
Abstract
The selection of a host is an important factor affecting the probability of survival of an herbivore. Its influence is greater on highly monophagous guests. Battus polydamas is a holigophagous herbivore on 24 species of Aristolochia, a toxic plant and frequent Troidini host. The large number of host species suggest that many species of Aristolochia are suitable as hosts and do not significantly affect survival probabilities of their guest. We examined two questions: Does B. polydamas exhibit a preference for host species? and What effect does the host have on butterfly development? We studied oviposition and herbivory by B. polydamas on eight species of Aristolochia. All species of Aristolochia were natural hosts, but there were significant differences between Aristolochia species with respect to the number of clusters laid. The butterflies preferred the apical zone for laying eggs. We measured host preference of eight species of Aristolochia by measuring leaf areas for those portions that had been consumed; leaves were administered simultaneously to different caterpillar groups (caterpillars for each group were from different clutches). Leaf nutritional quality was measured by the growth and development of caterpillar groups fed on a single specie of Aristolochia. We didn't observe significant differences between consumption rates of leaves, but the caterpillars attained the pupae stage on only three species of Aristolochia. Caterpillar development was faster in the host with lower sclerophile. We observed a close relation between sclerophile levels and oviposition and herbivory preferences, but that wasn't the only determining factor. Hosts don't offer an equal survival possibility. Female butterflies could use a suboptimal host when an ideal one wasn't present; then, they do deposit their eggs on flowers or young leaves. We concluded that female butterflies exhibit preferences for species of Aristolochia that provided higher survival probabilities.
Introducción
Los herbívoros se dividen, de acuerdo con la diversidad de hospederos que tengan, en generalistas, los que pueden comer plantas de grupos taxonómicamente poco relacionados, y especialistas, aquellos que consumen plantas de una especie -monófagos- o un grupo de especies estrechamente relacionadas -oligófagos-. Dentro de estos últimos se incluyen los géneros de la tribu Troidini (Lepidoptera: Papilionidae), cuyos principales hospederos son plantas de la familia Aristolochiaceae (Rothschild y Jordan 1906; Ehrlich y Raven 1964; Ackery 1984; Otero y Brown 1984; DeVries 1987; Brown et al. 1991).
Las Aristolochiaceae son bejucos con un alto y variado contenido de metabolitos secundarios, entre los cuales figuran alcaloides bencilisoquinolínicos, terpenoides, fenantrenolactonas y ácidos aristolóquicos, que actúan como defensas muy efectivas frente a la mayoría de los herbívoros (Chen y Zhu 1987: González 1991; Hegarty et al. 1991): no obstante, estas plantas están sometidas al ataque de especies de Troidini que, como Battus polydamas L., acumulan estos metabolitos secundarios y los utilizan como defensa ante sus propios depredadores, estableciendo tal grado de dependencia que estas mariposas no pueden consumir plantas de otro género (Urzúa y Priestap 1985; Brown et al. 1991). Battus polydamas es uno de los Troidini que presenta mayor número de especies de Aristolochia como hospedero (24, sensu Fagua y Ruiz 1993, 1996), al tiempo que es uno de los papiliónidos de mayor distribución en América (Rothschild y Jordan 1906; Young 1971, 1972; Scriber y Feeny 1979; Rausher 1979; Racheli y Pariset 1992; Tyler et al. 1994). Sin embargo, a excepción de lo realizado por Brown et al. (1981) y Miller y Feeny (1989), es poco lo que se conoce acerca del efecto que puedan tener los diferentes hospederos en el desarrollo y la supervivencia de las orugas de esta mariposa.
Al respecto, se sabe que la calidad del hospedero puede ser fundamental para la supervivencia de los herbívoros; elementos como nitrógeno, fósforo, azufre y potasio se encuentran en mayor proporción en las zonas de crecimiento de la planta (Medina 1977), y el nitrógeno disponible es determinante en la tasa de crecimiento y en los niveles de reproducción de poblaciones de insectos (Feeny 1975; Scriber y Feeny 1979; McNeil y Southwood 1978, citado por Harborne 1982). Loader y Damman (1991) encontraron, además, que una baja calidad nutricional de la hoja reduce la sobrevivencia de herbívoros de manera indirecta al incrementar el período de exposición a enemigos naturales. Por lo tanto, es vital para un herbívoro escoger el hospedero más adecuado, siendo la oviposición uno de los mayores focos de selección al incrementar la probabilidad de que la oruga alcance un recurso alimenticio en el que se pueda desarrollar (Chew 1977; Rausher 1979). La selección del recurso se inicia con el proceso mismo de la postura, las hembras necesitan ubicar de manera visual un hospedero, generalmente disperso, para luego seleccionar, mediante percepción química, las partes más aprovechables (Brues 1924; Rausher 1978, 1979, 1980; Feeny et al. 1983; Papaj y Rausher 1983); se reconocen variaciones individuales en cuanto a preferencias de oviposición debido a que las hembras de una especie pueden aceptar substratos subóptimos cuando el óptimo es raro o no se encuentra (Berembaum 1981; Singer 1984).
De acuerdo con lo anterior, la diversidad de hospederos de Battus polydamas indicaría que la especie de aristoloquia utilizada no afecta en gran medida las probabilidades de supervivencia del insecto; se plantearon entonces dos preguntas: ¿Presenta B. polydamas alguna preferencia por determinadas especies hospedero? y ¿Qué efecto tiene el hospedero en el ciclo de desarrollo del insecto? Con base en estas preguntas se planteó como hipótesis que, dado el gran número de hospederos de B. polydamas, que utiliza todas o la mayoría de las especies de Aristolochia presentes en una localidad, no deben existir grandes diferencias entre los hospederos utilizados en cuanto a número de inmaduros observados en especies coocurrentes en campo, preferencia de las orugas por una u otra especie ofrecida como alimento y los tiempos de desarrollo y las tasas de supervivencia de las orugas al ser alimentadas con solo una especie de hospedero. En consecuencia, los objetivos fueron: observar en campo el comportamiento de oviposición y herbivoría de la mariposa y determinar, mediante pruebas de laboratorio, las preferencias de las orugas y el efecto que tienen los hospederos sobre su ciclo de desarrollo.
Materiales y Métodos
Los estados inmaduros y las plantas hospederas se obtuvieron en sitios aledaños a los municipios de La Dorada y Victoria (Departamento de Caldas, Colombia), Mariquita (Depto. del Tolima), y Restrepo, Villavicencio, Puerto López, Granada y Fuente de Oro (Depto. del Meta) (Fig. 1). La temperatura media de los sitios está entre los 27° C y 29°C y tienen un régimen de lluvias bimodal con tendencia monomodal. Todos los sitios presentaron bosques ribereños con escasas plantas trepadoras y frecuentes arbustos y herbáceas importantes como fuentes de néctar para mariposas adultas. En cada sitio de estudio se observaron cuatro o más plantas de por lo menos una especie de Aristolochia.
