiento del Bolo Hartonal, jurisdicción del municipio de Pradera.
La presencia de poblaciones naturales de estos parasitoides en socas o cultivos abandonados de tomate muestran la existencia de un acervo natural que contribuye a reducir las poblaciones de la plaga en el ambiente. Esta reserva se conserva gracias a las constantes liberaciones masivas en el Valle del Cauca, no sólo en el cultivo del tomate de mesa sino en otros cultivos como algodón, soya, yuca, caña de azúcar.
En cultivos de tomate de mesa ubicados en el municipio de Palmira, se determinó el parasitismo por T. exiguum producto de las liberaciones de este benéfico. El parasitismo fue del 12.3% y 15.8%. Igualmente, en un cultivo de tomate de árbol ubicado en el municipio de Guacarí, se registró un parasitismo del 12% por esta misma especie (Tabla 2).
T. pretiosum parasitó el 10.3 y el 11.2% de huevos del pasador en cultivos comerciales de tomate de mesa de los municipios de Cerrito y Yotoco, respectivamente. Algunos productores de esta hortaliza utilizan las dos especies de Trichogramma para el manejo de la poblaciones de la plaga. En el municipio de Rio Frío y Palmira se presentaron parasitismos del 54 y 30.7% usando las dos especies, en cultivos de tomate (Tabla 2).
Las dosis de Trichogramma utilizadas por los agricultores para el control del pasador fluctuaron entre 300 a 715 pulgadas/ha semana, fraccionadas en liberaciones cada 8 y 4 días, incluso en liberaciones diarias del benéfico, donde se alcanzó el parasitismo más alto.
Los porcentajes bajos de parasitismo obtenidos en los muestreos pudieron estar afectados por numerosos factores entre ellos las frecuentes aplicaciones de insecticidas, la oportunidad de la liberación, la distribución del parasitoide, el momento de la evaluación; el método, dosis y frecuencia de liberación del benéfico, etapa del cultivo, tamaño de la muestra, calidad del insumo, etc. Por estas razones, resulta difícil hacer una interpretación acertada que permita calificar adecuadamente la efectividad de este parasitoide. No obstante, pese a los niveles bajos de parasitismo, Trichogramma puede considerarse como uno de los componentes claves dentro de los programas de manejo integrado de Neoleucinodes en cultivos de tomate de mesa y tomate de árbol. Este parasitoide presenta la ventaja de eliminar la plaga en estado de huevo impidiendo el nacimiento de larvas que hacen daño a los frutos. En cultivos de lulo su utilización es casi imposible, ya que los tricomas o vellosidades de los frutos dificultan la acción del benéfico.
Parasitismo por Trichogramma pretiosum y T. exiguum sobre huevos de Neoleucinodes elegantalis en cultivos de tomate de mesa y tomate de árbol en algunos municipios del Valle del Cauca. 1997 - 1998.
Parasitismo natural
Parasitismo inducido
Parasitismo sobre larvas de N. elegantalis
Se recolectaron 1.229 frutos de lulo, 345 de tomate de árbol y 263 tomate de mesa; de los cuales se extrajeron 1.488, 134 y 537 larvas del pasador respectivamente.
Copidosoma sp. (Hymenoptera: Encyrtidae)
Copidosoma sp., identificado por Norman E. Woodley (Research Entomologist, SEL., USDA 1997), se registra emergiendo de larvas de Neoleucinodes recolectadas en campo. Fue señalado por Muñoz y Serrano en el año de 1989 en larvas del pasador procedentes de frutos de lulo del municipio de Pradera. Este Encyrtidae es una especie nativa que parasita los huevos pero los síntomas y signos de parasitismo se manifiestan en estado de larva, comúnmente conocida como "momia" (Fig. 2). Copidosoma presenta un alto grado de especificidad para parasitar los huevos del pasador. En ensayos realizados en CORPOICA (1997) se determinó que este parasitoide prefiere únicamente huevos de Neoleucinodes cuando, simultáneamente, se ofrecieron huevos de Trichoplusia ni, Phthorimaea operculella, Galleria mellonella y Neoleucinodes elegantalis.
