
Abstract
The cassava mealybug Phenacoccus herreni Cox Williams (Sternorrhyncha: Pseudococcidae) is an important pest of cassava, Manihot esculenta Crantz (Euphorbiaceae), especially during drought, when insect population's increase. This phenomenon might be a response to biochemical changes in cassava leaves induced by water deficiency, which enhances insect development. In order to verify this phenomenon it was first necessary to study its feeding behavior. Observation of the mouthpart by scanning electron microscopy and fase contrast microscopy were carried out and the feeding activity was studied by electropenetrography (EPG). Insect biological development and reproductive rate, and also infested leaf aminoacid composition from cassava plants grown with differential irrigation levels (well watered and deficiently watered) were evaluated. The influence on P. herreni development of the aminoacids that increased their proportion in water stressed plants was evaluated using an artificial diet. The results showed that P. herreni is mainly a phloemophagous insect with piercing-sucking mouthpart that is present in females throughout their life cycle. Male insects showed the same mouthpart only till the second ínstar. The fact that the females maintain their piercing-sucking mouthpart and they feed on phloem sap throughout their life cycle, we suggest that the females are more responsible for cassava damage and therefore subsequent studies were done on females. Water stress in plants induced a reduction in the duration on female development and an increase in fecundity, intrinsic natural growth rate (rm) and in weight. Leaf aminoacid composition showed a more important increase in the proportion of free asparagine. Using an artificial diet it was shown that this aminoacid may play an important role during the development of P. herreni. In fact, its absence in the diet induced an insect development delay.
Introducción
En Sur América, el piojo harinoso Phenacoccus herreni Cox & Williams es un insecto plaga que afecta a la yuca Manihot esculenta Crantz (Euphorbiaceae) en especial durante la estación seca, donde la población del insecto se incrementa (Bellotti et al. 1983; Noronha 1990). Este fenómeno sugiere la existencia de algún tipo de interacción entre los compuestos sintetizados por la planta durante la estación seca, que quizá favorezcan desde el punto de vista nutricional a los insectos, traduciéndose en un incremento de la población. Se ha observado, en la mayoría de las plantas, que el estrés hídrico induce un incremento en la concentración de compuestos tales como aminoácidos, carbohidratos y ácidos orgánicos, cuya acumulación les permite disminuir el potencial osmótico de las células (Lamaze et al. 1996). De esta forma, la mayor concentración de este tipo de compuestos en las plantas puede favorecer el desarrollo de los insectos (Mattson y Haack 1987).
Con el fin de estudiar la repercusión de los cambios bioquimicos en hojas de yuca inducidos por déficit hidrico sobre el desarrollo de P. herreni, se empezó por conocer el comportamiento alimenticio del insecto.
El comportamiento alimenticio de los insectos de la familia Pseudococcidae no ha sido tan ampliamente descrito como el de Aphididae, sin embargo, todos los autores han identificado un comportamiento alimentario floemófago para la familia Pseudococcidae (Pesson 1944; Grassé 1951; Albrigo y Brooks 1977; Campbell 1990; Calatayud et al. 1994).
Trabajos anteriores sobre diferentes especies de piojo harinoso, describen en detalle el aparato bucal de las hembras, no asi, el de los machos. Este hecho llama la atención, ya que P. herreni presenta un marcado dimorfismo sexual, tanto asi, que machos y hembras se creia que pertenecian a especies distintas, lo que llevó a pensar que tanto las estructuras bucales como su comportamiento alimenticio, podrían ser diferentes. La literatura describe que los piojos adultos macho tienen un aparato bucal atrofiado (Grassé 1951; Coquis y Salazar 1975; Bellotti et al. 1983), sin embargo no se conoce cómo es la estructura de su aparato bucal durante las diferentes fases metamórficas.
La microscopia óptica ha sido la herramienta más utilizada en la descripción de la trayectoria de los estiletes de los homópteros dentro del tejido vegetal. McLean y Kinsey (1964) idearon un sistema de registro eléctrico para estudiar el movimiento de penetración de los estiletes en el tejido vegetal. La técnica consiste en hacer un seguimiento de los cambios de potencial dentro de un circuito eléctrico, a lo largo del recorrido de los estiletes dentro del tejido vegetal. Posteriormente, ésta fue modificada con el fin de detallar más la actividad de los estiletes y su interacción con los tejidos de la planta, y fue llamada electropenetrografia (EPG)(Tjallingii 1978). Las señales registradas mediante la metodología son obtenidas gracias a las propiedades eléctricas de la planta y a los eventos que se presentan en los estiletes del insecto y en sitios próximos a ellos (Tjallingii 1985). De tal manera, que las señales pueden tener su origen en las fuerzas electromotrices (fem) producidas por la actividad mecánica de los estiletes o pueden ser producto de la resistencia inducida por el insecto que varia de acuerdo a la fuente eléctrica externa de los canales de los estiletes. Esta técnica ha sido utilizada en el estudio del comportamiento alimenticio de áfidos (Tjallingii 1978; Kimmins y Tjallingii 1985), del piojo harinoso de la yuca Phenacoccus manihoti (Calatayud et al. 1994) y especialmente en el estudio de la resistencia de las plantas a los áfidos (Kimmins 1989; Montlor y Tjallingii 1989).
En este trabajo se observó el desarrollo del aparato bucal de P. herreni, desde el segundo instar hasta el estado adulto de hembras y machos por microscopia electrónica de barrido y óptica de contraste de fases. Simultáneamente, se estudió la actividad de los estiletes de hembras y machos de P. herreni, dentro los tejidos de hojas de yuca utilizando la técnica de electropenetrografia (EPG), con el fin de identificar el comportamiento alimenticio de este insecto.
Después de esta identificación y con base en lo anterior, en el presente trabajo se estudió también el desarrollo y la tasa reproductiva del piojo harinoso criado sobre plantas de yuca, tanto en condiciones normales de riego como de déficit hídrico, al igual que los cambios en la composición aminoacídica de hojas de ambos tipos de planta. Adicionalmente, se verificó con dietas artificiales, la influencia de los aminoácidos cuya proporción aumentó en plantas bajo déficit hidrico, sobre el desarrollo y crecimiento de P. herreni.