Los sitios de estudio se visitaron entre enero y septiembre de 1992. Se buscaron inmaduros de lepidópteros en los vástagos de Aristolochia y en la vegetación aledaña; las posturas y orugas colectadas se depositaron, junto con un explante de la planta sobre la cual se encontraron, en vasos plásticos desechables (de 10 onzas), cubiertos con una película de papel-vinilo. Se anotaron: ubicación en la planta, número de individuos, estado de desarrollo y coloración, especie del hospedero y órgano en el que se hizo la colección. Para la ubicación se definieron: posición apical (la mitad distal del vástago) y posición basal (la mitad proximal del vástago, incluida la base de la planta).
Se compararon la forma y el tamaño foliar de cada Aristolochia, el hábitat de la planta y el número de huevos encontrados para detectar posibles relaciones entre estos factores y preferencias de oviposición. También se tomaron 30 hojas en diferente estado de desarrollo de cada especie de hospedero, a las cuales se les observó el tipo de indumento y se les midió el área foliar y el peso en fresco y en seco para posteriormente aplicar las fórmulas propuestas por Veneklass (1985) para medir la suculencia y el porcentaje de esclerofilia, como valores indicativos de la consistencia foliar.
Material fresco de Aristolochia se colectó y mantuvo en refrigerador para ser utilizado como alimento para las orugas, aunque se procuró emplear material obtenido de plantas extraídas de la zona de estudio y cultivadas en invernadero para las pruebas de laboratorio.
Una vez en laboratorio, los inmaduros se colocaron dentro de una cámara de cría a temperatura promedio de 26° C y humedad relativa del 75%. Todo el material obtenido se encuentra depositado en la Colección de Entomología del Instituto de Ciencias Naturales (ICN-MHN) de la Universidad Nacional de Colombia.
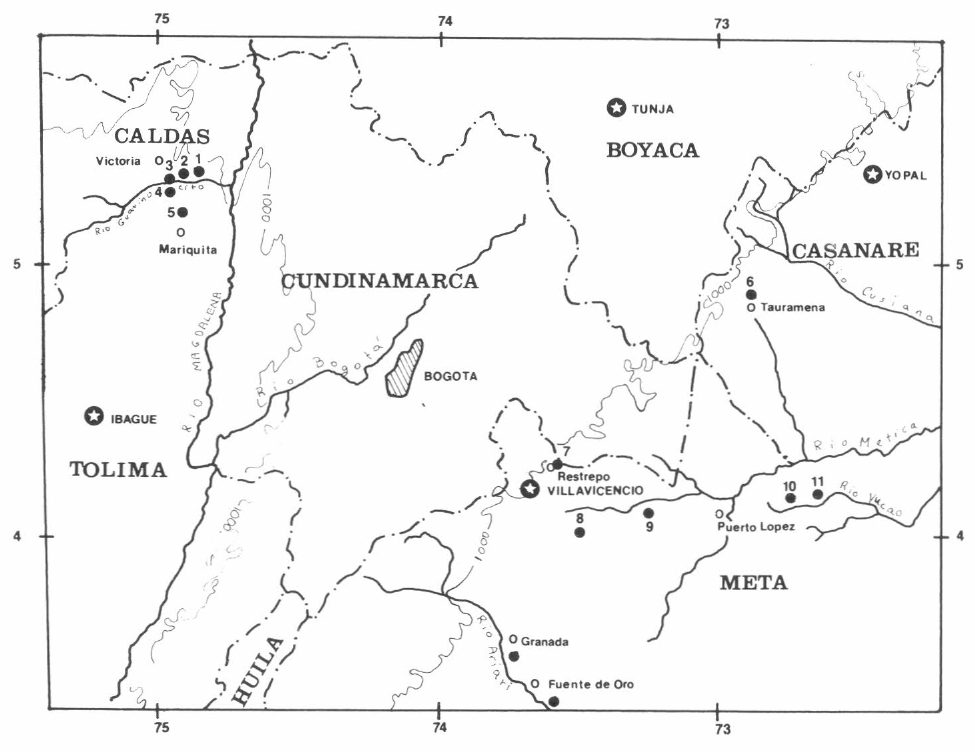
Mapa del área de trabajo. Se señalan los sitios de estudio. 1, 2, 3, sitios aledaños al Valle del río Guarinó. 4, 5, Río Medina - Mariquita. 6, Tauramena. 7, Restrepo. 8, 9, Villavicencio. 10, 11, Puerto López. 12, Granada. 13, Fuente de Oro.
Se realizaron dos pruebas de laboratorio para estudiar los hábitos alimentarios de las orugas; así:
Resultados
Se encontraron ocho especies de Aristolochia en los sitios de estudio: Aristolochia acutifolia Duchartre, A. maxima Jacq. (pertenecientes a la Serie Thyrsicae), A. morae González, A. odoratissima L., A. pilosa H.B.K., A. ringens Vahl (Serie Hexandrae, subserie Hexandrae), A. cordiflora Mutis ex H.B.K. y A. leuconeura Linden (Serie Hexandrae, subserie Anthocaulicae; sensu Pfeifer 1966 y González 1990, 1991). En la mayoría de los sitios de muestreo se encontró más de una especie y su abundancia fue variable (Fig. 2a); la más frecuente fue A. pilosa, seguida de A. acutifolia, A. leuconeura y A. ringens. En cuanto a hábitat, A. cordiflora y A. leuconeura se encuentran predominantemente hacia el interior del bosque, A. acutifolia y A. maxima predominan en bordes de bosque, A. odoratissima predomina en zonas intervenidas pero con sombrío y A. pilosa, A. morae y A. ringens son más comunes en espacios abiertos muy intervenidos.
Las especies que presentaron mayor valor de suculencia fueron Aristolochia pilosa y A. ringens, mientras que A. acutifolia y A. maxima mostraron el menor valor; por el contrario, el porcentaje de esclerofilia fue mayor en A. maxima, A. cordiflora y A. acutifolia y menor en A. odoratissima y A. pilosa (Fig. 2b).
Las características del tipo de indumento dividen las aristoloquias en dos grupos, en el primero están las especies de haz foliar, tallo, pecíolo y perigonio glabros (A. morae, A. ringens, A. cordiflora y A. leuconeura); en el segundo están las restantes que, a excepción de A. odoratissima, son especies con indumento piloso.
