
Abstract
In order to incorporate multiple resistance to three major insect pests of common bean, the leafhopper, Empoasca kraemeri Ross & Moore (Homoptera: Cicadellidae), the Mexican bean weevil, Zabrotes subfasciatus (Boheman) (Coleoptera: Bruchidae) and the bean pod weevil, Apion godmani (Wagner) (Coleoptera: Curculionidae), two improvement methods were compared: Gamete Selections (G.S.) in early generations and selection of advanced lines, developed by Recombinant Inbred Lines (R.I.L.) method. In the G.S. method, the initial selection for resistance to Z. subfasciatus was done by means of detection in F₂ of the presence of arcelin, conditioning factor of resistance to this insect. Progenies positive for arcelin were further submitted to various evaluations in the field for resistance to E. kraemeri in Colombia and to A. godmani in Mexico. Of the 216 progenies selected for the presence of arcelin, 17 F₈ lines were found to combine resistance to the three insects. The R.I.L. did not have an early evaluation and were submitted to the attack of the three insects in F₆. Laboratory tests with Z. subfasciatus and field trials with E. kraemeri and A. godmani allowed to identify 23 F₇ lines, which combined the three resistances. It was concluded that the R.I.L. method allows the incorporation of multiple insect resistance with considerable savings in resources and time.
Introducción
Dentro del grupo de las leguminosas comestibles, el frijol común, Phaseolus vulgaris L., es una de las más importantes debido a su amplia distribución en los cinco continentes y por ser complemento nutricional indispensable en la dieta alimenticia, principalmente en Centro y Sur América (Debouck e Hidalgo 1985), al punto que se estima que la producción y consumo de frijol en el mundo es mayor que todas las otras leguminosas de grano, exceptuando la soya y el maní (Singh 1996). Se estima que la producción de frijol en América Latina y África (principales consumidores de frijol) presentará un déficit aproximado de más de 3'150.000 toneladas para el año 2.000, de acuerdo con el crecimiento demográfico calculado para esa época (Janssen 1988).
Ante la necesidad de incrementar la producción agrícola, se oponen limitantes de áreas cultivables y de calidad de suelos, así como las pérdidas causadas por enfermedades, malezas e insectos, las cuales se estiman en un 37% de la producción a nivel mundial. De todas estas pérdidas causadas por factores bióticos, los insectos son responsables del 13% (Cardona 1996).
El consumo de insecticidas para el control de insectos es creciente en América Latina. El excesivo uso de insecticidas a su vez causa otros problemas como son resistencia de insectos a químicos, resurgencia, plagas secundarias que se convierten en primarias, contaminación del medio ambiente y contaminación de alimentos, atentando contra la salud de consumidores y de productores (Cardona 1996). Con todos estos problemas causados por la interferencia humana en la dinámica de poblaciones de insectos, el control genético por resistencia varietal aparece como una alternativa, base fundamental de sistemas de manejo integrado de plagas.
Entre las especies importantes para las cuales se ha explorado resistencia varietal como alternativa de control en frijol, se pueden citar el picudo de la vaina Apion godmani (Wagner) (Coleoptera: Curculionidae), el gorgojo Mexicano Zabrotes subfasciatus (Boheman) (Coleoptera: Bruchidae) y el lorito verde Empoasca kraemeri (Ross & Moore) (Homoptera: Cicadellidae).
El picudo de la vaina es una plaga importante en México, Guatemala, El Salvador, Honduras y norte de Nicaragua. Este insecto, que se presenta en épocas de lluvia que coinciden con las siembras, puede causar enormes pérdidas en los rendimientos del frijol, pudiendo llegar éstas a un 90%, produciendo un gran desequilibrio en la economía campesina (Schoonhoven y Cardona 1985b; Garza 1996).
El gorgojo Mexicano es una importante plaga del frijol almacenado en América Latina; este insecto es responsable del desequilibrio económico de muchos campesinos y comercializadores del grano, porque impide el almacenamiento por períodos largos para atender necesidades en épocas de escasez (Schoonhoven 1985).
El lorito verde es la plaga más común del frijol en América Latina. Este insecto se encuentra desde la Florida y norte de México, hasta el Perú y Brasil (Schoonhoven y Cardona 1985a). En condiciones de alta temperatura y sequía, sus poblaciones aumentan considerablemente, llegando a causar la pérdida total de las cosechas (Cardona 1994).