Materiales y Métodos
Matcríllf animal.
Se utilizaron insectos de P. herreni colectados de la cría masiva del CIAT, Palmira, Valle. Los insectos se mantuvieron en condiciones de invernadero sobre plantas de yuca de la variedad CMC-40 a una temperatura media de 30°C, y humedad relativa media del 45%. Para estudiar la influencia del déficit hídrico en plantas de yuca sobre el desarrollo de P. herreni, los insectos se criaron sobre plantas con irrigación diferencial durante cuatro generaciones, iniciando con insectos de primer instar.
Material vegetal.
Se utilizaron plantas de yuca M. esculenta de un mes de edad, de la variedad CMC-40 cultivadas en casa de malla. Las plantas utilizadas se transportaron de la casa de malla al laboratorio donde se realizó la EPG con 24 horas de anticipación al experimento, con el fin de preadaptarlas a las nuevas condiciones de luz y temperatura. Para estudiar la influencia de plantas en déficit hidrico sobre el desarrollo de P. herreni, se utilizaron plantas de yuca de un mes de edad, de la variedad CMC-40, sometidas a riego diferencial: plantas control con 400 ml de H₂O (3 veces por semana) y plantas con déficit hídrico irrigadas con 100 ml de H₂O (1 vez por semana). Al cabo de un mes de régimen hidrico las plantas estuvieron listas para ser infestadas con insectos de primer instar.
Observación del aparato bucal de P. herreni por microscopía electrónica de barrido.
Se colectaron individuos a partir del segundo instar hembras y machos hasta el estado adulto, ya que en el primer instar de P. herreni no se puede diferenciar el sexo. Seguidamente, los insectos se introdujeron en una solución fijadora compuesta de buffer fosfato (K₂HPO₄ 0.1 M pH 7.2) y glutaraldehido al 2.5%, en la cual permanecieron por un período de 20 h. Posteriormente, los insectos se postfijaron en solución de tetróxido de osmio al 2% por 1 h. Luego, las muestras se deshidrataron y se pasaron sucesivamente por soluciones de etanol al 25, 50, 75, 90 y 100% de concentración, cada una durante 20 minutos. Para retirar la cera y evitar que ésta oscureciera las micrografias, las muestras se introdujeron sucesivamente en mezclas de xilol-etanol en proporción 1:1 y 2:1 por 30 min, y por último en xilol durante 3 h. Se adicionó nuevamente xilol-etanol (1:1) por 30 min y por último se dejaron en etanol al 100%. Después, se secaron por punto crítico y se recubrieron con oro-paladio hasta su observación al microscopio electronico de barrido (Jeol JSM-820).
Observación del aparato bucal de P. herreni por microscopía óptica de contraste de fases.
Se observaron piojos desde segundo instar hasta estado adulto de macho y hembra de P. herreni. Los piojos se colocaron en una mezcla de xilol-fenol 1:1 durante 12 h para remover la cera y aclarar la muestra, y de esta forma, facilitar la observación interna por transparencia de los estiletes a lo largo de su desarrollo ninfal. Seguidamente, en una lámina portaobjetos se colocó la muestra y sobre esta se dejó caer una gota de lactofenol e inmediatamente se selló con una lámina cubreobjetos. Los micropreparados se observaron en un microscopio Leitz-Aristoplan.
Actividad de alimentación de P. herreni por electropenetrografia (EPG).
La penetración de los estiletes fue monitoreada mediante un sistema EPG-DC de cuatro canales con una resistencia introducida de 1 GΩ (Modelo Giga 4 DACQ 95, Holanda). En el dorso de los insectos se fijó un filamento de oro de 5 cm de largo y 17.5 m de diámetro con una gota de pintura de plata. Antes de ser colocados sobre el haz de la hoja, los insectos fueron conectados a un amplificador de señales. Los ensayos se llevaron a cabo en una caja de Faraday a 30°C. En todos los experimentos se utilizó un sistema de registro de datos Mac Adios 8 ain que convierte y archiva las señales EPG. Las señales archivadas fueron transferidas después a un programa para MacIntosh MacStylet (Febvay et al. 1996). La duración de los registros fue de 8 h y el tiempo 0 equivale al momento en que los insectos son colocados sobre la hoja. Por cada estadío de desarrollo de macho y hembra, se realizaron 5 EPG's. Los porcentajes medios de permanencia de cada parámetro EPG fueron comparados de acuerdo con el estadio de desarrollo y el sexo, mediante tabla de contingencia.
Influencia del déficit hídrico de la planta hospedera sobre el desarrollo de P. herreni.
El desarrollo y crecimiento de los piojos, criados sobre 5 plantas control y 5 plantas con estrés hídrico, se evaluó con base en el tiempo total de maduración de las hembras, desde primer instar hasta el adulto maduro (i.e. hembras que empiezan a formar ovisaco), la fecundidad (número de huevos por hembra), la tasa intrinseca de crecimiento natural (rm) según la formula de Birch (1948) y el peso de hembras adultas. Al final de la evaluación del desarrollo, las hojas maduras donde estaban posados los piojos, provenientes de plantas control y en estrés hídrico, fueron procesadas independientemente para su análisis en aminoácidos según (Calatayud et al. 1998).
Los datos obtenidos fueron evaluados mediante análisis de varianza, seguido de un análisis 2x2 de las medias con la prueba PLSD de Fisher. Antes de usar el análisis de varianza, se verificó la normalidad de las distribuciones y la igualdad de la varianza con las pruebas de Wald-Wolfowitz y de F de igualdad de varianza, respectivamente. Se necesitó solamente transformar el peso (en 1/X) para obtener estas condiciones necesarias para el uso correcto del análisis de varianza. Los porcentajes medios de aminoácidos obtenidos para las hojas de plantas control y en estrés hídrico, fueron comparados mediante tabla de contingencia.
