
Abstract
To screen nine commercial cultivars and 13 clones of potato of the Colombian Central Collection (CCC), the plastic container test was used. Potato tubers of approximately 60 g were buried into a layer of vegetal soil inside the plastic containers. The tubers were artificially infested with a known number of first instar larvae of the Andean weevil. The food preference of the adults was also investigated among the clones of the CCe. All the experiments were carried out under screenhouse conditions, at lCA's National Center of Research «Tibaitatá» in Mosquera (Cund.). All the screened materials were very susceptible to the attack of the insect on the tubers. The results are discussed based on recent published research.
Introducción
El gusano blanco de la papa, Premnotrypes vorax (Hustache) (Coleoptera: Cur culionidae), es la plaga más importante de la papa en las principales zonas pro ductoras de Colombia, Ecuador y Vene zuela. El control de esta plaga, en la región andina, se basa principalmente en el uso de insecticidas.
Desafortunadamente no existe un umbral de acción definido que permita a los agricultores un uso racional de los insec ticidas, debido, en gran parte, a las carac terísticas propias del insecto. Durante los meses de enero y febrero, los adultos de P. vorax, para las condiciones de la ve reda Sote Panelas, en el municipio de Motavita (Boy.), se pueden encontrar en grandes cantidades durante el día, escon didos en sus «refugios naturales» debajo de terrones grandes. Este tipo de refugio permite a los adultos protegerse de la luz solar y, también, les proporciona un nivel adecuado de humedad. Una vez iniciadas las siembras y después de la emergencia de las plantas, los adultos tienden a con gregarse al pie de éstas e inmediatamente empiezan a enterrarse buscando las raíces. Muestreos efectuados durante todo el ciclo del cultivo demuestran que en el campo hay adultos hasta aproximada mente dos meses después de la emergencia de las plantas (Valencia 1977).
En la literatura consultada no existe un trabajo que relacione la densidad de po blación de los adultos de la plaga al comienzo de la estación de cultivo, con el daño en los tubérculos al momento de la cosecha. Por esta razón, los agricultores realizan entre dos y cuatro aplicaciones de insecticidas para el control de la plaga, según la frecuencia e intensidad de las lluvias.
La resistencia genética de plantas contra una plaga específica puede influir direc tamente en la reducción del número de aplicaciones de insecticidas para su control. Los trabajos de búsqueda de resistencia genética contra el gusano blan co de la papa se iniciaron en 1969 (Espinel y Torrado 1969). Posteriormente, en prue ba de laboratorio, Valencia (1977) regis tró a la especie silvestre Solanum polya denium Greenm. como resistente al daño de las larvas.
Alcázar (1988) investigó, bajo condicio nes de campo e invernadero, el nivel de susceptibilidad al daño de los adultos del gorgojo de los Andes, P. suturicallus Kuschel, en el follaje, y el de las larvas en los tubérculos de cinco entradas de cada una de las especiesSolanum stenotomum, Juz. et Buk., S. juzepczukii Buk., S. tuberosum ssp. andigena J.S. Hawkes, S. acaule Bitt., y dos entradas de S. ajanhuiri Juz. et Buk. Bajo condiciones de campo, encontró diferencias de sus ceptibilidad estadísticamente significa tivas entre las especies de Solanum y entre los clones de cada especie, en cuanto al daño de los adultos en el follaje y de las larvas en los tubérculos. Bajo condiciones de invernadero también encontró dife rencias estadísticamente significativas en relación con la longevidad de los adultos, la oviposición de las hembras, el daño del adulto en el follaje y el de las, larvas en los tubérculos.
Zenner de Polanía (1990), en un resumen sobre los trabajos de investigación reali zados por el grupo de entomólogos del Instituto Colombiano Agropecuario -ICA en el cultivo de papa, menciona como promisorio un grupo de clones de la Colección Central Colombiana (CCC). Estos clones fueron seleccionados bajo condiciones de campo, entre 1983 y 1987. Los nueve clones que ella menciona como de interés para continuar investigando, mostraron un promedio de daño, en tuberculos, de 48,67% ± 10,49. La mayor parte de este daño se debe a larvas de una segunda generación y por lo tanto la autora concluye que este hecho estaría indicando que el «factor de resistencia» podría ser una sustancia química presente en la planta después del aporque del cultivo y que ésta luego se diluye a medida que las plantas maduran.