El tiempo que transcurre desde la presencia del primer síntoma hasta la emergencia del parasitoide fue de aproximadamente 23 días, a 25 C° y 70% de humedad relativa. Se vio que de una larva de Neo-leucinodes, parasitada por Copidosoma, pueden emerger entre 100 y 200 adultos del benéfico. Muñoz y Serrano (1989) encontraron que el número de adultos del parasitoide emergidos por larva fue de 74 a 120.

Adultos de Copidosoma sp (Hymenoptera: Encyrtidae) parasitoide huevo larva de Neoleucinodes elegantalis. Obsérvese la larva del pasador parasitada o "momia" albergando las pupas del benéfico. (Foto: Ricardo Alzate).
Los porcentajes naturales de parasitismo por Copidosoma sp. se presentaron principalmente en cultivos de tomate de árbol ubicados a alturas entre 1600 y 1800 msnm; en los municipios de Toro, Trujillo, La Cumbre, Buga y Guacari. Los niveles de pararasitismo fluctuaron entre un 14.5 y un 60% (Tabla 3). Muñoz y Serrano (1989) registraron un porcentaje de parasitismo natural del 1.4% al examinar larvas del pasador extraídas de frutos de lulo.
Lixophaga sp. (Diptera: Tachinidae)
Lixophaga sp. identificado por Norman E. Woodley (Research Entomologist, SEL., USDA. 1997) (Fig. 3), es un parasitoide nativo de larvas de Neoleucinodes de último instar. Al parecer, las hembras del Tachinidae depositan sus maggots o larvas en los orificios de salida de los frutos atacados; estos penetran en las galerías y buscan a las larvas del pasador que se encuentren todavía dentro del fruto para parasitarlos.
La larva de N. elegantalis parasitada por Lixophaga sp. no presenta características particulares que permitan discriminar entre una larva sana y una parasitada. Sin embargo, cuando la larva del parasitoide se encuentra bien desarrollada, ésta se puede observar a través de la piel de la larva hospedera. Bajo condiciones de laboratorio se observó que la larva del díptero consume todo el interior de la larva de Neoleucinodes y posteriormente sale a empupar. El tiempo de duración de Lixophaga sp. en estado de pupa fue de 18 días aproximadamente (20 observaciones) a 25 C y 70% humedad relativa. De una larva Neoleucinodes parasitada por Lixophaga sp. pueden emerger entre 1 y 3 individuos del benéfico.
Este parasitoide se muestra como una alternativa viable para el manejo del pasador en cultivos de lulo, ya que fue uno de los parasitoides más abundantes encontrados en larvas provenientes de frutos de esta solanácea. Su cría masiva podría ser importante en la estructuración de programas MIP para el manejo de esta plaga en cultivos de lulo.
Los porcentajes de parasitismo natural por Lixophaga sp. fluctuaron entre 0.8 y 22% (Tabla 3). Este parasitoide emergió de larvas extraídas en su totalidad de frutos de lulo de cultivos ubicados a alturas entre 1500 y 1750 msnm en municipios de Argelia, Bolivar, Tulúa y Buga. Muñoz y Serrano (1989) observaron un parasitismo de 0.08% por Lixophaga sp. al examinar larvas de Neoleucinodes en cultivos de esta solanácea del municipio de pradera (V). En cultivos de lulo, ubicados en Anolaima (Cundinamarca), se registró entre 11 y 47% de parasitismo sobre larvas del pasador parasitadas por este díptero (ICA 1988b).
Parasitismo sobre larvas de Neoleucinodes elegantalis en cultivos de lulo y tomate de árbol en algunas localidades del Valle del Cauca. 1997-1998

Lixophaga sp. (Diptera: Tachinidae) parasitoide de larvas de último instar del pasador del fruto Neoleucinodes elegantalis. (Foto: Hery F. Viáfara M.).