Por años se ha venido trabajando en el desarrollo de líneas de frijol resistentes a cada una de estas especies. Debido a que los tres insectos coexisten en algunas regiones de América Tropical, se decidió incorporar resistencia múltiple a ellos en líneas de frijol. Con el fin de poder diseñar programas exitosos para mejorar caracteres heredados cuantitativamente en frijol, es importante tener un conocimiento básico acerca del mecanismo genético que controla un carácter dado. Se sabe que la resistencia del picudo se debe a una combinación de mecanismos de antibiosis (hipersensibilidad morfológica, encapsulamiento de huevos y larvas de primer instar) y de antixenosis para oviposición (Garza 1996; Garza et al. 1996). La resistencia a picudo puede ser controlada por dos genes dominantes con niveles bajos a moderadamente altos de heredabilidad (Garza et al. 1996). Los dos genes (Agm, Agr) que gobiernan la resistencia a picudo son complementarios. Agm sólo, no confiere resistencia; Agr sólo, confiere únicamente resistencia intermedia. La alta resistencia conseguida resulta cuando los dos genes están presentes. Hasta ahora, estos genes han sido incorporados por el método de pedigree (Cardona y Kornegay 1998). Para el gorgojo Mexicano, el mecanismo responsable de la resistencia es la antibiosis, conferida por una proteína (arcelina) que causa efectos deletéreos en la biología y supervivencia del insecto (Cardona et al. 1989; Cardona y Kornegay 1998). La proteína se encuentra en los cotiledones de los materiales resistentes, es heredada a través de un solo gen dominante (herencia mendeliana simple) y puede ser transferida fácilmente por retrocruza (Cardona y Kornegay 1988; Cardona et al. 1990).
Tolerancia, y en menor grado antixenosis, son los mecanismos de resistencia al lorito verde, siendo la tolerancia más común e importante (Cardona y Kornegay 1998). La resistencia a este insecto es poligénica y heredada cuantitativamente (Kornegay y Temple 1986; Kornegay y Cardona 1988). Esta resistencia se ha logrado incorporar mediante el método de selección recurrente.
Un carácter muy importante a tener en cuenta cuando se hace resistencia varietal es el rendimiento. Éste, siendo cuantitativo y controlado por muchos genes (Davis 1985), puede presentar interacción con el ambiente, reduciendo eficiencia en las evaluaciones y selecciones, además de presentar una moderada correlación entre el puntaje de daño por alimentación de empoasca y el rendimiento no protegido (Kornegay y Cardona 1988).
El uso de selección por gametos y las pruebas en generaciones tempranas es muy conveniente cuando se desea mejorar para un número máximo de caracteres agronómicos. Además, las selecciones tempranas incrementan la frecuencia de líneas resistentes e intermedias a medida que avanzan las generaciones (Singh et al. 1998). Asimismo, Davis (1985) comenta que seleccionar por un carácter cuantitativo como rendimiento debe comenzar en la generación más temprana posible.
Por otro lado, el método de descendencia de semilla única para generar líneas puras recombinantes se usa para avanzar tantos genotipos diferentes y potencialmente favorables como sea posible hasta un estado más homocigoto en un espacio mínimo (Kornegay y Cardona 1988). Esta práctica aumenta la oportunidad de hallar mayores niveles de recombinantes resistentes, especialmente con caracteres cuantitativamente heredados.
Singh et al. (1998) mencionan que cuando un carácter es controlado por uno o dos genes dominantes y sus padres donantes contribuyen genéticamente de manera proporcional a la cruza final, podría esperarse una alta proporción de líneas avanzadas que tengan resistencia necesaria aunque no fueran evaluadas y seleccionadas en generaciones tempranas.
Durante muchos años el CIAT ha generado con éxito materiales con resistencias independientes a cada insecto, pero teniendo en cuenta que éstos coexisten en algunas regiones del Trópico, se decidió utilizar materiales con resistencias independientes para crear líneas élites con resistencia múltiple. Otro objetivo que se busca es economizar tiempo y recursos mediante la comparación de dos métodos de evaluación, uno en generaciones tempranas y otro en generaciones avanzadas, ya que el mejoramiento tradicional ha implicado la utilización de muchos recursos, tiempo y espacio. De igual manera, se busca introducir el carácter de rendimiento en los materiales para que puedan ser utilizados por los agricultores.