Influencia del cambio de la composición aminoácidica de una dieta artificial sobre el desarrollo y crecimiento de P. herreni.
La dieta utilizada se preparó con base en la descrita por Febvay et al. (1988), para áfidos y adaptada para piojo harinoso por Calatayud et al. (1998). Para verificar el papel indispensable de algunos aminoácidos sobre el desarrollo de P. herreni, se prepararon diferentes dietas que carecían de un aminoácido (cuya proporción se vió aumentada en plantas sometidas a estrés hídrico) más una control (i.e. con todos los aminoácidos). Las dietas se infestaron con individuos de primer instar de la cria masiva. El bioensayo se mantuvo hasta cuando la mayoría de las hembras sobre las dietas control presentó ovisaco. Luego las hembras procedentes de cada tipo de dieta se pesaron en una microbalanza para evaluar su crecimiento. Después de transformar los datos en Ln ((1/X)+10) para requirir la igualdad de la varianza, se compararon mediante análisis no parámetrico con la prueba de U de MannWhitney, debido a que las distribuciones no fueron normales.
Resultados y Discusión
Observación microscópica del aparato bucal de P. herreni durante sus fases de desarrollo.
Se registraron micrografías por microscopía electrónica de barrido de machos y hembras durante sus fases de desarrollo. El análisis de estas impresiones permitió observar las diferencias en el desarrollo del aparato bucal de P. herreni entre insectos de los dos sexos. La evolución del aparato bucal no mostró diferencias externas entre el segundo y tercer ínstar de insectos macho (Fig. 1), y hembra durante todas sus fases de desarrollo (resultado no presentado). Los aparatos bucales presentaron piezas con orientación hipognata, el rostro insertado en el limite posterior de la cabeza y labium bien desarrollado, lo que concuerda con descripciones anteriores (Pesson 1944). Por el contrario, el cuarto instar de machos presentó una marcada retracción del aparato bucal externo, que en el estado adulto, termina con un atrofiamiento total, formando en su reemplazo una invaginación (Fig. 2). Este resultado muestra que la involución del aparato bucal de los machos se inicia en el cuarto estadio de desarrollo. Para observar la parte interna del aparato bucal, se obtuvieron impresiones por microscopia de contraste de fases de machos y hembras, durante sus fases de desarrollo. El análisis de las imágenes corroboró las observaciones realizadas por microscopia electrónica de barrido, mostrando la involución del aparato bucal de los machos a partir del cuarto instar (resultado no mostrado). La facilidad de observar internamente el aparato bucal de los insectos por transparencia, utilizando la técnica contraste de fases, permitió evidenciar que en los machos de tercer instar en estado temprano de desarrollo se observan muy tenues los estiletes (Fig. 3B). Además, insectos macho de tercer instar en estado tardío de desarrollo presentan piezas bucales en estado de involución, las cuales carecen de estiletes (Fig. 3C), y por consiguiente el aparato bucal en este estadio no es funcional. Estos resultados sugieren que a partir de este momento los machos no necesitan alimento para desarrollarse, ya que al aislar insectos macho de tercer instar en cajas Petri en ausencia de alimento, estos pudieron desarrollarse hasta el estado adulto (resultado no presentado).
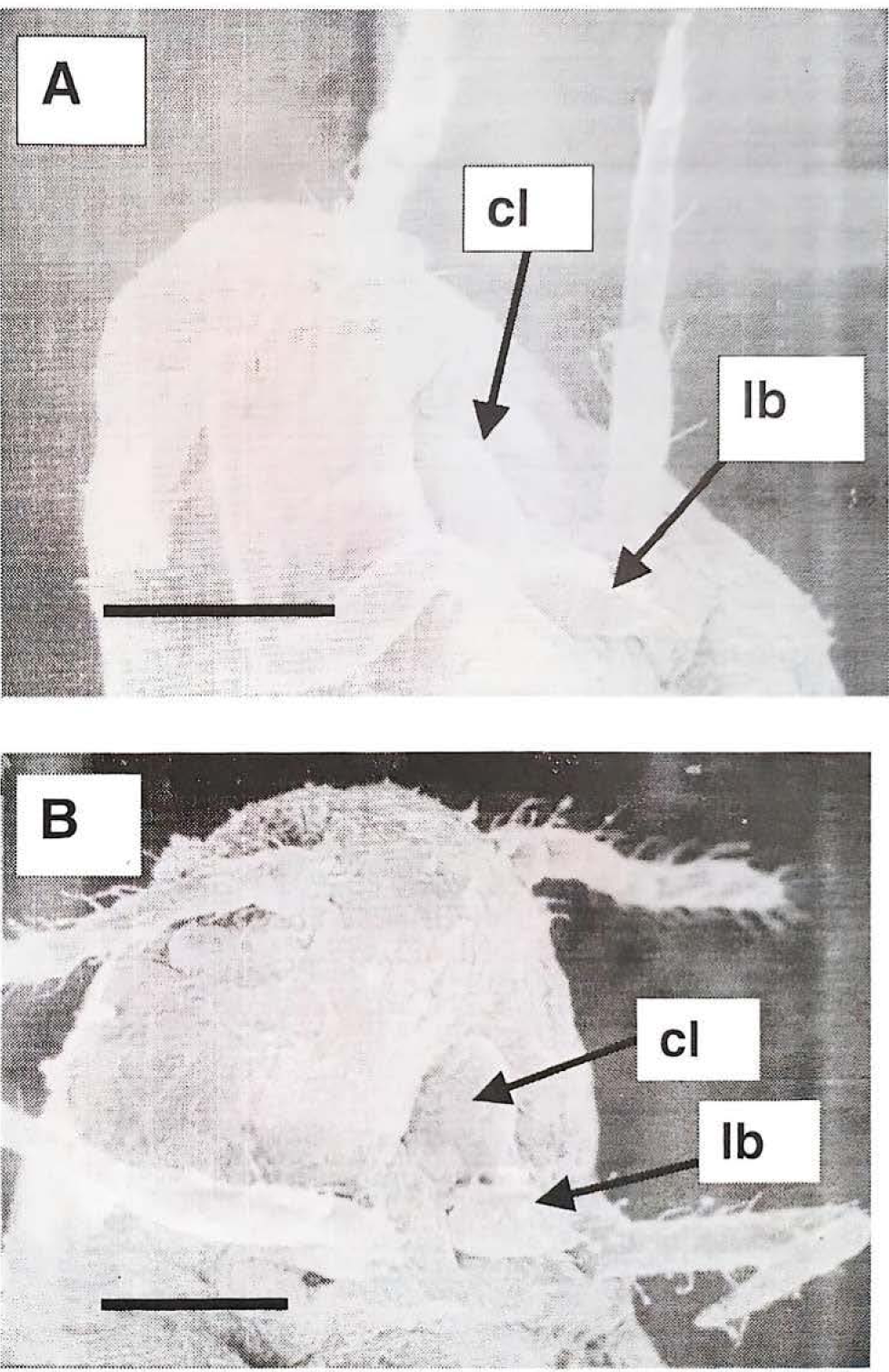
Micrografias en microscopia electrónica de barrido. A) Macho de segundo ínstar y B) macho de tercer instar. Las micrografias muestran que el macho en estos estadios de desarrollo presenta un aparato bucal externo completo (con labium (lb) y clipeus (cl) visibles) y comparable al de las hembras (raya = 50 μm).