La mayor parte de las investigaciones sobre resistencia genética al gusano blan co de la papa se realizaron bajo condicio nes de campo, en las cuales, los clones citados como resistentes por Zenner de Polanía (1990) siempre tuvieron larvas de gusano blanco causando daños en los tubérculos, lo que estaría indicando una ausencia de antibiosis (Painter 1968). De igual manera, un menor índice de daño en los tubérculos de esos clones en compa ración con otros incluídos en la investi gación, podría indicar que, bajo condi ciones de campo, estaría operando un mecanismo de no preferencia o antixeno sis (Painter 1968; Kogan y Ortman 1978).
En la literatura entomológica está bien documentado el hecho de que en algunos órdenes de insectos, como Lepidoptera y Diptera, los requerimientos nutricionales de los adultos son diferentes a los de las larvas. Por lo tanto, cuando una hembra selecciona un lugar para ovipositar no obtiene información sensorial sobre la constitución nutricional de las plantas, mientras que las larvas sí son sensitivas a estos componentes nutricionales al momento de alimentarse (Wiklund 1974; Beck y Schoonhoven 1980). Esta dife rencia sugiere diferentes patrones de comportamiento y, posiblemente, facto res genéticos diferentes (Wiklund 1974).
Con la intención de profundizar en el conocimiento acerca de la resistencia genética al gusano blanco de la papa y de entender mejor la forma como se produce la interacción entre el insecto y la planta hospedante, se iniciaron trabajos de ta mizado de cultivares comerciales y de los clones citados previamente como re sistentes.
Materiales y Métodos
Los trabajos experimentales se efectua ron bajo condiciones de invernadero en el Centro de Investigaciones «Tibaitatá» del ICA, en Mosquera (Cund.).
Para el tamizado de los diferentes culti vares y clones se utilizó la prueba del envase plástico. En envases plásticos de 12,5 cm de diámetro y 8 cm de altura, se puso una capa de suelo vegetal de 1 cm de altura, encima de esta capa se colocó un tubérculo del cultivar o clon a evaluar y luego se tapó con suelo vegetal. La capa de suelo cubrió totalmente el tubérculo hasta unos 0,5 cm por encima del borde superior del mismo. Los tubérculos se infestaron artificialmente con un número conocido de larvas de primer instar del gusano blanco de la papa (Fig. 1a).
Cortes transversales de las unidades experimentales utilizadas. a. Para el tratamiento de los cultivares comerciales y de los clones de la CCC, b. Para la prueba de preferencia de alimentación de las larvas. La flecha indica el lugar de infestasión con las larvas de P. vorax.
Para la obtención de las larvas de primer instar, en el campo se colectaron adultos de gusano blanco, los cuales, general mente, se encuentran en sus «refugios naturales», debajo de terrones grandes. Las poblaciones de adultos son más a bundantes en campos que mantienen siembras consecutivas de papa. Los adul tos colectados se llevaron al invernadero y se depositaron en número de 50 indivi duos por cada caja de plástico de 35 x 22 x 12 cm. La tapa de cada caja de plástico se perforó y encima se colocó una tela porosa de popelina, para favorecer el intercambio gaseoso. Como fuente de alimento para los adultos se utilizó una mezcla de hojas y tajadas de tubérculos de papa criolla (S. phureja Juz. et Buk.)(Valencia y Bohórquez 1992).
Como substrato de oviposición se utili zaron tallos de cebada de 4 cm de longitud, los cuales se trataron previamente con alcohol etílico al 80% para la eliminación de ácaros. Entre 10 y 15 porciones de tallos de cebada se utilizaron por cada caja de oviposición. Estos tallos se cambiaron semanalmente, y los usados se colocaron en envases de plástico para su incubación. Bajo las condiciones de invernadero en «Tibaitatá», con una temperatura media de 17°C, la eclosión de los huevos se produjo a los 19 días, aproximadamente.