Materiales y Métodos
Se usaron ocho líneas de frijol común previamente identificadas por su resistencia a alguno de los insectos materia del estudio. Estas líneas fueron EMP 233, EMP 268, EMP 423 y EMP 445 con resistencia a lorito verde; RAZ 2 y RAZ 51 como resistentes al gorgojo Mexicano y Amarillo 154 y Dicta 105 como resistentes al picudo de la vaina; las cuales tenían características fenotípicas favorables como color y hábito, además de mostrar buena respuesta al carácter de rendimiento.
Se realizaron cruzas simples y con la progenie F₁, se hicieron cruzas dobles que se denominaron A y B. A estas F₁ se les realizó una prueba bioquímica (electroforesis) para determinar la presencia de arcelina I (Arc) (Cardona et al. 1990). Las progenies F₂ Arc⁺ fueron cruzadas nuevamente con EMP 423 y Dicta 105 para crear las cruzas triples D y K. A las progenies F₁ de estas cruzas triples también se les determinó la presencia de Arc mediante serología.
Los materiales de este ensayo se dividieron en dos grupos para comparar los dos métodos. El primer grupo de progenies positivas para arcelina fue evaluado en campo usando Selección de Gametos (S.G.) desde F₂ hasta F₅ para E. kraemeri y en F₄ y F₆ se evaluaron para Z. subfasciatus y A. godmani. El segundo grupo de progenies, no seleccionadas por la presencia de arcelina, fue avanzado usando descendencia de semilla única para generar Líneas Puras Recombinantes (L.P.R.) F₆, las cuales se evaluaron por resistencia a E. kraemeri y Z. subfasciatus y posteriormente en F₇ por resistencia a los tres insectos.
Se utilizaron diferentes sitios de evaluación (Colombia, Honduras y México) debido a que en Colombia no hay presencia natural del picudo de la vaina. Para el lorito verde y el picudo de la vaina, las evaluaciones se hicieron en campo y para el gorgojo Mexicano se realizaron en laboratorio.
Las escalas de evaluación que se utilizaron fueron las siguientes: para E. kraemeri, los materiales que mostraron calificación de daño ≤ 7.3, en una escala visual de 1 a 9, fueron clasificados como resistentes o intermedios mientras que aquellos con calificación ≥ 7.4 fueron clasificados como susceptibles. Para A. godmani, los materiales que presentaron un porcentaje de daño en la semilla inferior al 40% fueron considerados resistentes o intermedios. En el caso de Z. subfasciatus, se utilizaron dos escalas: porcentaje de emergencia de adultos y porcentaje de semilla sana. Fueron resistentes o intermedios aquellos materiales con porcentajes de emergencia de adultos menores de 40%. En términos de daño en la semilla se clasificaron como resistentes o intermedios los materiales con niveles de daño inferiores al 40%.
Con el fin de evitar algún escape en las diferentes evaluaciones, la presión de infestación por E. kraemeri en condiciones de campo promedió 10 ninfas por trifolio. Se contó además con un testigo tolerante (ICA Pijao) y un testigo susceptible (BAT 41) como indicadores de una infestación uniforme. Para Z. subfasciatus se infestaron tres repeticiones de 30 semillas con seis parejas de insectos por repetición, garantizando así una buena infestación y la no competencia por alimentación entre larvas. También fueron utilizadas líneas RAZ como testigos resistentes y la línea ICA Pijao como testigo susceptible. Para las evaluaciones con A. godmani se utilizó como testigo resistente a J-117 y como testigo susceptible a Canario 107. Estos testigos fueron distribuidos dentro y alrededor del ensayo. El no escape se comprobó debido a que el testigo susceptible presentó en promedio un porcentaje de semilla dañada igual o superior al 30% a través del campo.
Las evaluaciones para seleccionar materiales con resistencia a E. kraemeri y a A. godmani se hicieron con un diseño de bloques completos al azar con tres repeticiones. Para evaluar la resistencia a Z. sufasciatus se utilizó un diseño completamente al azar. Para comprobar el carácter rendimiento bajo presión de E. kraemeri, los materiales se evaluaron en un diseño de bloques completos al azar con tres repeticiones en presencia y ausencia del insecto, evaluando el daño mediante una escala visual y midiendo los rendimientos de las líneas en estudio. Los resultados fueron analizados mediante un programa estadístico modificado para SAS.
