
Abstract
The ant assemblaje was used for first time in Colombia to study fragmentation in Tropical Dry Forest and to show the biological value of the remaining forests. Fourteen samplings were made in seven forest fragments and their surroundings, all of them located along Cauca river valley, in either of its margins, and in a range of 220 km from north to south (Valle and Cauca departments). Each forest was visited twice. During each visit, both forest and matrix (the surroundings) were sampled in a four-100 m. transect system. Each transect included 40 stations, 160 tramps, and 40 litter samples. Additionally, eight hours of manual collection were invested per visit. From a total of 3006 ant captures, 2093 were made inside forest and 913 in matrix. From a total of 137 ant species, 123 were captured at least once inside a forest and 74 in a matrix. A highly significant correlation between all forests and their respective matrix richness was found. In all cases, more than 50% (until 89%) of the ant species were captured only in forest (never in its matrix). Shannon diversity index inside the forest fragments fluctuated between 1.96 and 3.52. Shannon diversity indexes were significantly higher in forest than in matrix in all cases except one. There was no statistical relationship between area and richness in this set of forests, probably because all of them are very small forest fragments fluctuating between 4-15 ha.
Introducción
Colombia ocupa un lugar privilegiado como poseedor de especies biológicas, siendo considerado, junto con Brasil y Perú, como uno de los países de megadiversidad (Brown 1989) y poseedor de varios centros de endemismo, al igual que el Ecuador (Terborgh y Winter 1983). Sin embargo, es, también, uno de los países con mayor destrucción del bosque húmedo tropical en el mundo, con aproximadamente 600.000 ha anuales de bosque talado. La región de los Andes, que concentra la mayoría de la población colombiana, es la más afectada por la acción del hombre (Saavedra y Freese 1986) con el consecuente retroceso acelerado y fragmentación de los hábitats naturales (Van Velzen 1991).
Dentro de la gran variedad de zonas de vida que se presentan en los Andes colombianos, la zona de bosque seco tropical se localiza en las planicies aluviales de los valles del río Cauca y del río Magdalena, que corren en sentido sur-norte entre las cordilleras occidental y central, en el primer caso, y entre la central y oriental, en el segundo. La importancia biológica del bosque seco tropical es sustentada por Janzen (1988a, 1988b), quien sostiene que éste es el más amenzado de los grandes hábitats de bosque tropical de tierras bajas. El presente estudio se enfocó hacia la franja de bosque seco tropical del valle geográfico del río Cauca, suroccidente de Colombia, cuya importancia biológica y el acelerado deterioro hacen resaltar la urgencia de buscar información y proponer soluciones para esta problemática.
El fraccionamiento del hábitat puede afectar la diversidad de los insectos en diferentes formas de acuerdo con las necesidades de cada especie, sus propiedades (movilidad, dinámica poblacional), las condiciones en que se presenten los fragmentos (aislamiento, dimensiones) (Brown 1989) y los estados sucesionales (Usher y Jefferson 1989).
Es así, como algunos taxa de insecta han sido propuestas como grupos indicadores de biodiversidad y de deterioro ambiental. Entre éstos, están los Coleoptera Cicindélidos (Pearson y Cassola 1992), Lepidoptera Heliconiidae y Formicidae. De acuerdo con Brown (1989) y Pearson (1993), las especies indicadoras deben ser de fácil detección, de fácil evaluación y deben proveer valiosa información.
Brown (1989) evaluó una serie de 15 grupos de insectos que se destacan como indicadores, teniendo en cuenta 12 criterios. Las hormigas, junto con un grupo de lepidópteros, recibió el máximo puntaje de 21 puntos sobre 24 totales. Las hormigas (Hymenoptera: Formicidae) se han destacado por sus posibles cualidades como indicadores de biodiversidad (Roth
Teniendo en cuenta los antecedentes anteriores, se planteó el presente trabajo, el cual pretende determinar la importancia faunística relativa de las manchas relictuales de bosque seco con base en especies de la familia Formicidae y relacionar la diversidad y composición de hormigas.