Observaciones realizadas por Trochez (1997) 5 indican que el parasitismo por Copidosoma es anulado por la presencia de Lixophaga sp., cuando se presentan casos de parasitismo múltiple, especialmente en aquellas larvas de Neoleucinodes provenientes de frutos de tomate de árbol.
Parasitoides de pupas de N. elegantalis
Se recolectaron pupas de 10 fincas en 8 municipios del Cauca y Valle del Cauca. De estas fincas, 3 eran productoras de lulo, 2 de tomate de árbol y 3 de tomate de mesa. Se recolectaron un total de 133 pupas del pasador. Como parasitoides de pupas se encontraron seis Hymenopteros, dos de la familia Chalcididae, uno de la familia Eulophidae y tres especies de Ichneumonidae, sin identificar.
Brachymeria sp. (Hymenoptera: Chalcididae)
Este Chalcididae fue determinado por Eric Grissell (Research Entomologist, SEL. USDA. 1997) como Brachymeria sp. (Fig. 4).
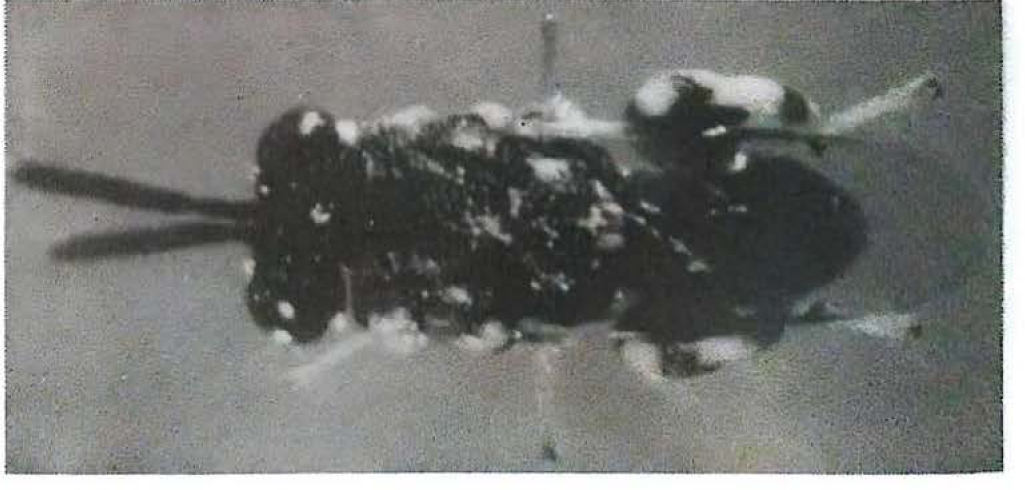
Brachymeria sp. (Hymenoptera: Chalcididae) parasitoide de pupas de Neoleucinodes elegantalis. (Foto: Hery F. Viáfara M.)
Bajo condiciones de laboratorio se hicieron observaciones sobre la biología de este parasitoide y se determinó que una pupa parasitada alberga a un sólo individuo del benéfico y que su periodo de desarrollo en pupas del pasador fue al rededor de 12 días; los adultos presentan alta longevidad (±40 días) su tamaño y su capacidad de vuelo facilitan su dispersión.
Las pupas de Neoleucinodes parasitadas por Brachymeria se caracterizan por presentar puntos negros que al parecer son las señales de oviposición dejadas por las hembras del parasitoide. Con el tiempo estas pupas pierden movilidad y se presentan cambios en la coloración pasando de un marrón claro a uno oscuro, casi negro. En días próximos a la emergencia del parasitoide, este se puede detallar cuando las pupas son observadas a trasluz.