Resultados y Discusión
La determinación de la proteína Arc⁺ en S.G. identificó 216 materiales, siendo éste el 50.5% del total evaluado (Tabla 1). Esto está de acuerdo con la teoría que expresa que cuando un carácter es controlado por un sólo gen en la primera generación, el 50% de los materiales herederá dicho carácter.
No. de semillas F₂ seleccionadas por la presencia (Arc⁺) o ausencia (Arc⁻) de arcelina. Selección inicial para resistencia al gorgojo Mexicano
A = (EMP423 x Amarillo 154) (RAZ 2 x EMP268)
B = (EMP445 x RAZ 51) (APN 18 x EMP423)
D= EMP423 x A
K = Dicta 105 x A
Después de cuatro ciclos de selección gamética mediante la escala visual de daño para E. kraemeri, se seleccionaron 26 materiales resistentes e intermedios, mientras que con dos evaluaciones después de F₄ en L.P.R. se seleccionaron 35 materiales (Tabla 2). Esta diferencia favorable a L.P.R. se dió posiblemente porque al avanzar genotipos por descendencia de semilla única, aumentó la posibilidad de hallar mayores niveles de recombinantes resistentes, especialmente al trabajar con caracteres heredados en forma cuantitativa.
Número de líneas F₅ de frijol seleccionadas por tolerancia al lorito verde siguiendo dos métodos de selección
A = (EMP423 x Amarillo 154) (RAZ 2 x EMP268)
B = (EMP445 x RAZ 51) (APN 18 x EMP423)
D = EMP423 x A
K = Dicta 105 x A
I, intermedio; R, resistente
Para las S.G., después de tres selecciones por Z. subfasciatus, se obtuvieron 34 materiales resistentes e intermedios, mientras que en L.P.R. se generaron 47 líneas con resistencia, luego de dos selecciones a partir de F₆ (Tabla 3). La mayor cantidad de materiales generados por L.P.R. se debió a que la homocigosis alcanzada por ellos fue mayor que en las S.G., debido al método utilizado para adelantar las generaciones. Esto implicó un mayor número de recombinantes en L.P.R., mientras que en S.G., por haber una continua recombinación de semilla por línea, se generó heterosis, lo cual disminuyó la proporción de materiales homocigotos con buenos niveles de resistencia.
Número de líneas F₆ de frijol seleccionadas por resistencia al gorgojo Mexicano siguiendo dos métodos de selección
A = (EMP423 x Amarillo 154) (RAZ 2 x EMP268)
B = (EMP445 x RAZ 51) (APN 18 x EMP423)
D= EMP423 x A
K = Dicta 105 x A
I, intermedio; R, resistente
Las dos evaluaciones realizadas para resistencia a A. godmani permitieron seleccionar 23 materiales resistentes e intermedios por el método S.G., mientras que una evaluación después de F₆ realizada en las L.P.R. seleccionó 37 materiales (Tabla 4). Estas diferencias que favorecieron a L.P.R. posiblemente se debieron a que los dos genes que controlan este carácter presentan una heredabilidad baja a moderadamente alta, además son complementarios. Por lo tanto, al haber autofecundación sucesiva, la probabilidad de encontrar estos genes juntos fue mayor en las L.P.R.
Número de líneas F₇ de frijol seleccionadas por resistencia al picudo de la vaina siguiendo dos métodos de selección
A = (EMP423 x Amarillo 154) (RAZ 2 x EMP268)
B = (EMP445 x RAZ 51) (APN 18 x EMP423)
D= EMP423 x A
K = Dicta 105 x A
I, intermedio; R, resistente
Los materiales resistentes a E. kraemeri generados por S.G. y L.P.R presentaron en promedio, calificaciones muy similares en la escala visual utilizada para evaluar el daño de este insecto (Tabla 5). Esto se debió a que cuando se utilizan selecciones tempranas para transferir una característica poligénica, el nivel de homocigosis deseado se presenta en generaciones avanzadas como F₈, mientras que en las L.P.R. pudo presentarse dicha homocigosis en una generación más temprana, lo cual permitió observar mayor número de líneas resistentes en F₆. Los niveles de resistencia alcanzados por algunas líneas como 45 B 59, 60 B 84, 79 B 117 generadas por S.G. y 30 A 299, 61 B 697 y 156 B 356 generadas por L.P.R. fueron superiores a los de los padres (líneas EMP) y testigos resistentes, indicando la ganancia genética lograda por estas líneas en cuanto al daño de E. kraemeri se refiere.