Area de estudio
El valle geográfico del río Cauca comprende la zona plana de la cuenca del Alto Cauca, con una extensión de 421.000 ha., encontrándose rodeado por las cordilleras central y occidental, entre los departamentos del Valle y del Cauca.
El departamento del Valle del Cauca se ubica en la región central del occidente colombiano. La fisionomía dominante del valle es el río sinuoso que, con inundaciones estacionales de terreno bajo, mantiene madreviejas y humedales efímeros (Alvarez-López y Kattán 1995). Se sabe que este valle, alguna vez, fue dominado por extensa vegetación arbórea que fue destruída, casi en su totalidad, para implantar ganadería y cultivos, como caña de azúcar, algodón, sorgo, soya, maíz, tabaco, fríjol, hortalizas y otros. (Espinal 1968; Velasco 1982; Corporación Autónoma Regional del Valle del Cauca, C.V.C. 1990).
El objeto de estudio de este proyecto fueron siete fragmentos de bosque secundario, situados en la zona de bosque seco tropical (bs-T) del valle geográfico del río Cauca. Los bosques se sitúan a una altura sobre el nivel del mar entre 950 y 1000 m.; con una temperatura superior a los 24°C y la precipitación anual promedio fluctúa entre 1000 y 2000 mm anuales.
El valle geográfico del río Cauca se dividió arbitrariamente en tres zonas (sur, centro y norte), en donde se escogieron los siguientes bosques, todos pertenecientes al departamento del Valle del Cauca, a excepción del bosque de San Julián, que pertenece al departamento del Cauca:
Zona sur: Bosque San Julián (municipio de Santander de Quilichao, 6 ha de extensión) y bosque Colínder (o Colindres) (municipio de Jamundí, 12,3 ha)
Zona Centro: bosque El Hatico (municipio de El Cerrito, 7 ha), bosque El Vínculo (Municipio de Buga, 15 ha) y bosque Las Chatas (municipio de Buga, 8,7 ha).
Norte: bosque El Medio (municipio de Zarzal,10 ha) y bosque Las Pilas (municipio de Zarzal,14,8 ha).
Cada fragmento boscoso está rodeado por una matriz. Esta se define como el terreno antrópico circundante a los fragmentos boscosos. En este estudio las matrices fueron: matriz de caña (San Julián y El Medio), matriz de potrero ralo (Las Chatas y Las Pilas) y matriz de potrero arbolado (Colinder, El Hatico y El Vínculo).
Metodología
El estudio comprendió un total de 14 muestreos intensivos, dos en cada uno de los bosques descritos, realizados entre agosto de 1994 y febrero de 1995. La estrategia consistió en realizar dos veces lo que se llamará "serie de muestreos", en la cual se visitaron todos los bosques, en el menor tiempo posible, entre muestreo y muestreo.
En cada bosque, con ayuda de una brújula y un decámetro, se trazaron cuatro transectos de 100 metros cada uno, dos hacia el interior del bosque, comenzando a los 20 m desde el borde y dos hacia el exterior y enfrente de los primeros, pero comenzando a 20 m del borde hacia afuera. Los dos transectos dentro del bosque eran paralelos y estaban separados entre sí por 50-70 m. Los dos transectos hacia el exterior del bosque, es decir, en la matriz antrópica circundante al bosque, estaban dispuestos de la misma manera (Fig. 1 a).