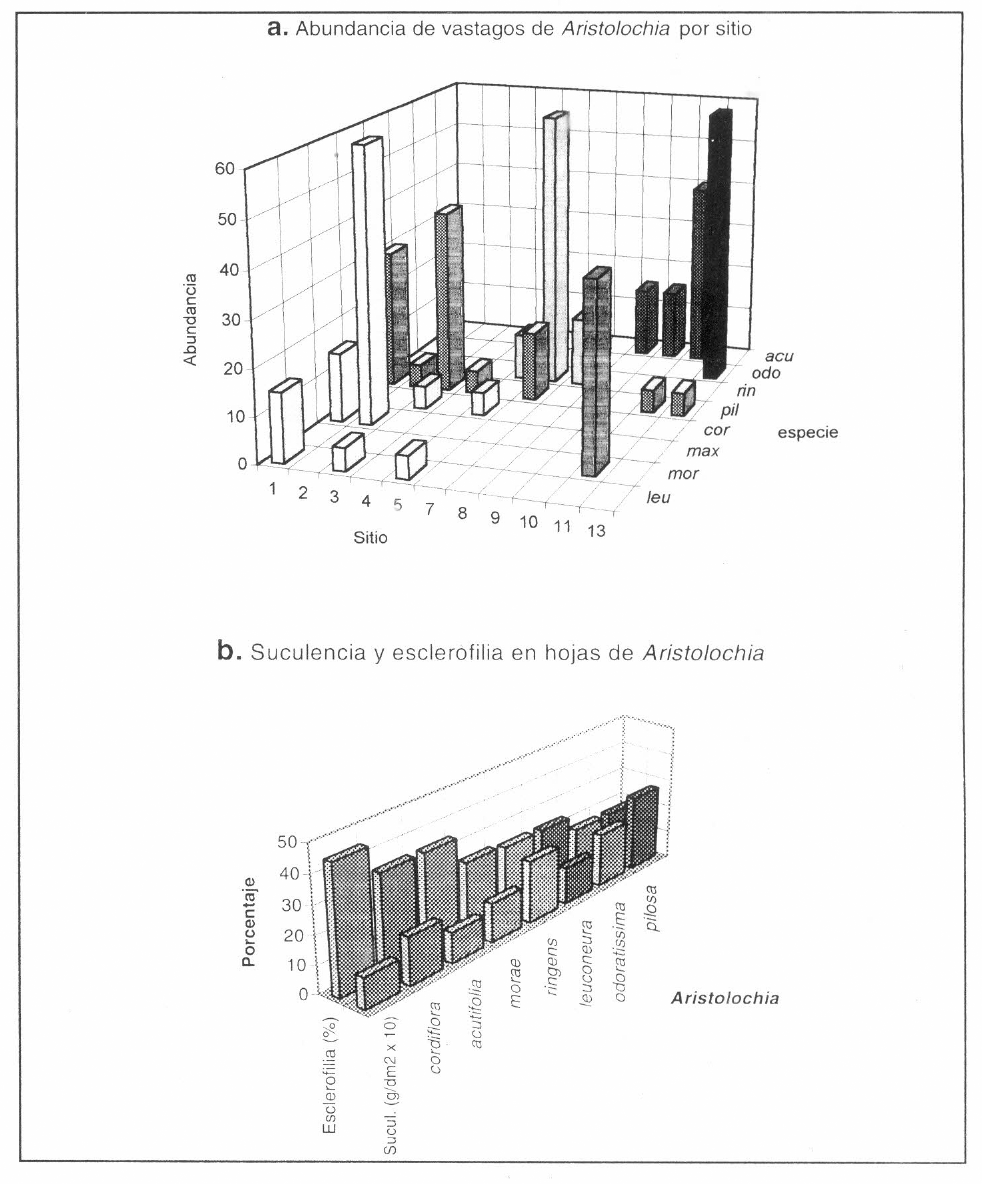
a: Abundancia cualitativa de las especies de Aristolochia en cada sitio de estudio. acu: A. acutifolia. odo: A. odoratissima. pil: A. pilosa. rin: A. ringens. cor: A. cordiflora. max: A. maxima. mor: A. morae. leu: A. leuconeura. b: Suculencia y porcentaje de esclerofilia de hojas de Aristolochia spp.; el valor de suculencia ha sido multiplicado por 10 para permitir su diagramación simultanea.
En cuanto a las mariposas, se encontró que las posturas de Battus polydamas tienen generalmente varios huevos de color amarillo (X= 5.51, s=2.40, n= 76; rango 1 a 16), bastante contrastante con el color de la vegetación y solo en dos casos (de 76 estudiados) se pudo observar más de una postura por vástago o individuo de Aristolochia. No hay diferencias significativas en el número de huevos por postura de acuerdo con la ubicación basal o apical en el vástago (p=0.86, prueba-t), ni se observaron diferencias en el número de huevos dependientes de la abundancia del recurso. Aristolochia pilosa presentó el mayor número de posturas, resultado debido probablemente a la mayor frecuencia y abundancia de la especie en los sitios de estudio (Figs. 2a y 3a). En general, el mayor número de huevos y posturas por sitio correspondió al hospedero más abundante, siendo generalmente menores los registros en las especies escasas coocurrentes. La excepción fueron A. ringens y A. morae que, pese a ser las más abundantes en algunos sitios, tuvieron siempre un número bajo de registros (Figs. 2a y 3a).