Ensayo Preliminar
Para determinar el número óptimo de larvas de primer instar del gusano blanco de la papa emplear en los ensayos, se estableció el siguiente experimento: u sando el arreglo experimental descrito, se probaron diferentes densidades de infestación en tubérculos de la variedad Parda Pastusa. Las densidades investiga das fueron: 3, 6, 9, 12, 15, 18 y 21 larvas del primer instar por tubérculo. Cada densidad tuvo tres repeticiones. El expe rimento se evaluó a los 45 días, cuando las larvas del último instar estaban aban donando los tubérculos: Se contó el nú mero de larvas que se encontraban en el suelo y, también, las que aún se encon traban dentro de los tubérculos.
Tamizado de cultivares comerciales
Los siguientes cultivares comerciales fueron evaluados en esta prueba: ICA-San Jorge, Diacol-Capiro, ICA-Chitagá, Dia col-Monserrate, ICA-Nariño, ICA-Pica cho, Parda Pastusa, ICA-Puracé y Crio lla. En esta prueba se usaron 10 larvas de primer instar por tubérculo, y cada cultivar tuvo cinco repeticiones. La evaluación se hizo como en el ensayo preliminar.
Tamizado de clones
Los clones de la CCC citados por Zenner de Polanía (1990) como resistentes al daño de las larvas del gusano blanco de la papa también fueron evaluados. Estos clones fueron: 4342, 4769, 4269, 4654A, 4636, 4362, 4337, 4273, 4251, 4766, 4350, 4268 у 4345. En esta prueba se usaron 10 larvas de primer instar y cada clone tuvo tres repeticiones
Investigación sobre preferencia
Para determinar la posible existencia de preferencia del adulto del gusano blanco de la papa por algún cultivar o clón en particular, se realizó el siguiente experi mento: en camas de 1,5 x 1,5 m, construi das sobre el suelo, se sembraron en un arreglo pentagonal, los clones 4337,4269, 4273, 405 y el cultivar susceptible Crio lla (CCC 81). La distancia de siembra entre semillas fue de 40 cm y el experi mento tuvo cuatro repeticiones.
Cuando las plantas tenían aproximada mente 15 cm de altura (30 días después de la siembra), en cada repetición se liberaron 50 adultos de P. vorax, sin separar los sexos. Posteriormente se hi cieron las prácticas culturales de rutina (deshierba, aporque, riegos) y se dejó el experimento hasta la cosecha, cuando se determinó el peso y el número de tubér culos sanos e infestados, así como también el número de larvas encontrado de cada clón. Debido a que en la cosecha se dispuso de tubérculos frescos de los clones 405, 4269 y 4337, estos, junto con tubér culos de Criolla, fueron tamizados por medio de la prueba del envase plástico, utilizando 20 larvas de primer instar y tres repeticiones. Para evaluar este expe rimento se siguió la metodología usada en el ensayo preliminar.
Preferencia en la Alimentación de Adultos
De algunos de los clones utilizados en el experimento sobre preferencia se toma ron folíolos terminales de la tercera hoja más nueva de las plantas, para investigar la posible existencia de alguna preferencia para la alimentación de los adultos de P. vorax. Para estandarizar el tamaño de la unidad experimental, con la ayuda de un sacabocado (Cork borer, de Helix Inter national, England), se prepararon discos de hojas de 1,6 cm de diámetro (2 cmβ). Los discos de hojas de los clones 4337, 4269, 4273 y del cultivar Criolla se dis tribuyeron de manera equidistante dentro de cajas de petri de 15 cm de diámetro y 3 cm de altura sobre un disco de papel de filtro humedecido con agua destilada, para evitar el desecamiento de los discos de hojas. En cada caja de petri se liberó una pareja de adultos de P. vorax y el experimento se evaluó cuatro días des pués. El experimento tuvo nueve repeti ciones y, al final de la prueba, se midió el área foliar de los residuos de los discos, con la ayuda de un medidor portatil de área foliar.
Preferencia para la alimentación de las larvas
Con el fin de determinar si las larvas del primer instar son capaces de reconocer y posteriormente colonizar una fuente de alimentación en particular, se realizó este experimento. En cada envase plástico, similar a los utilizados para el tamizado de los cultivares comerciales, se enterra ron dos tubérculos de papa de los cultiva res Parda Pastusa y Criolla, siguiendo el arreglo que se muestra en la Figura lb. Cada caja se infestó con 20 larvas de primer instar del gusano blanco. El expe rimento tuvo nueve repeticiones y fue evaluado 20 días después de iniciado, cortando los tubérculos y contando el número de larvas que se encontraban dentro de ellos.