Al exponer pupas de Trichoplusia ni, Galleria mellonella y N. elegantalis a hembras de Brachymeria se obtuvieron porcentajes de parasitismo positivo del 60%, 65% y 65%, respectivamente. Se observó un 30% de superparasitismo en las pupas expuestas, por lo que se recomienda realizar estudios para determinar el número de parejas de Brachymeria y el tiempo de exposición más adecuado para propagarlos mas eficientemente (CORPOICA 1997).
Los porcentajes de parasitismo de estos dos Chalcididae se muestran en la Tabla 4. El parasitismo por Brachymeria sobre pupas recolectadas en tomate de mesa fue del 18 y 23%; este último porcentaje se presentó en una soca del cultivo ubicada en el municipio de Palmira.
Conura sp. (Hymenoptera: Chalcididae)
Este Chalcididae fue determinado por Eric Grissell (Research Entomologist, SEL. USDA. 1997) Al igual que Brachymeria, este parasitoide fue encontrado en pupas de Neoleucinodes en cultivos de tomate de mesa. Los Chalcididae del género Conura (Fig. 5), y en general todos los Chalcididae, son considerados como parasitoides o hiperparásitos. En Colombia se describieron recientemente cuatro especies nuevas dentro del género Conura (Spilochalcis). Las especies fueron colectadas principalmente en cultivos de palma africana de la región de San Alberto (Rio de Oro, Cesar) (ICA 1989). Una pupa del pasador alberga un solo individuo de Conura. Las características del parasitismo de Conura sp. en pupas de Neoleucinodes son muy similares a las presentadas por la especies Brachymeria sp. El parasitismo por Conura sp. fue de 4.7% en pupas recolectadas en una soca de tomate de mesa, ubicada también en el municipio de Palmira (Tabla 4).
Parasitismos sobre pupas de Neoleucidones elegantalis en cultivos de tomate de mesa, tomate de árbol y lulo en algunos municipios del Cauca y Valle del Cauca. 1997 - 1998
Aprostocetus sp. (Hymenoptera: Eulophidae)
Al igual que los miembros de la familia Trichogrammatidae y Encyrtidae, los individuos de la familia Eulophidae son muy importantes por ser, a menudo, utilizados en la lucha biológica contra plagas (Coronado y Marquéz 1972; Delvare 1988).
Como parasitoide de pupas de Neoleucinodes, en cultivos de lulo y tomate de árbol, se registra un Eulophido, determinado por Michael E. Schauff (Research Entomologist, SEL. USDA. 1997) como Aprostocetus sp. Una pupa del pasador parasitada por Aprostocetus sp. alberga entre 30 y 40 adultos del benéfico (Fig. 6).
El parasitismo de Aprostocetus sp. fue del 11.1% en pupas recolectadas tomate de árbol en el municipio de Guacarí. Igualmente, se encontró un 4% de parasitismo por este Eulophidae en pupas provenientes de un cultivo de lulo ubicado en el municipio de Popayán (Tabla 4).
Parasitoides de la familia Ichneumonidae
Se presentaron tres especies de Ichneumonidae parasitando pupas del pasador en cultivos de tomate de árbol, tomate de mesa y lulo. De una pupa de Neoleucinodes parasitada por cualquiera de los Ichneumonidae encontrados (Figs. 7–9) emergen un solo adulto del parasitoide. Una de las especies de Ichneumonidae mostró un amplio margen de distribución, al registrarse en pupas del pasador obtenidas de cultivos de tomate de mesa a 1.000 msnm y de cultivos de lulo y tomate de árbol a 1.800 msnm. El parasitismo por esta especie de Ichneumonidae fue de 12%, 44.4% y 4.7%. Estos parasitismos se encontraron en cultivos ubicados en los municipios de Guacarí, Palmira y Popayán (Tabla 4).
Los porcentajes de parasitismo por las otras dos especies de Ichneumonidos fueron del 4% en cultivos de lulo ubicados en el municipio de Popayán (Tabla 4).