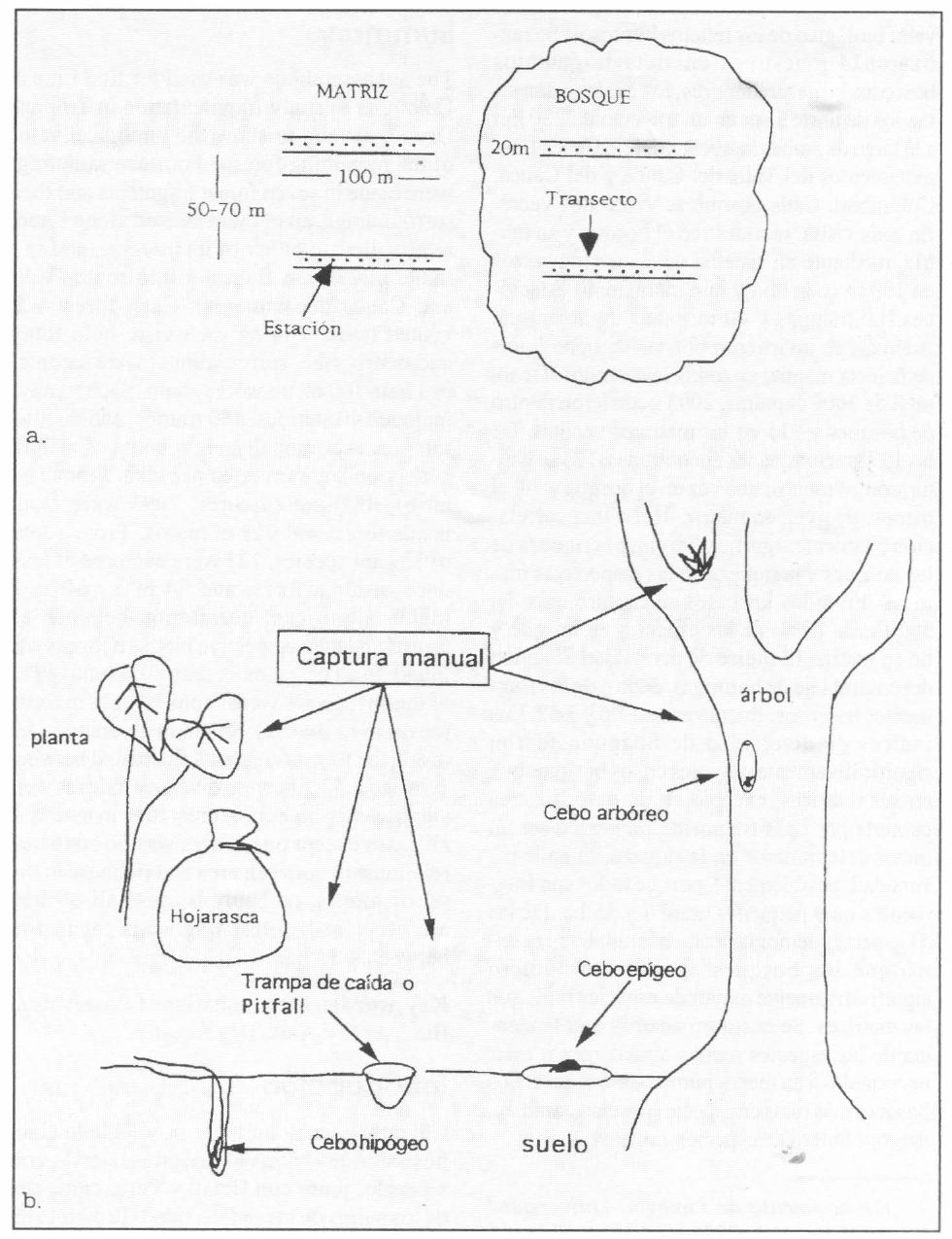
a. Esquema de un fragmento boscoso y su matriz vecina, mostrando el diseño de transectos empleado durante el estudio. b. Esquema de una estación de muestreo.
En cada transecto se marcaron 10 estaciones equidistantes (10 m) para muestrear hormigas con ayuda de cebos, trampas de caída, colecta de hojarasca y captura manual, como se detalla a continuación y se visualiza en la figura 1 b.