Daño causado por lorito verde (Empoasca kraemeri, EMP), porcentaje de emergencia de adultos de gorgojo Mexicano del frijol (Zabrotes subfasciatus, ZAB), y porcentaje de semilla dañada por el picudo de la vaina (Apion godmani, APN), en las mejores líneas para resistencia múltiple obtenidas por dos métodos de selección
Cruces: A = (EMP423 x Amarillo 154) x (RAZ 2 x EMP 268)
B = (EMP 445 x RAZ 51) x (APN 18 x EMP 243)
D = EMP 423 x A
K= DICTA 105 x A
Materiales utilizados como fuentes de resistencia: Líneas EMP para Empoasca; RAZ 2 y RAZ 51 para gorgojo Mexicano del frijol; Amarillo 154, APN 18, DICTA 105 para el picudo de la vama.
RAZ 44 es el testigo resistente mejorado para Zabrotes; ICA Pijao es el testigo susceptible para Zabrotes y el testigo tolerante para Empoasca; BAT 41 es el testigo susceptible para Empoasca; J-117 es el testigo resistente a Apion, Desarrural y Canario 107 son los testigos susceptibles para Apion
Los materiales resistentes a Z. subfasciatus generados por el método S.G. mostraron en promedio un porcentaje de emergencia de adultos similar a las líneas generadas por L.P.R. (Tabla 5). También se observó que con los dos métodos se logra ganancia genética con respecto a los padres (líneas RAZ) además de un considerable avance con relación al testigo susceptible (ICA Pijao). El testigo resistente RAZ 44 presentó un porcentaje de emergencia de adultos muy inferior a los materiales en estudio, debido a la continua purificación a la que ha sido sometido dicho testigo. La ganancia en las líneas generadas por S.G. (120 D 95) se debió a que, como la herencia para Z. subfasciatus es gobernada por un gen simple, la selección temprana por serología y/o electroforesis garantizó que los materiales que continuaron la selección, presentaran el gen que gobierna la proteína. En cada generación la presión de selección permitió obtener materiales con niveles altos y estables de la proteína, y por ende de resistencia, en generaciones avanzadas. Sin embargo, en las L.P.R. también se obtuvo ganancia genética (30 A 299, 60 B 547) debido a que, como en F₂ se presentó un alto número de recombinantes, la probabilidad de encontrar materiales con niveles altos de arcelina es muy grande, lo cual permitió resaltar la utilidad de los dos métodos para mejorar materiales con herencia mendeliana simple.
En el caso de los materiales resistentes a A. godmani generados por el método S.G., el porcentaje de semilla dañada fue significativamente inferior al testigo susceptible (Canario 107) y similar al testigo resistente (J-117). Con las selecciones tempranas no se observó en promedio ganancia genética frente a los padres ya que éstos tuvieron menor calificación de porcentaje de daño que los materiales generados por S.G. Sin embargo, la diferencia entre el daño de las líneas S.G. y los padres resistentes es mínima, comparada con la gran diferencia que existe respecto al testigo susceptible (Tabla 5). Los niveles de resistencia a A. godmani de los materiales generados por L.P.R. fueron comparables a los de los padres y el testigo resistente J-117. Esto fue debido a que al seleccionar materiales después de F₄ en L.P.R. se generó una mayor cantidad de recombinantes que presentaron los dos genes complementarios que gobiernan esta resistencia. Los materiales obtenidos por S.G. lograrían esta homocigosis con más de dos ciclos de selección (Tabla 5).
Comparando los resultados de las tres evaluaciones en F₈ (Fig. 1), se seleccionaron materiales con tres resistencias, 17 generados por selección de gametos y 24 obtenidos por descendencia de semilla única. Además, 15 materiales de S.G. y 22 de L.P.R. presentaron dos resistencias; dos S.G. y cuatro L.P.R. sólo mostraron una resistencia. Los demás fueron susceptibles.