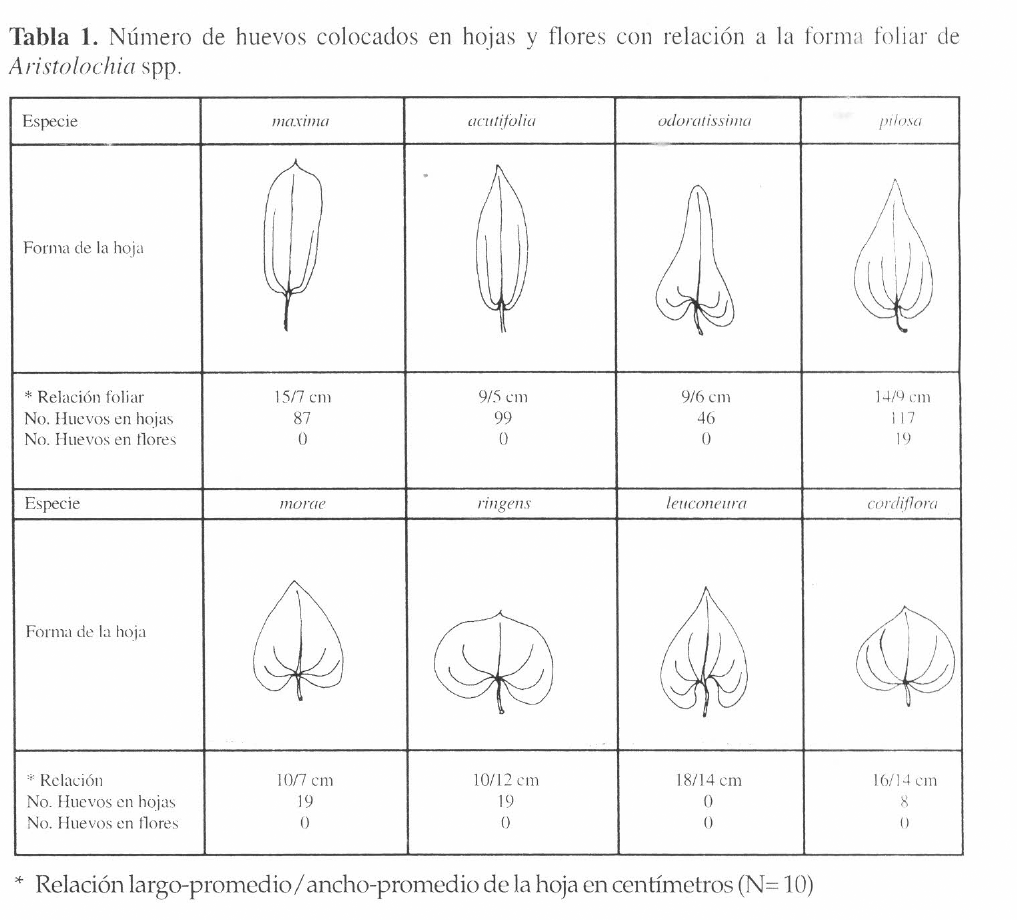
Se observaron más posturas en la parte apical de la planta, siendo mayor el número de registros en el tallo cuando éstos se encontraban en la mitad apical, mientras que en la mitad basal el sitio de oviposición predominante fue el pecíolo (Fig. 3b). Aristolochia pilosa fue el único hospedero en donde se encontraron huevos en todas las partes del vástago. La comparación de la forma y el tamaño foliar de cada Aristolochia con el número de huevos muestra mayor cantidad de registros en hospederos con hojas que tienden a ser alargadas y que se encuentran en el borde del bosque o en ambientes alterados con sombrío (Tabla 1).
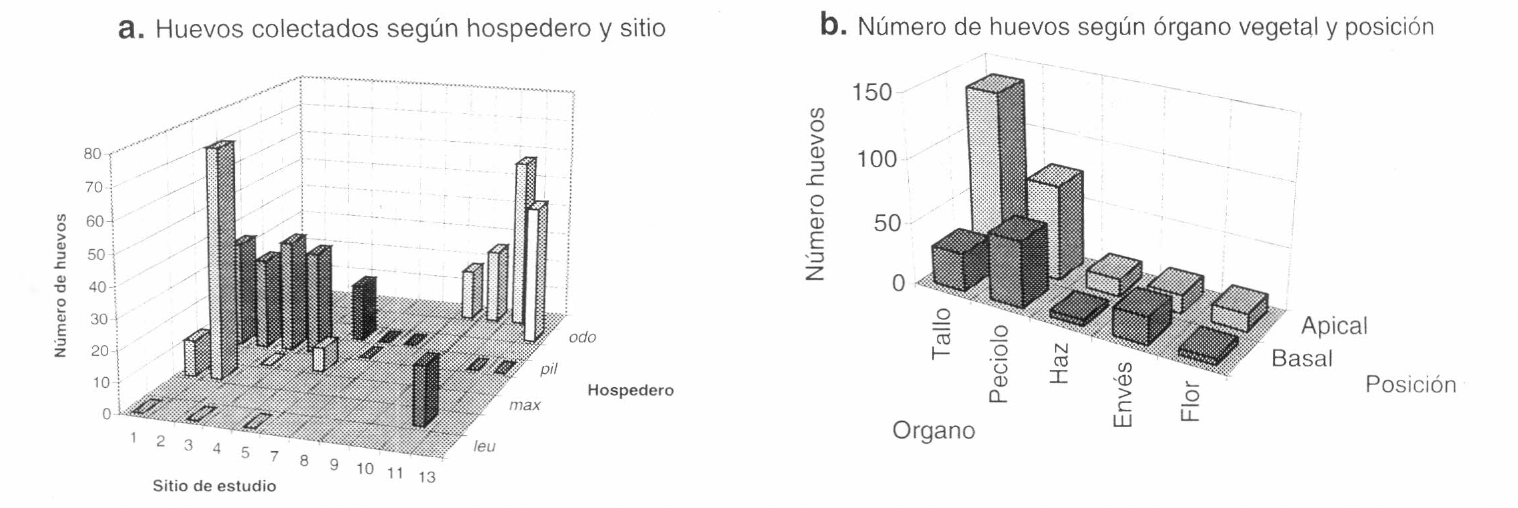
a: Número de huevos colectados de Battus polydamas en los hospederos de cada sitio de estudio. acu: A. acutifolia. odo: A. odoratissima. pil: A. pilosa. rin: A. ringens. cor: A. cordiflora. max: A. maxima. mor: A. morae. leu: A. leuconeura. b: Número de huevos de Battus polydamas colectados en los diferentes órganos vegetales y su posición dentro de la planta.
Las orugas son generalmente gregarias y de diferente coloración, mostrando gran variabilidad (Fig. 4a). El número de orugas observado es considerablemente menor que el de huevos (Figs. 3a y 4a), pero se mantiene el patrón de mayor número de orugas en el hospedero más abundante y el bajo número de registros en A. morae y A. ringens, pese a ser plantas abundantes. Un aspecto interesante es que el mayor número de orugas se detectó en A. leuconeura, un hospedero en donde no se hallaron posturas (Figs. 3a y 4a). En cuanto a posición, se observó un número mayor de orugas en la parte basal del vástago, lo cual podría estar relacionado con un incremento en la capacidad de asimilación de tejido esclerófilo de las orugas de últimos estadios, hecho observado posteriormente en animales criados en laboratorio; también se presentó una marcada preferencia por el envés foliar (Fig. 4b).