En cada estación se colocaron tres cebos: El primer cebo es de tipo corner modificado. El tubo se llenó con atún como atrayente y se enterró 10 cm bajo la superficie del suelo, por lo cual se llamó cebo hipógeo. El segundo cebo, consistente de un trozo de atún sobre una hoja de papel bond de 22 cm x 8 cm., se ubicó sobre el nivel del suelo y se le llamó cebo epígeo. El tercer cebo, comparable al segundo, se situó a 1,70 m de altura adherido al tronco de un árbol y se le llamó cebo arbóreo. Cada cebo se dejó actuar entre cuatro y cinco horas. Al terminar este tiempo, se procedió a recoger los cebos con las hormigas atraídas e inmediatamente se introdujeron en frascos con alcohol al 70% debidamente rotulados, los cuales, posteriormente, se refrigeraron hasta el momento de separar las hormigas en el laboratorio.
Además, en cada estación se instaló una trampa de caída (modificada de Jaffé
Adicionalmente, en cada estación, se colectaron cinco arrastres manuales de hojarasca, de 50 cm de longitud cada uno, que se depositaron en una bolsa plástica negra de 60 cm de ancho por 70 cm de largo. Al final de cada transecto, se pasó la hojarasca por un saco cernidor tipo Winkler (Jaffé
Finalmente, en cada transecto, se hizo un muestreo manual durante dos horas, buscando, equitativamente, en todos los sustratos accesibles, como suelo, hojarasca, epífitas, vegetación de sotobosque, troncos en descomposición, ramitas huecas o secas y árboles en pie. Las hormigas capturadas forrajeando se introdujeron en frascos rotulados con el sitio específico en que se encontraron. Los nidos encontrados se dispusieron en frascos separados y rotulados, tratando, en lo posible, de capturar representantes de todas las castas.
Resultados y Discusión
Riqueza y ubicación habitacional de las especies
Un total de 3006 capturas de hormigas, 2093 dentro de bosques y 913 en las matrices se realizó. El sitio más rico fue El Hático, con 81 especies, le siguen El Medio, con 60, Las Pilas, con 60, El Vínculo, con 49, Las Chatas, con 43, Colinder, con 43 y San Julián con 38 especies.
El total de 137 especies encontradas en el estudio corresponde, aproximadamente a la quinta parte del total de especies de Formicidae registradas, hasta la fecha, para el país (Fernández 1995), cifra muy considerable, dada la limitada extensión de los fragmentos estudiados.
La ubicación habitacional de las 137 especies encontradas en el estudio se exhibe en el esquema de la Figura 2: 123 se capturaron, al menos, una vez en bosque. De estas 123 especies, el 51% siempre fueron capturadas en bosque y nunca en la matriz, el 49% especies, al menos, una vez en ambos hábitats y el 11% especies se capturaron, sólo, en matriz.
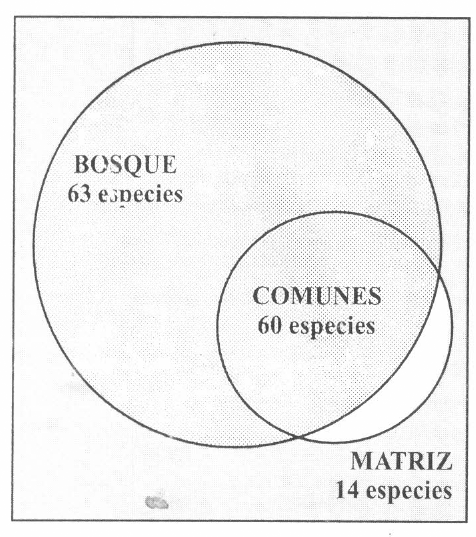
Ubicación habitacional de las 137 especies de hormigas encontradas en el estudio.
En cada uno de los siete sitios, se observó la misma tendencia, al encontrarse un mayor número de especies en bosque que en matriz. El bosque tuvo entre 53% y 79% de especies exclusivas, mientras que las especies exclusivas de matriz fluctuaron entre el 9 y el 19%.