Resultados y Discución
Ensayo preliminar
Los resultados del ensayo preliminar se muestran en la Figura 2. La línea de regresión Y= -1,576 + 0,873X, y el coeficiente de correlación r = 0,99 des criben la población de larvas de último instar recuperadas al final del ensayo. Se observa que los valores de las desviacio nes estándar son mayores cuando ocurren densidades extremas de larvas (3, 6 y 21 larvas por tubérculo). Por esta razón, para las pruebas subsiguientes se decidió usar 10 larvas del primer instar por tubér culo.
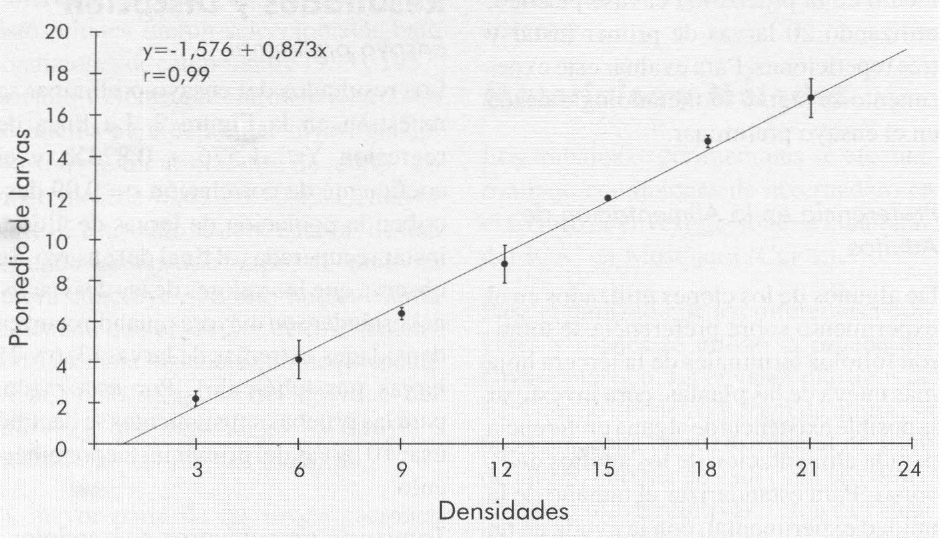
Línea de regresión obtenida de la interacción entre el número de larvas (densidades) colocadas artificialmente al inicio de la prueba y el número promedio de larvas obtenidas al final.
Tamizado de cultivares comerciales y de clones
Los resultados del tamizado de cultivares comerciales por susceptibilidad al daño de las larvas de P. vorax se muestran en la Figura 3. No hubo diferencias estadís ticamente significativas en el comporta miento de los nueve cultivares y todos fueron susceptibles al ataque de las larvas.
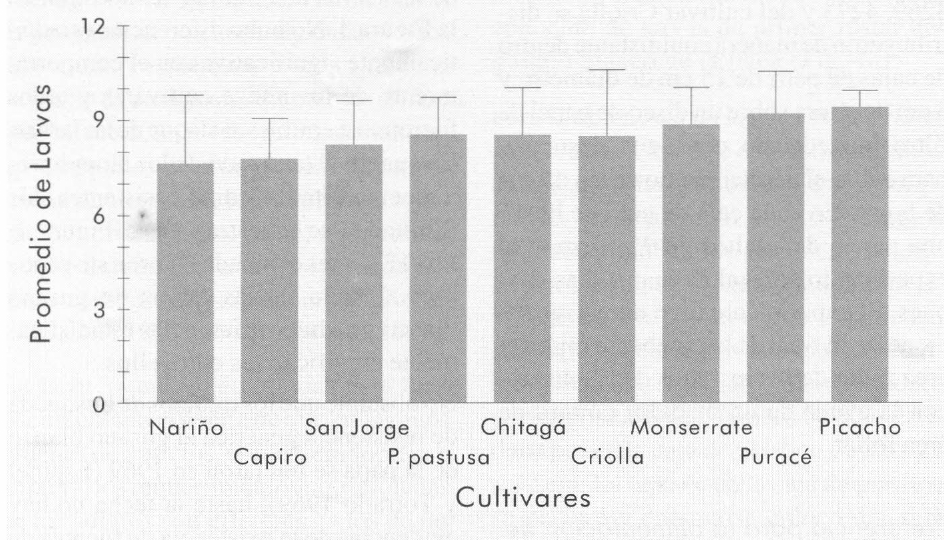
Número promedio de larvas obtenidas en la prueba de tamizado de cultivares comerciales de papa. Las líneas encima de las columnas representan el valor de la desviación estándar de cada tratamiento.