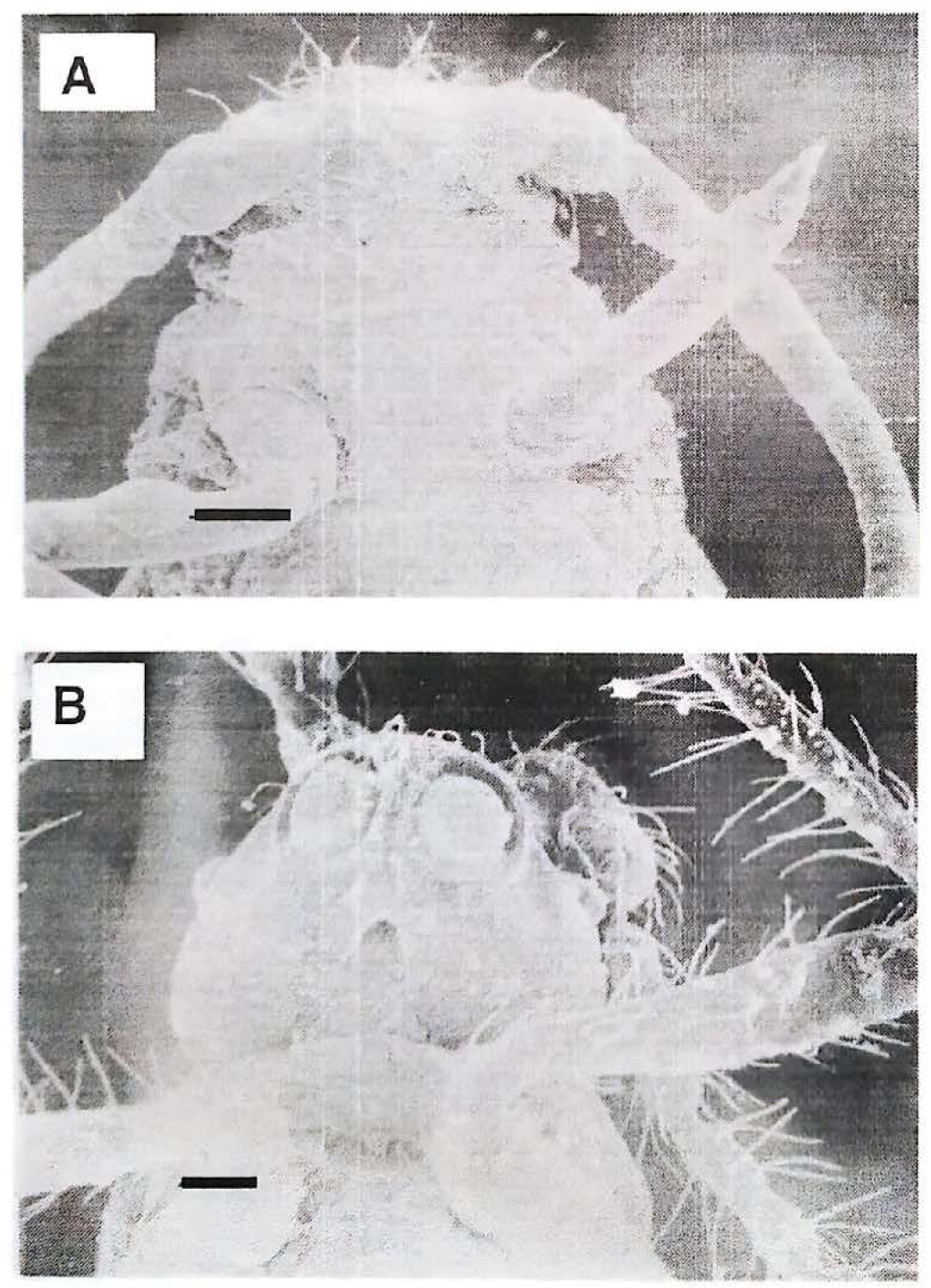
Micrografias en microscopía electrónica de barrido. A) Macho de cuarto instar y B) adulto. Las micrografias muestran que el macho en estos estadios de desarrollo presenta una ausencia del aparato bucal externo y en adulto, en su reemplazo, se observa una invaginación (raya = 50 µm).