Al comparar la riqueza de los bosques contra la riqueza de las matrices (Fig. 3), se encontró que era, significativamente, mayor en el bosque que en la matriz (prueba de t pareada, t=13,79; g.1.13; p<0,001).
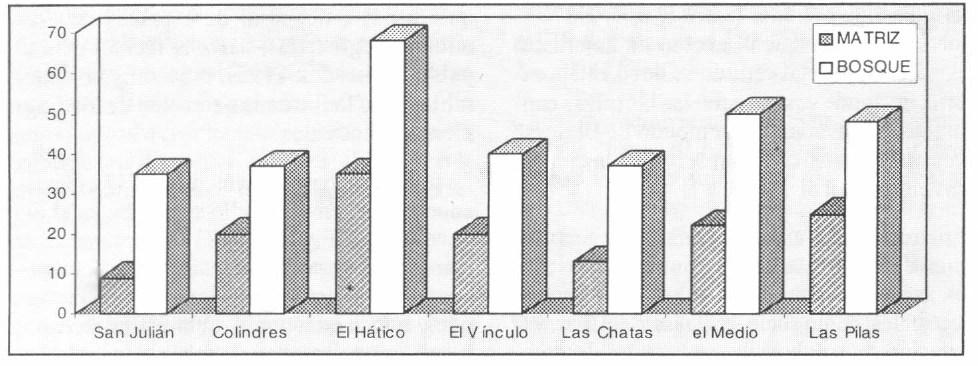
Discriminación de riqueza entre bosque y matriz (los bosques están dispuestos de norte (izquierda) a sur (derecha) del Valle del río Cauca.
Igual resultado se encontró al comparar individualmente la riqueza por estación entre cada bosque con su matriz mediante prueba de tó de Mann Whitney, siempre fue significativamente mayor en el bosque (Tabla 1).
Comparación de la riqueza de especies de hormigas capturada en bosque y matriz, usando, sólo, trampas y cebos. n = 40 en todos los casos.
Al comparar la riqueza entre los bosques, se observa que el bosque más rico (El Hático) tuvo 66 especies, casi duplicando al más pobre (San Julián), con 34 especies.