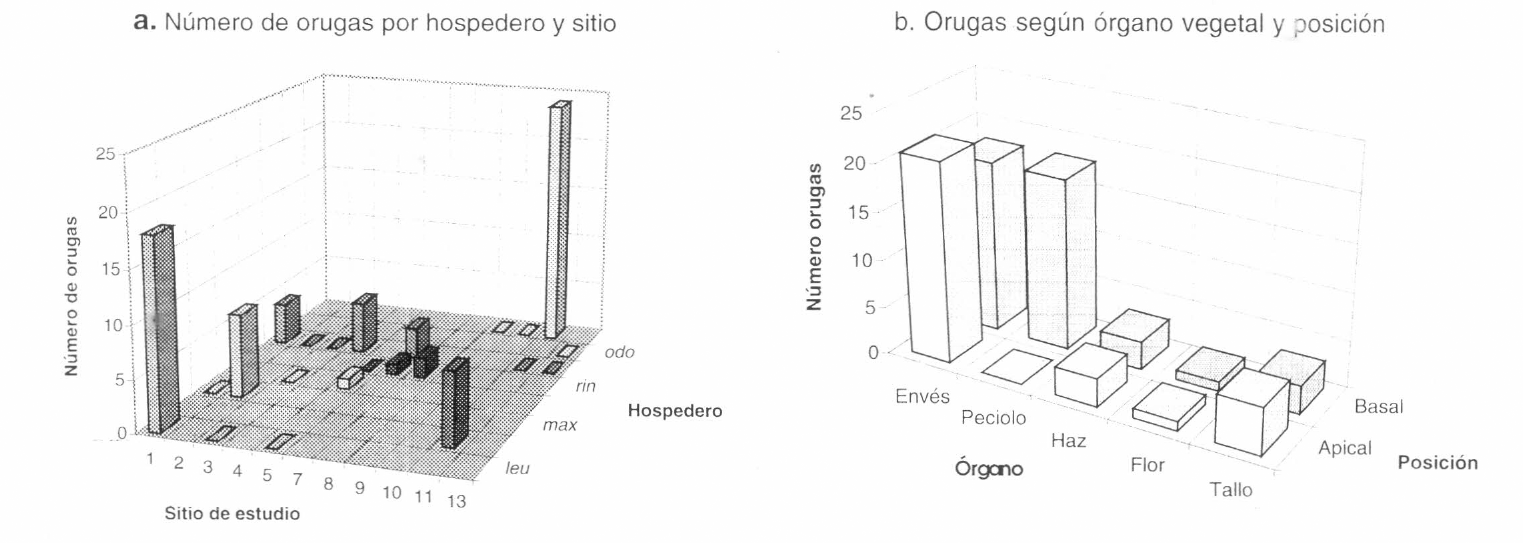
a: Número de orugas de Battus polydamas colectadas en los hospederos de cada sitio de estudio. acu: A. acutifolia. odo: A. odoratissima. pil: A. pilosa. rin: A. ringens. cor: A. cordiflora. max: A. maxima. mor: A. morae. leu: A. leuconeura. b: Número de orugas de Battus polydamas colectadas de acuerdo a su posición basal o apical y órgano del hospedero.
Battus polydamas, a pesar de ser un herbívoro con un amplio número de hospederos, fue el huesped que registró mayor número de inmaduros en Aristolochia pilosa, A. maxima y A. acutifolia, en comparación con el número total de registros de las especies consideradas huéspedes especialistas de aristoloquias encontradas en este estudio (solo se registraron otros tres grupos frecuentes: Parides spp., Mapeta xanthomella Walker y Battus spp., Fagua y Ruiz 1996; Fig. 5a). En B. polydamas el tiempo medio desde la eclosión de la oruga hasta la transformación a adulto es de 42.2 días; se presenta un aumento gradual en la duración de los 5 estadíos, en los cuales permanece en promedio 20.66 días; el tiempo en estado de oruga es bastante corto, representa solo la mitad del tiempo total de desarrollo a adulto. El incremento en la longitud de la oruga es mayor en las mudas 1º y 3º (Tabla 2). En crisálida dura en promedio 21.66 días. Los adultos llegaron a vivir hasta 6 días bajo condiciones de laboratorio.
En cuanto a las pruebas de laboratorio, se tuvo que el análisis estadístico de la prueba de preferencia no mostró diferencias significativas en el área foliar consumida de las Aristolochia en los estadíos analizados en conjunto (ANDEVA a dos vías, p=0.65, F=0.72; p=0.99, F=0.32), ni independientemente (ANDEVAs a una vía, p > 0.13 a 0.78 en todos los estadios). No obstante, vale la pena destacar que las orugas en primer estadío no comieron hojas de A. acutifolia, ni de A. maxima, y que, pese a variar el consumo de cada especie de un estadio a otro, siempre hubo un consumo ligeramente mayor de A. odoratissima a partir de la primera muda, mientras que A. morae y A. ringens siempre fueron poco o no consumidas (Fig. 5b). Como anotación se destaca que el área foliar consumida por un individuo, durante sus cinco estadíos, fue en promedio 310.48 cmβ (s= 10.3467; n=10).
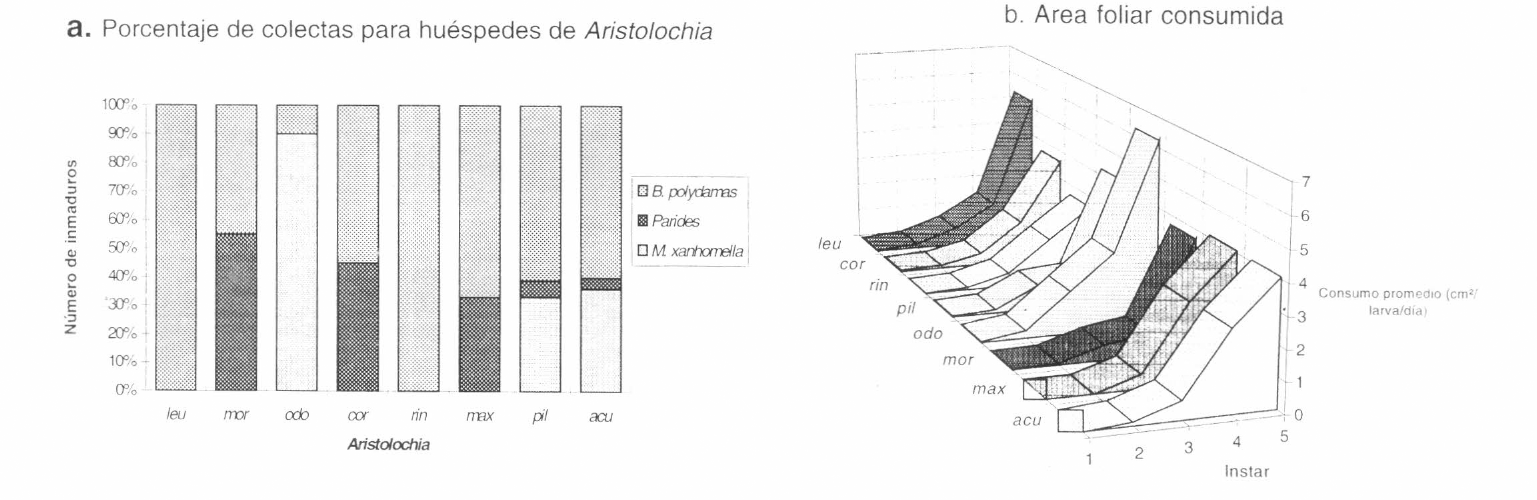
a: Porcentaje de uso de los hospederos por parte de los huéspedes registrados en campo; los datos incluyen número total de huevos y orugas. b: Consumo promedio de las especies de Aristolochia ofrecidas a orugas de Battus polydamas en la prueba de preferencia. acu: A. acutifolia. odo: A. odoratissima. pil: A. pilosa. rin: A. ringens. cor: A. cordiflora. max: A. maxima. mor: A. morae. leu: A. leuconeura.