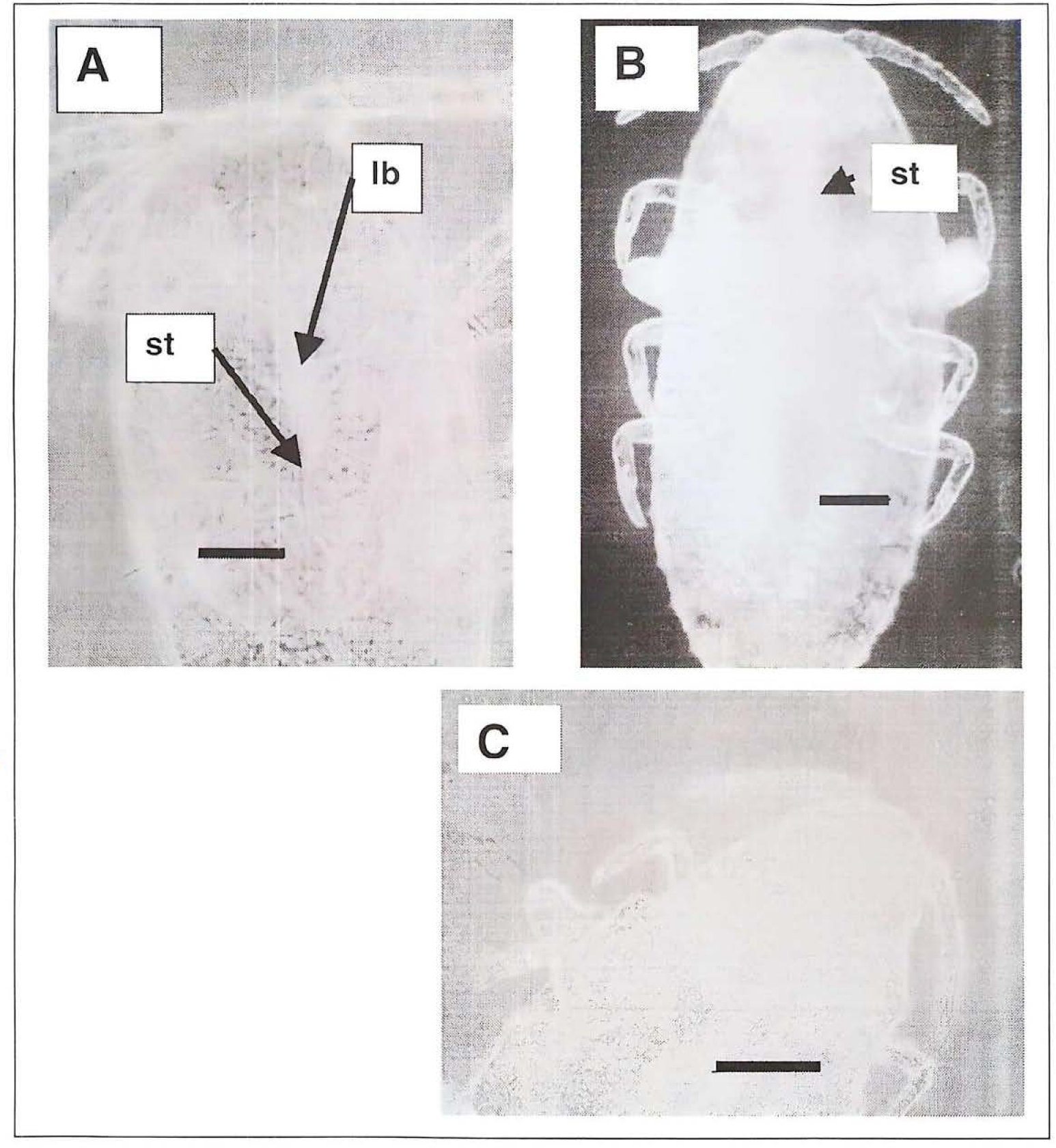
Micrografías en microscopia de contraste de fases. A) Macho de segundo instar en el que se observa por transparencia claramente el labium (lb) y los estiletes (st)(resultado similar con las hembras). B) Macho de tercer instar en el que se observan muy tenues los estiletes. C) Macho de tercer instar más avanzado en donde ya no se aprecian las piezas bucales (raya = 50 μm).
Comportamiento alimenticio de P. herreni por EPG.
Con base en las observaciones microscópicas, las actividades de alimentación de P. herreni se estudiaron con machos de segundo instar y con hembras de segundo, tercer instar, y adultas. Por medio de la técnica de EPG se distinguen básicamente 4 tipos de ondas diferentes, cada una característica de una actividad y localización precisa de los estiletes (Fig. 4). Las señales eléctricas básicas encontradas en áfidos (Tjallingii 1990) y en otra especie de piojo harinoso de la yuca Phenacoccus manihoti (Calatayud et al. 1994) han sido descritas como: np (no penetración), C (penetración extracelular), pd (caida de potencial: indicador de penetración intracelular pero no en el floema), y E (penetración intracelular en el floema con ingestión sostenida de savia). Cada vez que es registrada una caida de potencial se evidencia el paso de los estiletes a través de la membrana plasmática de una célula viva, pero esto hace dificil distinguir entre una célula del mesófilo y una del elemento criboso transportador de savia floemática. El criterio utilizado fue la duración de la señal y la aparición de miel durante la misma. En efecto, las caidas de potencial (pd) mayores de 8 min están relacionadas con la ingestión sostenida de savia floemática (Tjallingii 1990) y asociada generalmente con la producción de mielecilla. Las señales eléctricas obtenidas durante 8 h, se interpretaron identificándose los tipos de señales presentados utilizando el programa MacStylet (Febvay et al. 1996). En todos los EPG's, fueron identificadas las señales np, C, pd y E (Fig. 4); y se tuvo en cuenta la duración total de cada tipo de señal. Los registros realizados con machos y hembras de cada estadio de desarrollo estudiado, no presentaron diferencias significativas entre los porcentajes medios de permanencia de los estiletes en ninguna de las señales eléctricas, indicando que tienen el mismo comportamiento alimenticio y que se nutren principalmente de savia floemática (E representa el 50% del tiempo total de registro) (Tabla 1). Esta observación es apoyada por la presencia de aminoácidos en las gotas de miel excretadas por los insectos (resultado no presentado).