Conclusiones
Se reconocieron 10 especies de parasitoides que atacan los diferentes estados biológicos de Neoleucinodes elegantalis, algunos de ellos ya registrados en el Valle del Cauca y en otros de departamentos de Colombia: Trichogramma exiguum y T. pretiosum, parasitoides de huevos; Copidosoma sp., parasitoide huevo larva; Lixophaga sp., parasitoide de larvas de último ínstar; Brachymeria sp., Conura sp., Aprostocetus sp. y tres especies de la familia Ichneumonidae como parasitoides de pupas.
En zonas de ladera se presentó una mayor abundancia de entomofauna benéfica. Entre los parasitoides que revisten importancia en esta zona se encuentra Copidosoma sp., el cual registró porcentajes de parasitismos hasta del 60% en cultivos de tomate de árbol, por lo cual se constituye un parasitoide promisorio para el control de la plaga en esta solanácea.
Aunque los porcentajes de parasitismo natural presentados por Lixophaga sp. sobre larvas de Neoleucinodes elegantalis no fueron superiores al 22%, este benéfico podría ser una alternativa viable para el manejo del pasador en cultivos de lulo dada su abundancia en este cultivo.
Las numerosas y frecuentes aplicaciones de insecticidas en zona plana, especialmente en cultivos de tomate de mesa, hicieron que las poblaciones de enemigos naturales del pasador fueran escasas.
Trichogramma exiguum y T. pretiosum pueden considerarse un componente clave dentro de los programas de manejo integrado de Neoleucinodes en cultivos de tomate de mesa y tomate de árbol. En cultivos de lulo su utilización podría resultar ineficiente dada la cantidad de tricomas que presentan los frutos.
Las características biológicas de Brachymeria sp. tales como longevidad, tamaño y la facilidad que presenta su cría masiva, hacen que este parasitoide pueda ser considerado como otro componente dentro de la estructuración de programas MIP para el manejo del pasador en los diferentes cultivos de solanáceas.
La presencia y acción de estos benéficos es muy importante y deja un camino abierto a su posible utilización práctica, ya sea desarrollando técnicas de cría masiva o transportando benéficos de zonas de alta concentración a zonas en donde la presencia de la entomofauna benéfica sea baja.

Conura sp. (Hymenoptera: Chalcididae) parasitoide de pupas de Neoleucinodes. elegantalis (Foto: Hery F. Viáfara M.)
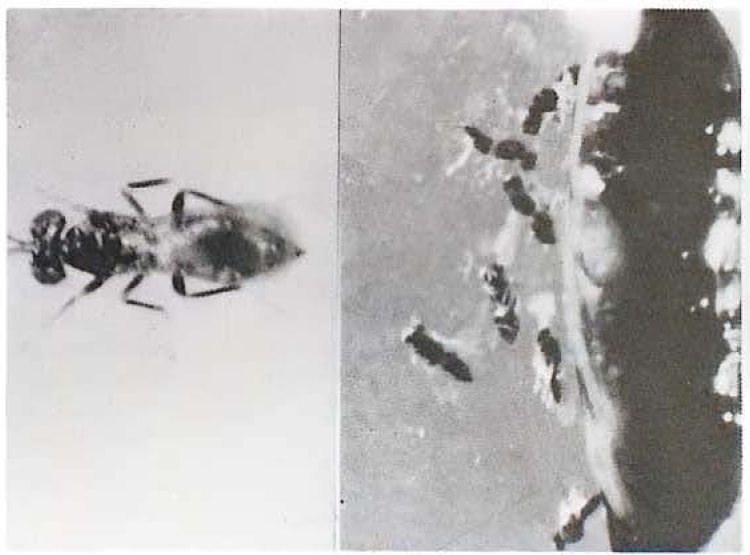
Aprostocetus sp. (Hymenoptera:Eulophidae) parasitoide de pupas de Neoleucinodes elegantalis. (Foto: Hery F. Viáfara M.)