La proporción de las barras (Fig. 3), también, muestra que la riqueza de la matriz parece seguir a la del bosque o viceversa. Efectivamente, se encontró una regresión altamente significativa entre la riqueza del bosque y la de la matriz (Fig. 4), con coeficiente de determinación R2=0,705 y t= 5.35; g.l.=13; p<0,001.
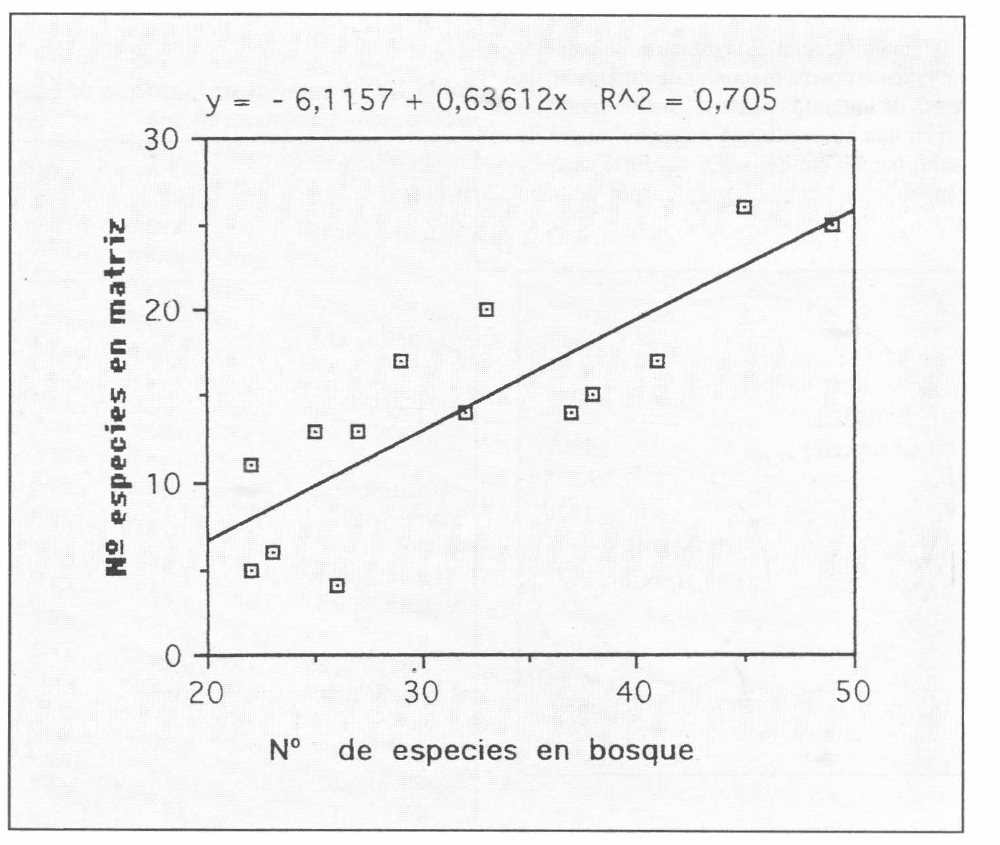
Regresión simple entre el número de especies en bosque y el número de especies encontradas en la matriz, en todos los sitios y muestreos (p<0,001). y = - 6,1157 + 0,63612x R^2 = 0,705
Indice de diversidad de Shannon
La tendencia observada en la riqueza se reflejó, para cada bosque y su matriz, en los índices de diversidad Shannon. (Tabla 2).
Indices de diversidad Shannon (H') en cada bosque y su matriz.
Cuando se disponen los índices de diversidad en una forma ordenada, de mayor a menor, se observa que el índice de diversidad del bosque más pobre (San Julián) se sitúa entre los índices de diversidad de las matrices de otros bosques ricos y el índice de diversidad de la matriz del bosque más rico (El Hático) se sitúa entre los índices de diversidad de otros bosques.