Porcentajes medios de permanencia de parámetros obtenidos en EPG sobre los registros realizados con machos de segundo ínstar y con hembras de segundo y tercer ínstar, y adultas de P, herreni.
C (penetración extracelular de los estiletes); pd (penetración intracelular de los estiletes pero no en el floema) y E (penetración intracelular de los estiletes en el floema con ingestión sostenida de savia).
Los valores de P se calcularon mediante tablas de contingencia (comparaciones de acuerdo con el estadio de desarrollo y el sexo.
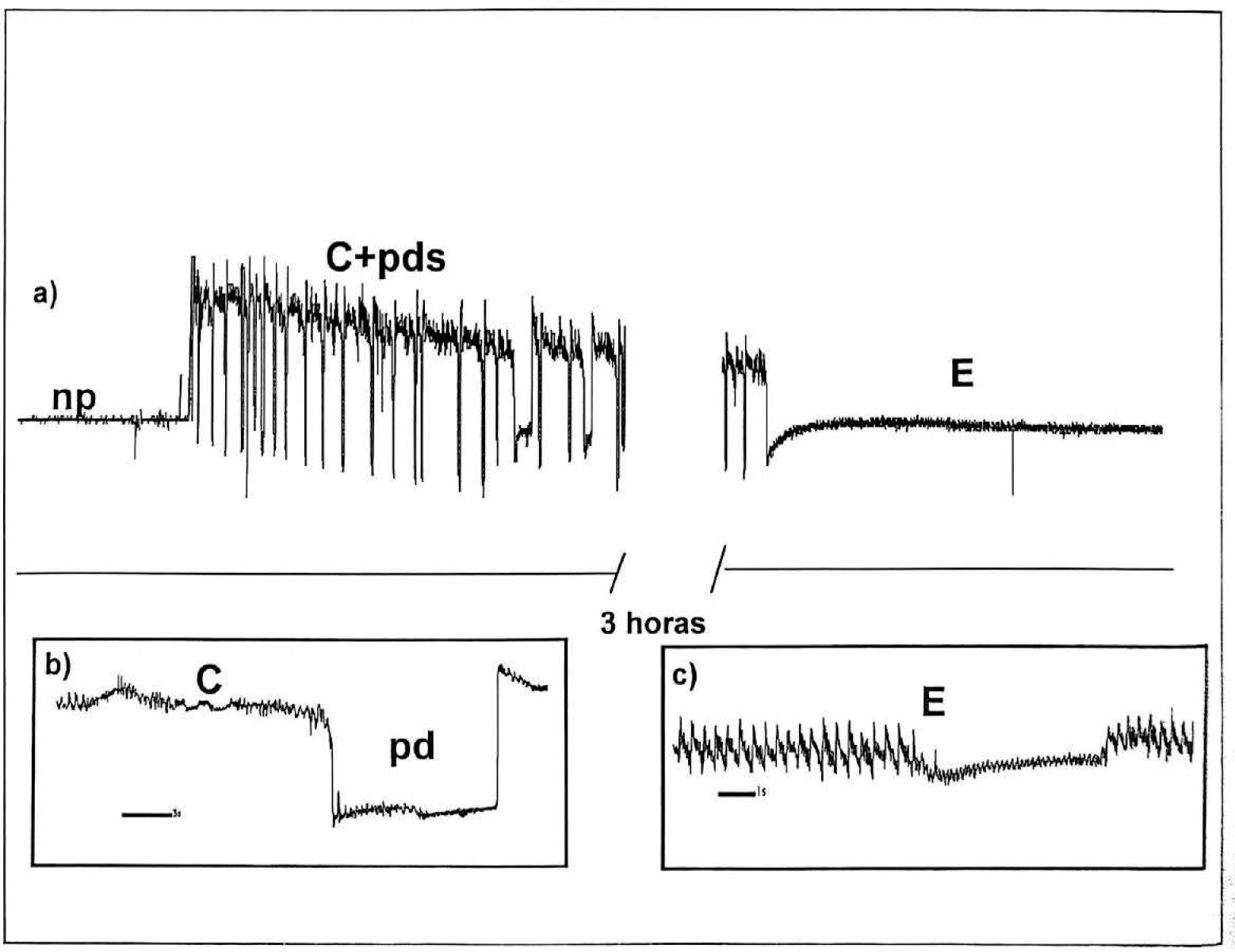
Patrones de EPG de P. herreni. a) Registro donde se observan las señales np (no hay penetración), C+pds (mezcla de penetración extracelular e intracelular) y E (posición intracelular con ingestión sostenida de savia del floema); b) Detalle de la señal C y de una caída de potencial (pd): c) Detalle de la señal E.
Este estudio sobre el comportamiento alimenticio de P. herreni, deja en claro que los insectos hembra se alimentan principalmente de savia floemática como las otras especies de Pseudococcidos estudiadas (Grassé 1951; Coquis y Salazar 1975; Calatayud et al. 1994). En cuanto a los machos de P. herreni es la primera vez que se realiza un trabajo sobre sus hábitos alimenticios desde segundo instar hasta el estado adulto. El insecto solamente se alimenta de forma continua de la planta (principalmente de savia floemática) durante sus dos primeros estadios de desarrollo (5-10 días), al cabo de los cuales sus piezas bucales empiezan a retraerse y cesa de alimentarse, confirmando que la principal función de los machos es su participación en el evento reproductivo (Varela y Bellotti 1981).
Por lo que, se sugiere que son las hembras de P. herreni, al conservar su aparato bucal y alimentarse de la savia del floema durante todo su ciclo de vida (Tabla 1), las que causan el mayor deterioro a los cultivos de yuca, ocasionando pérdidas hasta del 88% en rendimientos y del 74% en material de siembra (Herrera et al. 1987), por lo tanto los estudios siguientes se realizaron con hembras.