En cuanto al tamizado de los clones pre viamente citados como resistentes, los resultados se muestran en la Figura 4. Los 13 clones evaluados fueron suscepti bles al daño de las larvas de gusano blanco y no hubo diferencias estadística mente significativas entre ellos.
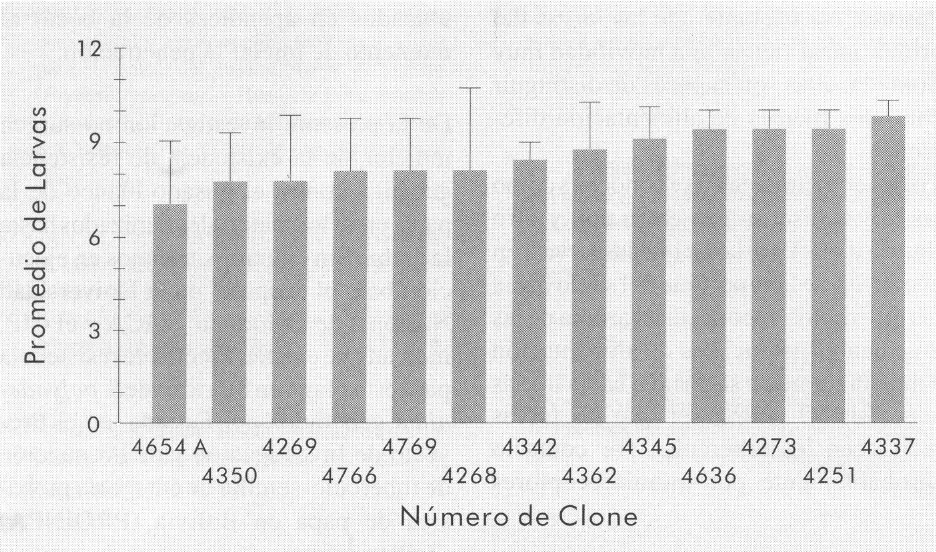
Número promedio de larvas obtenidas al final de la prueba del tamizado de los clones de la CCC con posible resistencia al daño de las larvas de P. vorax. Las líneas encima de las columnas representan el valor de la desviación estándar para cada tratamiento.
No obstante que los trabajos de búsqueda de resistencia genética al gusano blanco de la papa se iniciaron en 1969 (Espinel y Torrado 1969), hasta la fecha no hay evidencias de la existencia de fuentes de resistencia contra esta plaga en los ma teriales evaluados. Las evaluaciones rea lizadas bajo condiciones de campo (Espinel y Torrado 1969; Alcázar 1988; Zenner de Polanía 1990) se hicieron antes de conocerse el patrón de dispersión y colonización de esta plaga bajo condiciones de campo. Ahora se sabe que en campos nuevos, sometidos a un programa de rotación de cultivos, la infestación es mayor en las zonas perifé ricas del lote, y de allí hacia la parte interna del mismo, la infestación tiende a disminuir (Calvache 1 ).
Calvache, H. 1992. Comunicación personal. CENIPALMA, Apartado Aéreo 252171. Santafé de Bogotá
De otro lado, también se sabe que el patrón de dispersión en lotes sometidos a siembras consecutivas de papa es del tipo agregado (Valencia 1989a). Es decir, los adultos tienden a reunirse en determinadas zonas del lote, especial mente en las que haya un nivel adecuado de humedad (Valencia 1989a). Este tipo de agregación de los adultos da como resultado, al momento de la cosecha, un ataque larval por focos. Al producirse este tipo de ataque se tendrán cultivares o clones que dependiendo de su ubicación dentro del lote, mostrarán menores nive les de infestación, sin que esto signifique necesariamente la presencia de alguna fuente de resistencia.
De otro lado, los clones de papa citados por Zenner de Polanía (1990) como re sistentes, según la autora, sólo fueron atacados por larvas del primer instar de una segunda generación; las larvas de la primera generación fueron incapaces de desarrollarse en los tubérculos de esos clones.