En la prueba de calidad nutricional se encontró que la diferencia en el tiempo de permanencia en cada estadío larvario fue altamente significativa según el hospedero, al ser analizados todos los estadios en conjunto (ANDEVA a dos vías, P<0.001, F= 302.13; p<0.001, F=2,78; Tabla 3), y para los primeros cuatro estadios por separado (ANDEVA a una vía, p<0.001; en el 5° estadio p=0.15; Tabla 4). La duración en los primeros cuatro estadíos fue significativamente mayor en las orugas alimentadas con Aristolochia morae y A. ringens y menor en las criadas con A. odoratissima (Fig. 6a). En el quinto estadío las orugas en A. odoratissima presentaron menor tiempo de duración, aunque éste no fue significativamente diferente al de las alimentadas con A. pilosa u otras especies (Fig. 6a).
Número de huevos colocados en hojas y flores con relación a la forma foliar de Aristolochia spp.
Dimensiones promedio de la cápsula cefálica, longitud de la oruga al comienzo y final de la muda, y duración del período de desarrollo en Battus polydamas; la duración promedio en crisálida fue de 21.66 días. Datos obtenidos de 18 orugas críadas en dieta múltiple
Prueba de calidad nutricional; ANDEVA a dos vías del tiempo de duración de cada estadío larval de Battus polydamas en cada especie de Aristolochia suministrada como dieta (población de partida n=205; conformada por orugas recién eclosionadas)
V. explicada: variación explicada en el ANDEVA.
Prueba de calidad nutricional; ANDEVAs a una vía del tiempo de duración en cada estadío de orugas de Battus polydamas alimentadas con diferentes especies de Aristolochia
Prueba de calidad nutricional. Población inicial, número de orugas muertas en cada estadío, la fracción acumulada correspondiente y la mortalidad total en cada una de las dietas suministradas
P: Población inicial.
m..: Número de orugas muertas en cada estadío.
FA: Fracción acumulada.
En cuanto a la supervivencia, el análisis del número total de orugas muertas y sobrevivientes evidencia diferencias significativas entre las dietas (Xβ= 82.654; g.l.= 7; p<0.001; Tabla 5; Fig. 6b); los resultados que más se alejan de un comportamiento homogéneo son los de Aristolochia odoratissima y A. pilosa; aunque solo la mortalidad en A. odoratissima y A. acutifolia fue significativamente diferente de las demás dietas (Prueba de Kolmogorov-Smirnov; Tabla 6; Fig. 6b). Solo las orugas que consumieron odoratissima mostraron un comportamiento estable a lo largo de su período de desarrollo, en A. acutifolia y A. pilosa el número de sobrevivientes se reduce drásticamente en el cuarto estadío; las dietas restantes presentan una disminución gradual (Fig. 6b). La mortalidad menor se presentó en orugas alimentadas con A. odoratissima, seguidas de las criadas en A. pilosa, A. maxima y A. acutifolia; las orugas sólo llegaron hasta tercer estadío en Aristolochia cordiflora y a cuarto en A. morae, A. ringens y A. leuconeura; mientras que las alimentadas con A. acutifolia y A. maxima llegaron hasta crisálida, siendo las mantenidas en A. odoratissima y A. pilosa las únicas que alcanzaron el estado adulto (Fig. 6b).
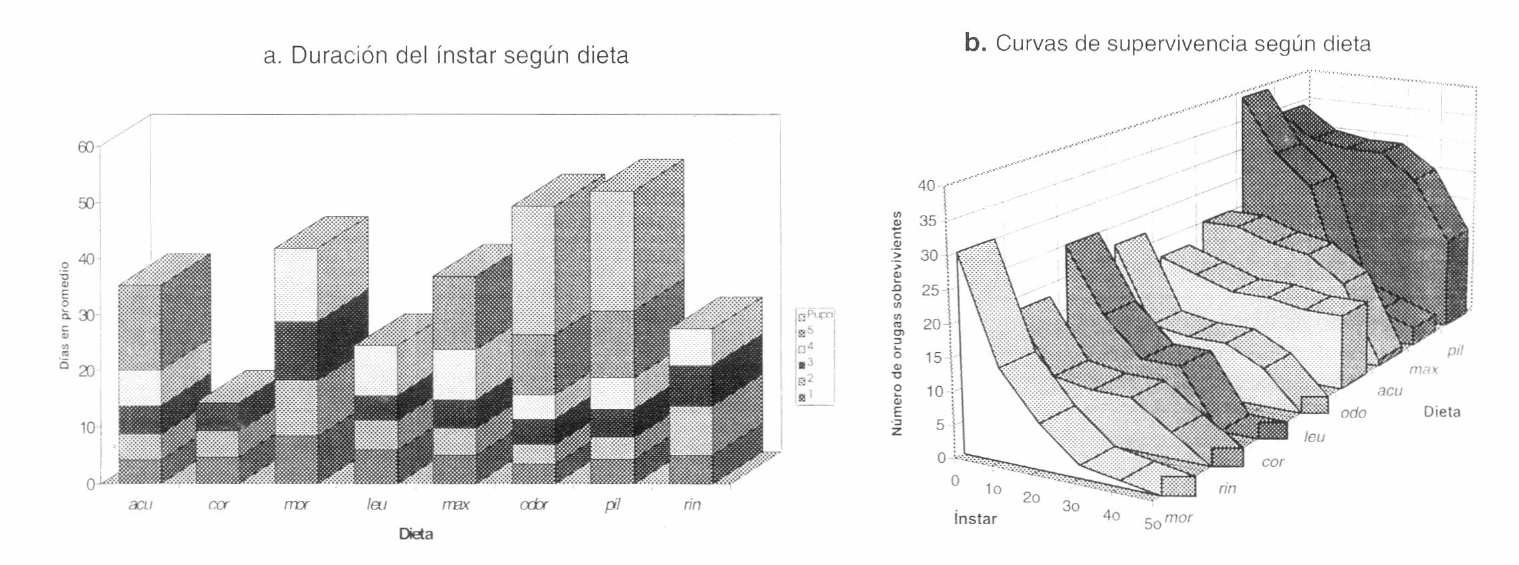
a: Duración promedio en días de cada ínstar en orugas de Battus polydamas criadas únicamente con una especie de Aristolochia en la prueba de calidad nutricional. acu: A. acutifolia. odo: A. odoratissima. pil: A. pilosa. rin: A. ringens. cor: A. cordiflora. max: A. maxima. mor: A. morae. leu: A. leuconeura. b. Número de orugas de Battus polydamas sobrevivientes en cada ínstar según la dieta suministrada en la prueba de calidad nutricional. acu: A. acutifolia. odo: A. odoratissima. pil: A. pilosa. rin: A. ringens. cor: A. cordiflora. max: A. maxima. mor: A. morae. leu: A. leuconeura.