Efecto del déficit hídrico de plantas de yuca sobre el desarrollo de P. herreni.
Las plantas de yuca sometidas a estrés hídrico se evaluaron morfológica y fisiológicamente, para determinar la validez del sistema de riego aplicado. Un mes después de haber sometido las plantas a condiciones de riego diferencial, las plantas estresadas hídricamente mostraron una área foliar menor de 52 cmβ (vs 64 cmβ en plantas control) y una altura menor de 34 cm (vs 52 cm en plantas control), mientras que la resistencia estomática de las hojas fue más importante de 30 s/cm (vs 6 s/cm en plantas control). Estos cambios en plantas estresadas hídricamente que contribuyen a evitar la pérdida de agua (Cock et al. 1985; Yao y Goue 1992; El-Sharkawy 1993), indican que las plantas utilizadas en este estudio han iniciado un proceso de adaptación al déficit hídrico.
Al comparar el desarrollo de los insectos criados sobre plantas en déficit hídrico con plantas control, se presentaron diferencias significativas (Tabla 2). Las hembras desarrolladas en plantas con estrés hídrico aumentaron su fecundidad (F=6.2; df= 15; P<0.05), su tasa intrínseca de crecimiento natural (rm)(F=13.6; df= 15; P<0.01), su peso (F= 14.8; df= 142; P< 0.001). También se observó, que las hembras desarrolladas sobre plantas estresadas hídricamente disminuyeron su duración de desarrollo. La figura 5 muestra cómo las hembras desarrolladas sobre plantas en deficiencia hídrica forman el ovisaco (fase considerada como estado adulto maduro) más rápidamente que sobre plantas control. Mientras que en el día 22 de desarrollo el 60% de las hembras sobre plantas en estrés hídrico había empezado a formar el ovisaco, en plantas control, aproximadamente el mismo porcentaje de hembras adultas se obtuvo 2.5 días después.
Fecundidad, tasa intrínseca de incremento natural (rm) y peso de hembras de P. herreni (media* ± error estándar seguida del número de repeticiones en paréntesis) criadas sobre la variedad CMC-40 de yuca sometida a diferentes regímenes de riego. Tratamiento: NS = plantas control y S = plantas con estrés hídrico.
Valores seguidos de diferentes letras son significativamente diferentes al nivel del 5% de la prueba PLSD de Fisher (prueba posterior a la ANOVA); a, b: comparaciones por filas de acuerdo con el factor tratamiento (NS vs S).
En conclusión, las plantas con déficit hídrico favorecen el desarrollo de P. herreni (los rm's son mayores en las plantas con irrigación escasa). Este resultado puede explicar en parte el aumento de la población del insecto durante las estaciones secas (Bellotti et al. 1983; Noronha 1990) y coincide con los resultados obtenidos en laboratorio con otra especie de piojo, P. manihoti, que mostró rm's mayores en plantas con estrés hídrico (Fabres y Le Rü 1988). Los resultados obtenidos sugieren la existencia de algún tipo de interacción entre los compuestos sintetizados por la planta en déficit hídrico que favorecen, desde el punto de vista nutricional, a los insectos, traduciéndose en el aumento de su crecimiento y de su desarrollo.
El análisis en aminoácidos libres de hojas provenientes de plantas, bien y mal irrigadas, presentó algunas diferencias (Fig. 6). Las plantas bajo estrés hídrico mostraron un incremento significativo en el porcentaje de asparagina (xβ= 9.2, P<0.01). Por el contrario, el porcentaje del ácido glutámico disminuyó significativamente (xβ= 4.8, P<0.05). Estos resultados están de acuerdo con algunos de los mecanismos de regulación osmótica utilizados por las plantas para adaptarse a la sequía mencionados por Lamaze et al. (1996). De hecho, los tejidos de las plantas en condición de sequía, generalmente presentan acumulación de asparagina. Además, el incremento significativo (P<0.01) encontrado sobre plantas bajo estrés hídrico, se presentó en el aminoácido asparagina (Fig. 6). Con el fin de verificar si este aminoácido podría tener un papel indispensable para el desarrollo de P. herreni, se estudió la influencia de su ausencia mediante una dieta artificial. En comparación con la dieta control, los insectos alimentados con la dieta carente de asparagina tuvieron un desarrollo retardado (tan solo alcanzaron el tercer instar después de 35 días) y su crecimiento fue menor, presentando un peso significativamente menor (Tabla 3) (P<0.001 según la prueba U de Mann-Whitney). De acuerdo con los resultados obtenidos se puede concluir que la asparagina parece indispensable en el desarrollo de P. herreni. El análisis de la composición en aminoácidos libres de los insectos, mostró que los piojos desarrollados con dieta carente en asparagina presentaron una concentración similar al de los insectos desarrollados con dieta control (resultado no presentado). Al parecer el retraso en el crecimiento y desarrollo de los insectos criados sobre la dieta carente de asparagina, no es debido al hecho que los insectos no pueden sintetizar este aminoácido, puesto que en la composición aminoácidica de los insectos apareció asparagina, sino que se trata más de un problema energético. Es muy probable que el hecho de suplir la carencia de asparagina represente para el insecto un desgaste energético mayor, debido a que tiene que utilizar rutas de síntesis menos frecuentes.