Experiencias recientes demuestran cla ramente que en campos sembrados con cultivares de diferente período vegetativo, la fluctuación de la población de larvas sigue una distribuciín bimodal, es decir que se producen dos picos poblacionales de las larvas en todo el ciclo del cultivo (Valencia 1989b). Entonces, en las prue bas de tamizado por resistencia efectua das bajo condiciones de campo, donde un número grande de clones con períodos vegetativos muy variables sean plantados juntos en el mismo lote, es de esperarse que se produzcan dos picos poblacionales de larvas, tal como lo demostró Valencia (1989b). Los clones tardíos o aquellos que inician la tuberización más tarde, dependiendo de la población del gusano blanco en el campo, tendrán un daño menor comparado con los clones precoces o semiprecoces que se siembran en el mismo campo. Ese fenómeno fue expli cado en parte por Valencia (1989b).
En relación con la metodología que se ha empleado en el invernadero para la eva luación de los niveles de susceptibilidad en tubérculos, se observa que ésta no se ha estandarizado. Así por ejemplo, se han utilizado tubérculos semienterrados (Alcázar 1988) y tubérculos enterrados (esta investigación); como fuente inicial de infestación se han utilizado larvas del primer instar en sus «pajitas» (Alcázar 1988) y larvas «libres» del primer instar (esta investigación). Estas variaciones en la metodología de evaluación contri buyen a incrementar, hasta cierto grado, las fuentes de error. La metodología de evaluación debe, hasta donde sea posible, tratar de duplicar bajo condiciones con troladas, un fenómeno biológico que nor malmente sucede bajo condiciones de campo. Al no conocerse el lugar exacto de oviposición del insecto bajo condicio nes de campo (al menos en la literatura consultada no existe ninguna cita sobre este particular), entonces se debería tratar de duplicar la infestación en tubérculos enterrados, tal como sucede bajo condi ciones de campo. Los resultados experi mentales que se presentan en este trabajo demuestran valores bajos en las desvia ciones estándar, lo que estaría indicando que hasta ahora esta es la mejor manera de tamizar clones de papa para determi nar sus niveles de susceptibilidad a esta plaga.
Los resultados obtenidos por Alcázar (1988) demuestran algunas incongruen cias. Así por ejemplo en el tamizado realizado en 1979, utilizando la prueba del envase de plástico, encontró diferen cias estadísticamente significativas entre los clones probados, mientras que en la prueba de 1980 no hubo esas diferencias, no obstante que fueron probados los mis mos clones. Las diferencias encontradas se podrían explicar si se considera que la unidad de infestación empleada por Alcázar (1988) fueron cinco larvas de P. suturicallus por tubérculo. Los resultados obtenidos en el ensayo preliminar de esta investigación demuestran que las densidades extremas (6 y 21 larvas/tu bérculo) producen valores de la desvia ción estándar muy altos, lo que afecta significativamente el posterior análisis estadístico. Por otra parte, Alcázar (1988) utilizó tubérculos semienterrados, lo que posiblemente influyó en el número de larvas que penetraron en los tubérculos, aumentado de esta manera las fuentes de error para el análisis estadístico.
La prueba que se ha usado para el tami zado de los cultivares comerciales y de los clones de la CCC es muy precisa, en cuanto mide estrictamente la interacción entre las larvas del gusano blanco y el tubérculo que se esta tamizando. Como las evaluaciones se hacen al final del ciclo larval, cuando las larvas de último instar abandonan el tubérculo y luego buscan una zona donde empupar, las evaluaciones en este preciso momento pueden, de un lado, permitir la detección de la influencia de la dieta (tubérculos) en la supervivencia de un número cono cido de larvas y, de otro lado, permitir la determinación del efecto de la dieta en el tamaño de las larvas y otros efectos como la duración del período larval, la presen cia de malformaciones, etc. Por lo tanto, después del tamizado de cultivares co merciales y de los clones de la CCC con una posible resistencia, se puede afirmar que todos ellos fueron susceptibles al daño de las larvas del P. vorax.