Con respecto al carácter de rendimiento, se pudo observar que las S.G. presentaron promedios de rendimiento significativamente mayores que las L.P.R. y que el testigo susceptible y estadísticamente iguales al testigo resistente bajo el ataque de E. kraemeri (Tabla 6). Aunque los rendimientos en ausencia del insecto no fueron significativamente diferentes, las S.G. presentaron mayor rendimiento promedio que las L.P.R. El testigo resistente (ICA Pijao) mostró mayor rendimiento bajo protección debido a su mejor adaptación y a que la presión de selección ejercida para combinar las tres resistencias implicó una disminución en los rendimientos. Comparando los dos ambientes se generaron los porcentajes de pérdida, los cuales mostraron que las selecciones gaméticas podrían perder menos en su rendimiento frente al ataque del insecto que el testigo resistente, mientras que las L.P.R. debido a su alto porcentaje de pérdida necesitarían del control químico para obtener buenos rendimientos. En cuanto al índice de susceptibilidad, las líneas S.G. produjeron lo suficiente para recuperarse del ataque del insecto, lo cual permitiría al agricultor obtener buenos rendimientos sin el uso de agroquímicos, mientras que las L.P.R. necesitarían el control químico suplementario. Lo anterior se explica porque el carácter de rendimiento presenta interacción con el ambiente, y si no se realizan selecciones tempranas, se reduce la eficiencia en las evaluaciones y selecciones debido a la correlación existente entre el puntaje de daño por alimentación de E. kraemeri y el rendimiento no protegido.
Rendimientos promedios entre dos metodos de selección de líneas F₈, los padres y testigos y sus respectivos porcentajes de pérdida e indice de susceptibilidad para E. kraemeri
Cardona y Kornegay (1998).
Las medias en cada columna seguidas por la mismas letras no son significativamente diferentes al 5% (Prueba de Rangos Múltiples de Duncan).
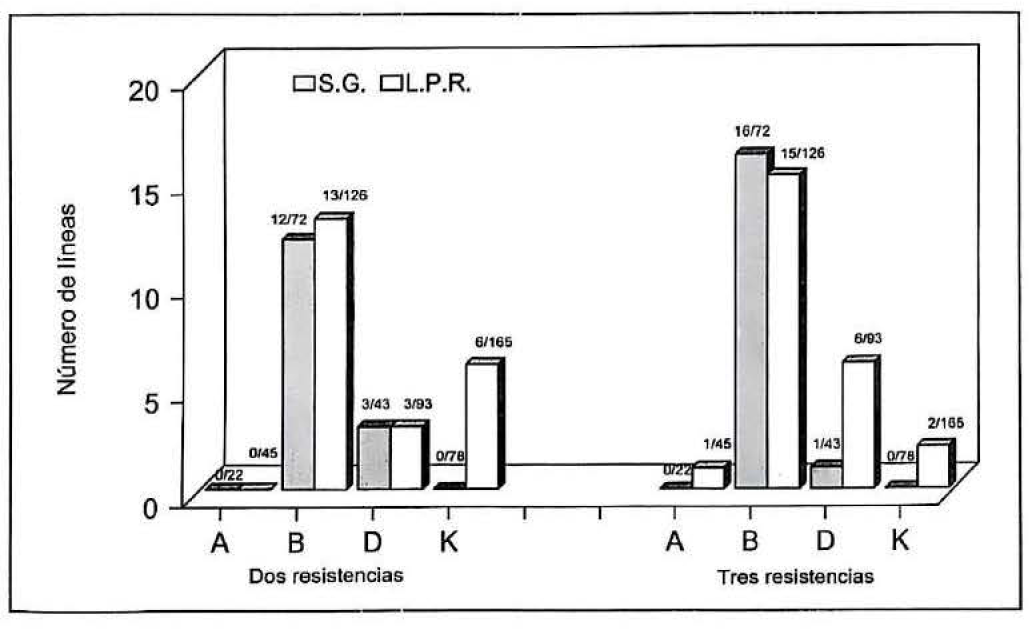
Líneas de frijol con dos y tres resistencias generadas mediante los métodos de selección de gametos y desarrollo de líneas puras recombinantes. Número de progenies seleccionadas respecto al total evaluado.
Conclusiones
• Se concluye que con el método L.P.R. se obtuvieron más líneas resistentes que con el método S.G. y se logró incorporar resistencia a los tres insectos con un considerable ahorro de recursos y tiempo. Por el método L.P.R. se generaron padres potenciales que poseen resistencia múltiple útil en programas de mejoramiento.
• Con el método de S.G. también fue posible incorporar resistencia múltiple y mantener, al mismo tiempo, buenos potenciales de rendimiento. Las líneas obtenidas por S.G. presentaron menores pérdidas en rendimiento frente al ataque de E. kraemeri, ofreciendo un ahorro considerable en insumos (plaguicidas) para los agricultores. Aunque pueden también ser usadas como padres potenciales en mejoramiento, su obtención demanda mayor trabajo y recursos.