Debido a estos resultados, se propuso, entonces, que estas manchas boscosas tienen diversidades altas (El Hático, El Medio y Las Pilas), diversidades medias (El Vínculo, Las Chatas, Colinder) y un bosque de baja diversidad, que fue San Julián.
Especies exclusivas en los bosques
Se encontró que a medida que un bosque se hace más rico, tiene mayor cantidad de especies exclusivas de ese bosque (Fig. 5), por lo cual es reservorio no, sólo, de mayor diversidad, sino, también, de especies raras.
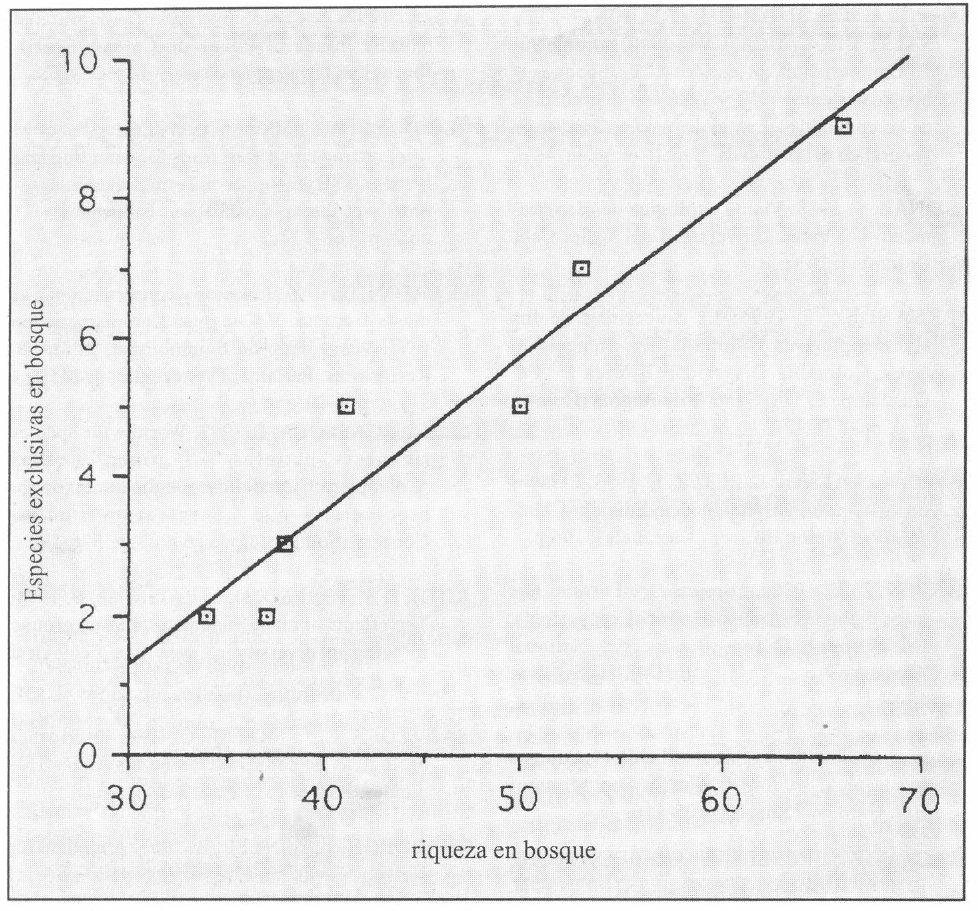
Regresión entre el número de especies exclusivas de los bosques y la riqueza (y= - 5,37 + 0,22x; R2=0,92).
Curva especies-área
Se encontró que no había relación entre la riqueza y el área de los bosques estudiados (p>0,5) (Fig. 6), hecho que se explica porque el área de estos fragmentos está dentro de la misma escala, fluctuando en el orden de 6 a 15 ha.
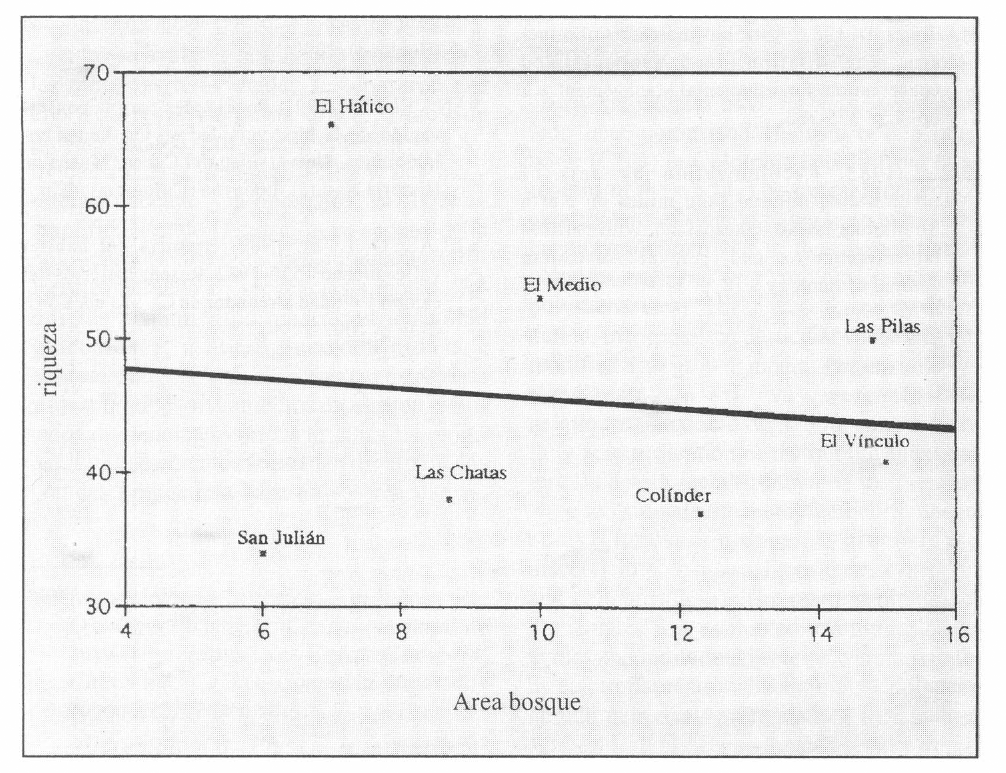
Riqueza de especies de hormigas en función del área de los bosques muestreados.