Investigación sobre preferencia
Con respecto a la investigación sobre preferencia, no obstante que en este ex perimento se utilizó una infestación ar tificial con adultos deP. vorax, los niveles de infestación al momento de la cosecha fueron muy variables, lo que no permitió el uso del análisis estadístico. Sin embar go, cuando fueron tamizados los tubér culos recién cosechados provenientes de estas parcelas, se obtuvieron los resulta dos que se muestran en la Figura 5. Una vez más, los clones 405, 4269 y 4337 fueron tan susceptibles como el cultivar Criolla, y no mostraron entre sí diferencias estadísticamente significativas.
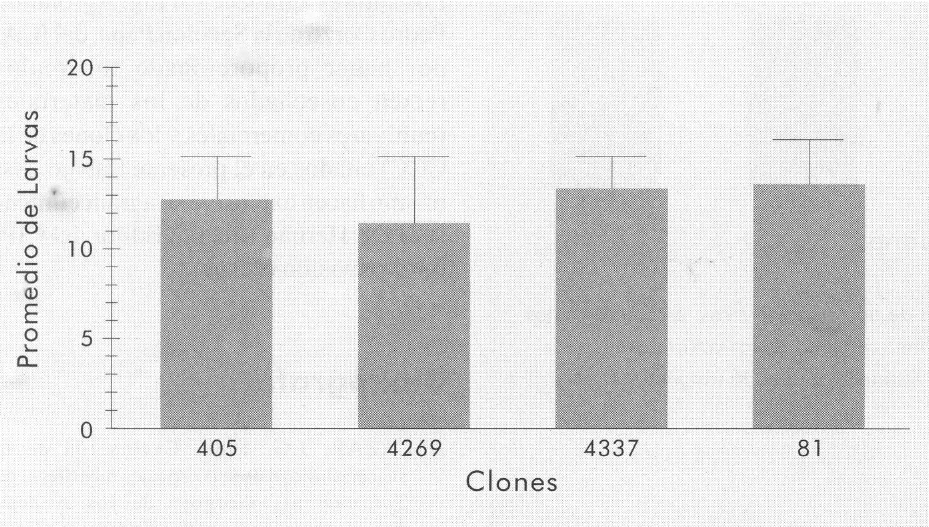
Número promedio de larvas obtenidas al final de la prueba del tamizado de tres clones con posible resistencia. Ensayo realizado con tubérculos recién cosechados. Las líneas encima de las columnas representan el valor de la desviación estándar para cada tratamiento.
Alimentación de adultos
En relación con la preferencia para su alimentación de adultos de P. vorax, los resultados de este experimento se mues tran en la Figura 6. Los adultos no mos traron preferencia por la alimentación en ninguno de los clones evaluados. La hi pótesis inicial fue que los clones con resistencia podrían tener un efecto de rechazo (repelencia) hacia los adultos de P. vorax. Sin embargo, los resultados experimentales demuestran que no hubo ningún efecto de los clones resistentes sobre ellos. Estos resultados estarían demostrando que la atracción o repelencia de estos clones hacia los adultos no tiene ninguna influencia sobre los niveles de infestación que se obtienen al momento de la cosecha.
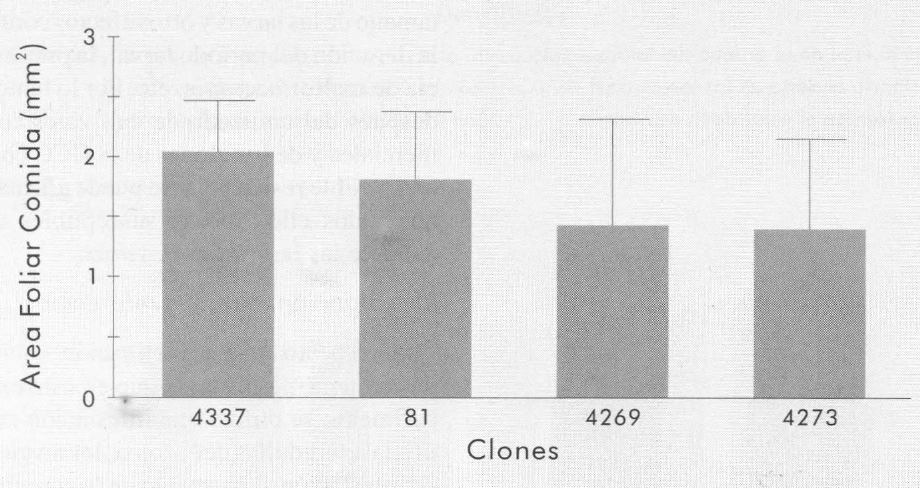
Area foliar consumida por una pareja de adultos de P. vorax, en cuatro clones de papa con posible resistencia. Las líneas encima de las columnas, representan el valor de la desviación estándar para cada tratamiento
Alimentación de larvas
Las larvas de primer instar del gusano blanco de la papa son capaces de distinguir entre dos fuentes de alimentación dife rentes. Los resultados de este experimen to se muestran en la Figura 7. Las larvas prefirieron los tubérculos del cultivar Criolla, con diferencias significativas (P =0,01), versus los tubérculos de Parda Pastusa. No obstante que las larvas del primer instar tienen una movilidad muy limitada, ellas son capaces de distinguir entre dos fuentes de alimentación dife rentes.