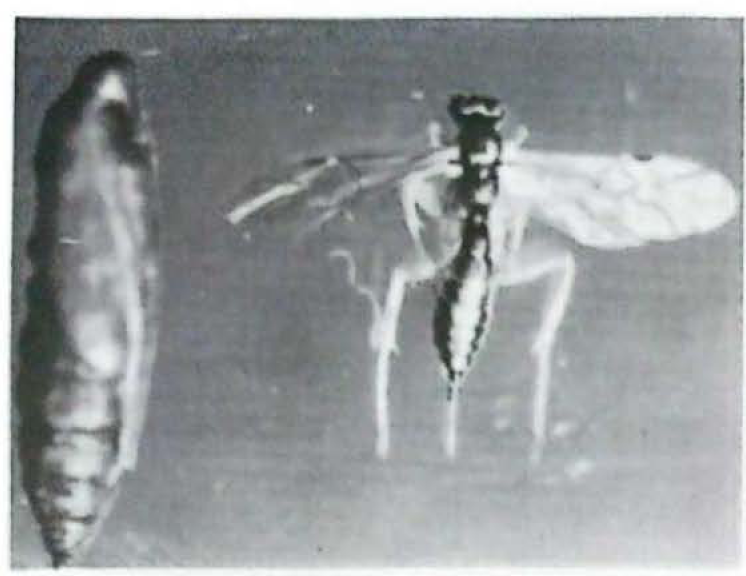
Hymenoptera: Ichneumonidae, emergido de pupas del pasador Neoleucinodes elegantalis. (Foto: Hery F. Viáfara M.).
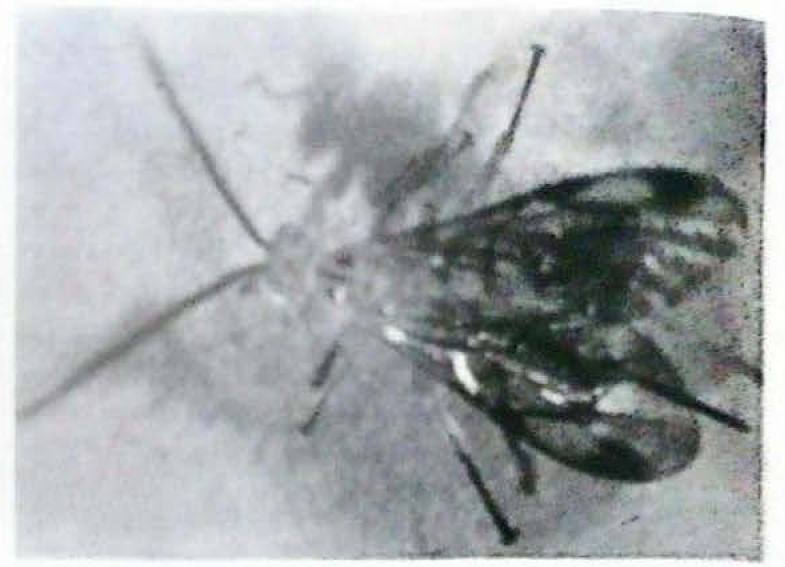
Hymenoptera: Ichneumonidae emergido de pupas del pasador Neoleucinodes elegantalis. (Foto: Hery F Viáfara M.).

Hymenoptera: Ichneumonidae emergido de pupas del pasador Neoleucinodes elegantalis. (Foto: Hery F. Viáfara M.).
Agradecimientos
Los autores agradecen a la Corporación Colombiana de Investigación Agropecuaria. CORPOICA. REGIONAL 5, al Comité de Cafeteros y Unidad de Asistencia Técnica Municipal (UMATA) de los municipios del Valle del Cauca, a los Productores de tomate de mesa, lulo y tomate de árbol del Cauca y Valle del Cauca y al Dr. Arnulfo Gómez-Carabalí. I.A. Ph. D.