Especies compartidas
Al examinar las relaciones en cuanto a especies compartidas por los fragmentos boscosos, se encontró que los bosques más ricos, siempre, compartieron mayor número de especies con los demás bosques. Esto se refleja en la Tabla 3, en donde El Hático y El Medio, siempre, compartieron mayor número de especies con todos los demás bosques y entre sí.
Número de especies que comparten cada par posible de bosques. S.J.:San Julián, Co: Colínder, Ha: El Hatico, Ví: El Vínculo, Cha: Las Chatas, Me: El Medio y Pi: Las Pilas.
El bosque más rico, El Hático, siempre compartió el mayor número de especies exclusivas de dos bosques con todos los demás fragmentos boscosos, hecho que muestra la necesidad de enfatizar en la protección de estos relictos especiales. A pesar de lo anterior, no todas las especies de bosques pobres están contenidas en los bosques ricos y cada remanente boscoso constituye, también, un aporte a la biodiversidad gama o regional en el valle geográfico.
Los resultados encontrados durante este estudio están mostrando que la mirmecofauna de los fragmentos boscosos no está siendo predominantemente influida por variables biogeográficas, como área del fragmento o distancia entre los fragmentos y que, posiblemente, otras variables, como podrían ser grado de perturbación o historia del manejo de cada fragmento y de sus alrededores, que podrían ser de importancia para determinar el número y la identidad de las especies que se conservan en cada uno.
Conclusiones
Todos los fragmentos boscosos estudiados en la zona plana del valle geográfico del río Cauca conservan mayor riqueza faunística en especies de hormigas que sus respectivas matrices antropogénicas vecinas, razón por la cual tienen importancia biológica suficiente como para propender por su conservación.
123 especies de hormigas en un área total de 74 ha de relictos boscosos fragmentados, aislados y relativamente perturbados es una cifra inusitadamente alta, con respecto al número de hormigas descritas para el Neotrópico. Con esto, se demuestra la importancia faunística relativa de estos bosques.
Dada la pequeña extensión de estos bosques, la riqueza en hormigas no es una función del área de cobertura y, tanto la riqueza, como la biodiversidad, parecen estar determinadas por otros factores, que podrían ser la historia y manejo de cada uno de los relictos.
La conservación de uno solo o unos pocos de los fragmentos boscosos no es suficiente para la óptima preservación de la biodiversidad gamma o regional, pues todos los bosques aportan, con especies propias posiblemente por haber atrapado muestras al azar de la fauna que se encontraba en el bosque continuo inicial.
La matriz antropogénica que domina el paisaje de la zona plana del valle geográfico del río Cauca, también, aporta a la biodiversidad global de Formicidae, pero, a diferencia de los fragmentos boscosos, no está en peligro inmediato de desaparecer.
Footnotes
Agradecimientos
El presente estudio fue financiado por el Programa de Becas para la Conservación, WCS/ GEA/FES (Wildlife Conservation Society, GEA y Federación para la Educación Superior de Colombia), el Fondo José Celestino Mutis de la Financiera Eléctrica Nacional (FEN) y la Universidad del Valle. A estas entidades, los autores agradecen su valiosa colaboración.