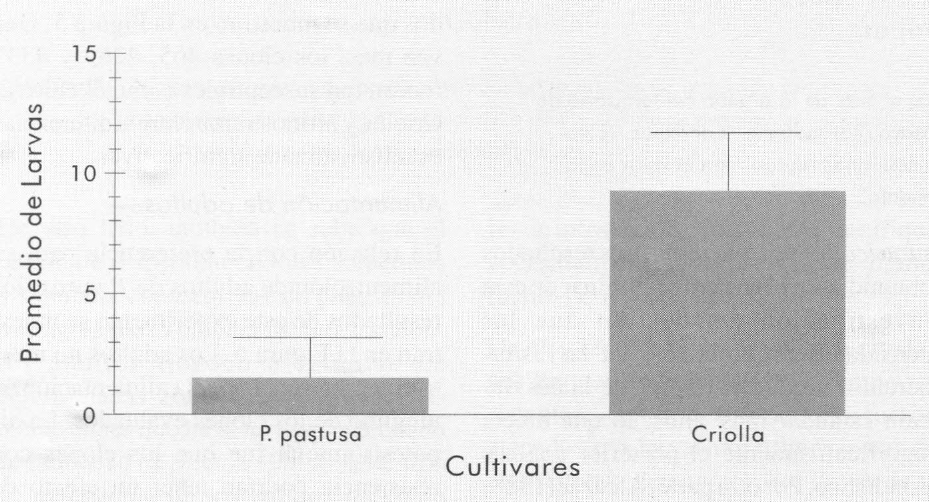
Preferencia demostrada por las larvas de primer instar de P. vorax, cuando se les ofreció como fuente de alimentación tubérculos de papa de los cultivares Parda Pastusa y Criolla. Las líneas encima de las columnas representan el valor estándar para cada tratamiento
Lo que actualmente no se conoce es cómo las larvas discriminan entre uno y otro tipo de papa. Podría ser que las larvas, en el tránsito de la superficie del suelo hacia los tubérculos, reconocen el aroma de las dos fuentes potenciales de alimentación e inmediatamente se dirigen hacia la más adecuada. O podría ser que las larvas reconocen los tubérculos por contacto (probablemente por quimioreceptores ubicados en apéndices de la boca) al momento de iniciar la penetración.
De la presente investigación no surgen indicios de la existencia de resistencia genética contra el gusano blanco de la papa, entre los materiales tamizados hasta la fecha. Sin embargo, trabajos en ejecu ción bajo el auspicio de la Universidad Nacional de Colombia, el ICA y el CIP, indican que existen fuentes de resistencia para esta plaga en híbridos de S. polyade nium generados por Estrada, en el Pro yecto de Investigación y de Producción de tubérculo-semilla de categoría prebá sica de papa en Bolivia (PROINPA) (Trillos 2 ).
Trillos, 1993. Comunicación personal. C.I. «La Selva», ICA, Rionegro (Ant.).
Footnotes
Agradecimientos
Los autores agradecen al Ing. Agrónomo Pedro Corzo de la Sección Papa, del ICA, por haber proporcionado tubérculos recién cosechados de los materiales (cultivares comerciales y los clones de la CCC) citados en el presente trabajo. Así mismo hacen extensivo su agradecimien to al Dr. Hernán Rincón, Editor del CIP, por la revisión efectuada.