Durante el presente trabajo se observaron cinco especies de parásitos de las familias Braconidae (2) e Ischneumonidae (3) (Hymenoptera). Aproximadamente el 14% de los huevos de Battus polydamas (55 de 428), además de tres orugas (11% de 27 colectadas en campo), resultaron parasitadas.
A manera de análisis se puede decir que el color conspicuo de los huevos de Battus polydamas y su ubicación en tallos y pecíolos de la mitad apical, donde son bastante notorias, puede ser evidencia de un mecanismo de inhibición de oviposición similar al registrado en otras especies de mariposas, el cual generaría la distribución de una postura por vástago de Aristolochia observada. Por otra parte, la distribución aproximadamente normal de las posturas permite decir que, para este caso, el número de huevos depositados no depende de la abundancia del recurso sino que puede asociarse mejor a un comportamiento definido de la hembra, la cual coloca generalmente entre cuatro y seis huevos por postura.
Prueba de calidad nutricional; prueba de Kolmogorov-Smirnov de la mortalidad de orugas de Batus polydamas en cada dieta
dmax: diferencia maxima.
dmin: diferencia mínima significativa con 95% de confianza.
Diferencia significativa (cuando dmax > dmin).
Aunque no muy estrecha, se presentó una relación entre las especies que Battus polydamas prefirió como hospederos en el campo y los resultados de la prueba de calidad; la capacidad de esta mariposa para consumir varias plantas del género quedó manifiesta en la baja selectividad observada en esta prueba, capacidad que aumenta con el desarrollo de las orugas. Por el contrario, se observó una alta coincidencia entre las especies preferidas en el campo y aquellas que permitieron a las orugas alcanzar el mayor desarrollo en el menor tiempo. Vale la pena señalar que los hospederos más utilizados en el campo son plantas con hojas más largas que anchas; esta característica podría desempeñar un papel importante en la identificación visual del hospedero y es un tema que merece una mayor profundización. Es importante recalcar que cuando ocurrieron dos o más hospederos con potenciales de supervivencia similares para las orugas, el hospedero más utilizado por las hembras fue el más abundante; por el contrario, si el hospedero más frecuente brindaba pocas posibilidades de supervivencia éste era, generalmente, poco utilizado, presentando gran número de posturas únicamente cuando los hospederos más óptimos no se encontraban.
El tiempo menor en el período de desarrollo de Battus polydamas en Aristolochia odoratissima y A. pilosa puede ser evidencia de una mayor calidad nutricional de los tejidos de sus hojas, probablemente asociada a la baja esclerofilia y mayor suculencia. El uso alto de A. pilosa no evidencia que el indumento sea un mecanismo de defensa muy efectivo, probablemente debido a la baja resistencia mecánica de los tricomas, al tamaño relativamente grande de las orugas y a su comportamiento gregario de alimentación. El número bajo de registros en A. odoratissima podría estar relacionado con el hábitat de esta planta, que si bien es la de mejor potencial de supervivencia, prefiere el interior del bosque perturbado o cultivos con bastante sombrío, ambientes poco frecuentados por B. polydamas.
Los datos de crecimiento lento y mortalidad alta obtenidos en Aristolochia acutifolia y A. maxima no concuerdan con el amplio registro de huevos y orugas; esto podría relacionarse con los valores altos de consistencia y el indumento corto pero correoso en las dos plantas, que permite el desarrollo de las orugas, pero quizás a costa de un incremento en el período de desarrollo y la mortalidad. Es posible que la preferencia observada por sitios de oviposición en lugares apicales, donde las orugas de primeros estadíos tienen acceso continuo a las hojas tiernas, disminuya esta presión y les permita desarrollarse normalmente; en favor de ésto cabe recordar que, en la prueba de preferencia, el área foliar consumida de estas especies se incrementa a medida que las orugas adquieren mayor tamaño y por ende mayor capacidad mecánica para cortar tejidos.
Contrario a las especies que permitieron el desarrollo, los hospederos de potencial más bajo coincidieron en tener una forma foliar aproximadamente circular. Un aspecto interesante es que dos de los hospederos de menor potencial (Aristolochia morae y A. ringens) no poseen indumento desarrollado, son glabras, muy suculentas y siempre fueron poco consumidas, incluso por orugas de último estado, lo que podría asociarse con la existencia de mecanismos de defensa bioquímicos diferentes a los presentes en los hospederos más utilizados. En cuanto a A. cordiflora, la tercera especie con bajo potencial de supervivencia, éste pudo estar asociado con su porcentaje alto de esclerofilia, producto de la esclerotización de las venas de 2º a 5º órdenes de la planta (cf. González 1989), los cuales actúan como barrera mecánica al fraccionar en pequeñas areolas, a manera de retículo, el área de tejido blando. Por otra parte, el período de desarrollo prolongado y la mortalidad alta de las orugas en A. leuconeura, puede relacionarse con la producción de un exudado amarillento que podría tener una función defensiva similar a la atribuida al látex de otros grupos vegetales (González 1989).
Teniendo en cuenta las preferencias reconocidas de Battus polydamas por sitios de oviposición próximos al ápice de los vástagos, el amplio rango de hospederos registrados, y la preferencia y mejor desarrollo observado en hospederos suculentos y poco esclerófilos, es probable que la susceptibilidad de un hospedero identificado (calidad de una planta para ser atacada por un fitófago) esté determinada principalmente por la cantidad de tejido joven o por el grado de esclerofilia que presenten sus órganos, además de las característica químicas de las plantas, que pueden estar asociadas a una forma foliar definida.