Peso (media* ± error estándar seguida del número de repeticiones en paréntesis) y nivel de desarrollo alcanzado de hembras de P. herreni criadas durante 35 días con diferentes dietas (dieta control con todos los aminoácidos y una sin asparagina)
Valores seguidos de diferentes letras son significativamente diferentes al nivel del 5% de la prueba U de Mann-Whitney
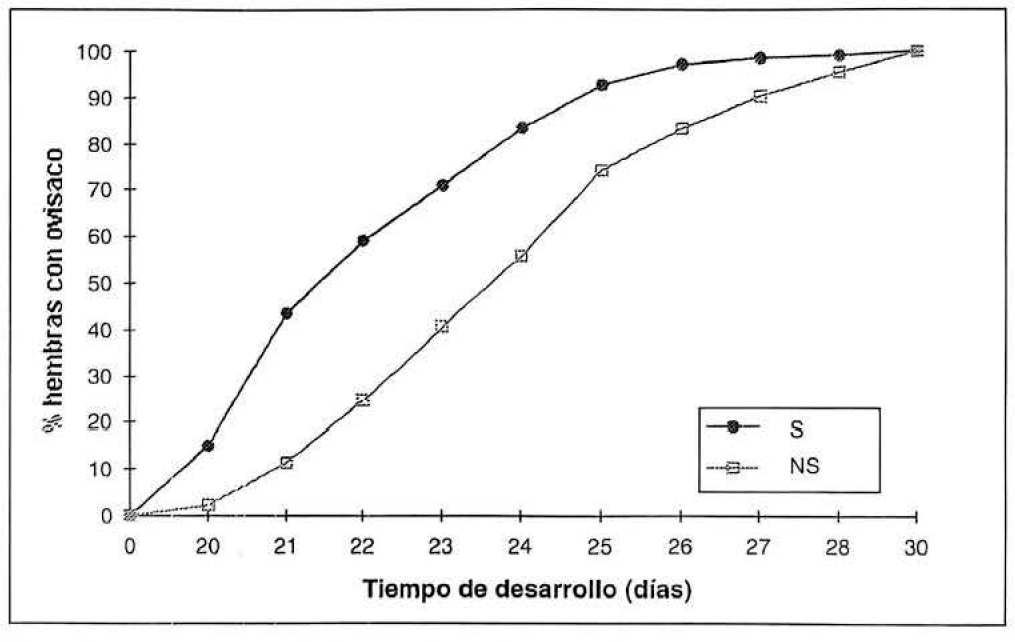
Porcentaje acumulado de hembras adultas maduras (i.e. que han empezado a formar el ovisaco) de acuerdo con el tiempo de desarrollo de P. herreni criado sobre plantas control (NS) y en déficit hídrico (S). Observaciones realizadas sobre 52 hembras para las plantas control y sobre 92 hembras para las plantas en déficit hídrico.
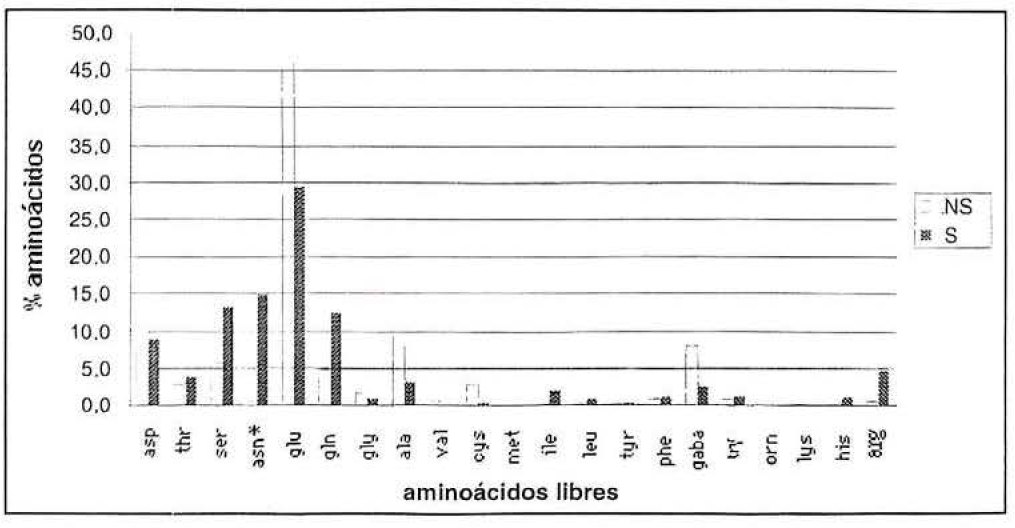
Composición en aminoácidos libres de hojas maduras infestadas por P. herreni y provenientes de plantas en condición normal de riego (NS) y bajo déficit hídrico (S). * Incrementó de manera más significativa en plantas bajo déficit hídrico mediante tabla de contingencia con P<0.01.
Conclusiones
Se confirmó que Phenacoccus herreni es un insecto principalmente floemófago. Los machos pierden la funcionabilidad de sus partes bucales en el tercer ínstar, sin embargo se desarrollan hasta el estado adulto, participando principalmente en la reproducción de la especie, por lo que las hembras, en conclusión, constituyen la plaga para la planta hospedera.
Se evidenció, en condiciones de laboratorio, que las plantas bajo déficit hídrico favorecen el desarrollo y crecimiento de las hembras de P. herreni. La composición aminoacídica de las hojas de yuca varía de acuerdo con la disponibilidad de agua del suelo: el déficit hídrico produce un incremento en el porcentaje de algunos aminoácidos, siendo el más importante asparagina.
Mediante la utilización de una dieta artificial se pudo concluir que este aminoácido es importante, y por ende incide directamente en el desarrollo de las hembras de P. herreni, por cuanto su ausencia se manifiesta en el retraso del desarrollo y reducción en el peso del insecto. Sin embargo, sería interesante reproducir la composición en aminoácidos obtenidas en plantas bajo déficit hídrico y probar artificialmente su influencia sobre el desarrollo de P. herreni.
Footnotes
Agradecimientos
Gracias a Guillermo Guzmán y José Alejandro Arroyave por su asistencia técnica en microscopía, a Josette Guillaud por su ayuda en los análisis de aminoácidos, y a Josefina Martinez, Ana Milena Caicedo y a Javier Hernández por sus lecturas críticas y sus sugerencias al manuscrito final. El presente estudio fue financiado por el ministerio de la educación francesa (Proyecto ICE 54, ORSTOM-CIAT).