A manera de resumen, se puede decir que los registros de orugas y posturas de Battus polydamas en todas las especies de Aristolochia confirman el comportamiento oligófago de «generalista intragenérico» que tiene esta mariposa. No obstante, el uso de los hospederos no es indiscriminado, presentándose diferencias en cuanto a número de registros y preferencias asociadas a las probabilidades de desarrollo de las orugas. Estos resultados permiten rechazar la hipótesis planteada para el presente estudio y contestan plenamente las preguntas de la investigación.
Discusión y Conclusiones
Los hallazgos generales del trabajo complementan los estudios anteriores (Young 1971; Rausher 1979; Scriber y Feeny 1979; Brown et al. 1981; Miller y Feeny 1989; Racheli y Pariset 1992) al comprobar que Battus polydamas utiliza de varias especies de Aristolochia como hospederos, ratificando su condición de «polífago» de este género de plantas, lo que explica en buena parte el amplio areal que presenta la mariposa. La distribución de una postura por vástago o individuo de Aristolochia, la ubicación de éstas en zonas apicales y el color conspicuo de los huevos coincide con patrones encontrados en otras mariposas aposemáticas (Gilbert 1975; Rausher 1979; Chew y Robbins 1984). Este mecanismo puede contribuir a una mejor distribución del recurso, especialmente cuando es limitado, como en el caso de los tejidos jóvenes o asimilables de hospederos de alta consistencia. Por otra parte, la distribución normal del número de huevos, independiente de la abundancia del recurso, difiere de trabajos previos en mariposas del mismo género (Rausher 1979; Chew y Robbins 1984), donde se propone que la hembra define el número de huevos de acuerdo a la disponibilidad del recurso.
En cuanto a la preferencia de Battus polydamas por hospederos con una determinada forma de hoja, los resultados coinciden con lo observado por Rausher (1978, 1979, 1980, 1981, 1983) en B. philenor, la cual comparte la preferencia por las zonas apicales y los tejidos jóvenes como sitios de oviposición o de alimentación de las orugas en especies coriáceas, así como la ubicación indiscriminada de las orugas en especies suculentas. Esto apoyaría una preferencia generalizada de Battus por este tipo de tejidos.
De acuerdo con Miller (1987), los ácidos aristolóquicos son estimulantes del proceso de alimentación de las orugas de Battus polydamas, por lo cual su presencia en las aristoloquias (Chen y Zhu 1987) es importante en la identificación del hospedero por parte de hembras y orugas; en adición, teniendo en cuenta la alta especificidad comentada de la mariposa por sitios de oviposición próximos al ápice de los vástagos, junto con el amplio rango de hospederos registrados, se considera que la susceptibilidad de un hospedero identificado (calidad de una planta para ser invadida por un fitófago) estaría también determinada por la cantidad de tejido joven o por el grado de esclerofilia que presenten sus órganos.
De acuerdo con Loveless (1961, 1962, citado por Rausher 1980), el menor tiempo del ciclo de desarrollo y la menor mortalidad en las orugas alimentadas con especies de alto grado de suculencia y baja esclerofilia podría ser la resultante de la mayor concentración de nutrientes, en especial de nitrógeno, que hay en este tipo de tejidos. Esto apoya la hipótesis de Scriber y Fenny (1979), Harborne (1982) y Loader y Damman (1991), quienes señalan que este nutriente es determinante en la tasa de supervivencia de las orugas. Se recalca que la alta mortalidad debida a la baja calidad nutricional, a la que se suma la prolongación del período de desarrollo de las sobrevivientes con el consiguiente aumento en el periodo de exposición a depredadores, puede generar una población de adultos extremadamente baja, fenómeno que podría ser el origen y explicación de la selectividad de las hembras en cuanto al hospedero.
En este sentido, si bien en las pruebas realizadas con orugas no se observó una preferencia marcada, los registros de inmaduros observados en campo hicieron evidente las preferencia por determinadas especies de Aristolochia, especialmente aquellas donde la mortalidad y el período de desarrollo fueron menores; la hipótesis del trabajo es entonces rechazada y los resultados apoyan la opinión de Singer (1984) en cuanto a que generalmente existe un hospedero «óptimo», el cual es reemplazado por otro de menor calidad cuando el primero no se presenta. Los resultados coinciden también con los de Chew (1977) y Rausher (1979) al mostrar que el hospedero más utilizado es aquel que brinda un mayor potencial de sobrevivencia a las orugas, ratificando la importancia de la selección del sitio de oviposición.
Llama la atención el número mayor de orugas de Battus polydamas en el envés foliar; esto podría ser parte de su comportamiento defensivo, junto con la coloración aposemática y las secreciones de los osmaterios (Honda 1983 y Damman 1986, citados por Miller 1987; Brower 1984), por lo que se recomiendan más estudios al respecto. Dado que las orugas y los adultos acumulan ácidos aristolóquicos (Urzúa y Priestap 1985) y son rechazadas como alimento por parte de aves depredadoras (Brower y Brower 1964; Brower 1984), es probable que la presión mayor de depredación se deba a invertebrados. Los casos de parasitismo en huevos y orugas coleccionadas en campo, coincidentes con lo registrado en otros trabajos (Young 1971; Rausher 1980; Brown et al. 1981; Miller 1987; Racheli y Pariset 1992), muestran alto grado de parasitismo por himenópteros (Braconidae y Chalcidioidae) y dípteros (Tachinidae), lo que hace pensar que la protección química, que les defiende de vertebrados, no es muy efectiva contra estos insectos. De igual forma, se tiene que el polimorfismo en las orugas de B. polydamas apoya la hipótesis de Brown et al. (1981), en el sentido de que los patrones de coloración no son geográficamente homogéneos y que varían dentro de una población; en este sentido, estamos de acuerdo con Clarke et al. (1963), quienes señalan que la coloración de las orugas está determinada genéticamente, y que la proporción de uno u otro patrón en la población puede depender de qué tan inconspicua o aposemática sea para los depredadores.
Footnotes
Agradecimientos
Esta investigación constituyó parte del trabajo de grado de los dos primeros autores. Agradecemos al Instituto de Ciencias Naturales y el Departamento de Biología de la Universidad Nacional de Colombia. Al Fondo "José Celestino Mutis" de la FEN-Colombia, financiadora del estudio. A Indiana Bustos y Frank G. Stiles por su apoyo en el análisis estadístico. A Frank G. Stiles, Orlando Vargas y Liz Aristide por sus valiosos comentarios y observaciones. A Nury Quintero, Gustavo Lozano, Roberto Jaramillo, Javier Estrada, Javier Fuertes, Natalia Florez, Camilo Torres y Clemencia Gómez por su apoyo en diferentes fases del estudio. A Fredy Castellanos y Sparta por su cooperación.