
Abstract

“Lactobacilli are the most abundant bacteria present in normal vaginas of women of child-bearing age.”
In the last decade, there have been quite a lot of scientific advances reported regarding the immunology and pathophysiology of human vaginal microbiota. As a result, physicians have a more comprehensive understanding of the role that lactobacilli exert in the vaginal milieu and their interactions with vaginal mucosa, pathogenic bacteria and fungi. This commentary addresses those advances in vaginal microflora, vaginal epithelial cells, immunity, bacterial vaginosis (BV) and candidiasis. Additionally, recent reports regarding lactobacilli biofilm formation will be discussed. Last, an assessment of future implications is presented.
Vaginal microflora
Lactobacilli are the most abundant bacteria present in normal vaginas of women of child-bearing age (with some exceptions due to race, geographic location and ethnic groups) (Figure 1). They are mainly derived from intestinal microbiota. The significance of lactobacilli presence for a healthy vaginal milieu has been recognized ever since Albert S Döderlein discovered a vaginal bacillus that he named Döderlein, in 1892. This bacillus was subsequently renamed as Lactobacillus [1]. Döderlein claimed that, in normal vaginal secretions, the combination of Döderlein's bacilli and acidity was essential for keeping the vagina free of pathogenic bacteria. This fundamental concept still continues to be utilized today.

Normal fresh mount microscopic preparation.
The vaginal microflora continuously go through modifications regarding the number and type of bacteria present, almost daily, in response to exogenous and endogenous factors [2]. These factors include the menstrual cycle phases, pregnancy, sexual intercourse, vaginal hygiene and medications (contraceptives, antibiotics and immune suppressants). Alterations occurring in the vaginal environment may increase or decrease the selective advantages for specific microorganisms.
Lactobacilli are obligate homofermenters – glucose derived from vaginal epithelial cells is used to produce lactic acid. The lactobacilli most commonly found in women of child-bearing age are Lactobacillus crispatus, Lactobacillus gasseri, Lactobacillus iners and Lactobacillus jensenii [2]. Healthier vaginas harbor mostly L. crispatus and L. jensenii.
The majority of lactobacilli have a small genome of approximately 1.8–2 Mb. They have lost several genes that used to code specific biochemical actions. This loss has been balanced by the gain of many other significant genes. Some of these genes were acquired by the mechanism of horizontal gene transfer through bacterial conjugation, while others were acquired by phage infection. Quite a few of these gained genes have supported protein transporter systems that have facilitated lactobacilli to attain vaginal colonization and adaptation [3].
Lactobacilli functions
The main role of lactobacilli is to maintain a hale and hearty balanced ecological vaginal milieu with a pH range between 3.8 and 4.4. This acidic pH is responsible for inhibiting the growth of most pathogenic bacteria.
Lactobacilli put forth this inhibition by a number of mechanisms, including the production of hydrogen peroxide, lactic acid and ribosomal-produced antimicrobial peptides (bacteriocins). Lactic acid potentiates both the activity of bacteriocins and that of hydrogen peroxide [4]. The antimicrobial activity of bacteriocins includes cell membrane permeabilization with ATR amino acids and ion efflux, and transmembrane potential and pH gradient depletion. It is worth mentioning that bacteriocins express host cell immunity and do not have hemolytic or cytotoxic activity.
Lactobacilli-specific vaginotropism is facilitated by the presence of pili that act as ligands for attachment to glycolipid receptors present on vaginal epithelia cells. Additionally, lactobacilli compete for binding to receptors on vagina epithelial cells (VEC), in effect impeding pathologic microorganisms from adhering to these cells [5]. Supplementary bactericidal actions of lactobacilli products include: lactic acid, which is a potent inhibitor of Neisseria gonorrhoeae; and hydrogen peroxide, which suppresses the growth of Gram-negative bacteria, Gram-positive facultative and obligate anaerobe bacteria, including Escherichia coli, Gardnerella vaginalis and Mobiluncus species. Moreover, a recently identified bacteriocin, Lactocin 160A, specifically targets the cytoplasmic membrane of G. vaginalis [6]. Furthermore, certain bacteriocins seem to be protective against an infection of HIV, while others appear to induce membrane permeabilization on Gram-negative bacteria. Lactobacillus crispatus is specifically recognized to be a hydrogen peroxide producer. Hydrogen peroxide is involved with host myeloperoxidase and chloride ions in the formation of potent toxic oxidants that are bactericidal [6].
All of the above described actions further help lactobacilli to establish itself in the vagina and guarantee it long-term permanence to thrive with minimal competition.
Vaginal immunity
Vaginal immunity differs from systemic immunity. Reproductive tract immune cells safeguard against bacterial pathogens in the vagina and cervix, and contribute to establishing immune tolerance for sperm and embryo/fetus in the uterus. The systemic immune system is considerably influenced by sexual hormones, though leukocytes present in the reproductive tract do not hold estrogen and/or progesterone receptors [7]. The innate immune system recognizes molecular patterns connected with pathogens (pathogen-associated molecular pattern). When a pathogen-associated molecular pattern is recognized in the vagina, it activates the systemic immune system (lymphocytes T and B) causing the release of proinflammatory cytokines and the start of the cellular and humoral immunity. The innate immune system activation occurs rapidly, although several days are required for the acquired immunity to also become functional [1].
Several of these factors are active in the vagina, including soluble components such as mannose-binding lectin (MBL), and complement components and membrane-associated components called as Toll-like receptors (TLRs). MBL is an antimicrobial protein synthesized by the liver and is present in the vagina. MBL binds to bacterial mannose-containing polysaccharide surfaces causing complement activation and bacteriolysis. Women deficient in MBL are more susceptible to recurrent Candida albicans infections [2]. Additionally, the vagina contains defensins – molecules with nonspecific antimicrobial activity. Moreover, in the vagina, secretory leukocyte protease inhibitor (inhibits proteases, destroys Gram-positive and Gram-negative bacteria and blocks HIV) has been detected. Vaginal levels of this protein are reduced in patients with BV. Last, nitric oxide and phagocytic cells are also present in the vaginal milieu [1].
Vaginal epithelial cells
As we have seen, VECs play a very important role in vaginal immunity. A newly published manuscript regarding vaginal immunity describes the vaginal epithelium in women of child-bearing age as being comprised of loosely linked cells packed with glycogen. VECs are penetrable by bacteria and viruses in addition to molecular and cellular mediators of immune defense. Hence, Anderson et al. proposed that VECs provide an exclusive microenvironment that preserves vaginal health by nurturing endogenous lactobacilli while holding acquired and innate immunity mediators [8].
VECs hold, in their surface, complex TLRs. Up to recently, 11 of these TLRs have been described: TLR1 and TLR2 distinguish, in Gram-positive bacteria, their lipoproteins and peptidoglycans. TLR3 identifies, in actively replicating viruses, their double DNA chain. TLR4 recognizes liposaccharides in Gram-negative bacteria. TLR5 responds to flagellins found in flagellated bacteria. TLR9 discriminates DNA sequences that contain the CpG dinucleotide exclusively in the nonmethylated state (the DNA sequence in humans is highly methylated). Additionally, VECs release positively charged peptides that speedily bind to negatively charged surfaces of the pathogenic bacteria, disrupting the bacterial membrane and causing bacteriolysis [5].
VECs are likewise the first barrier against infections by C. albicans. These cells express different pattern recognition receptors by means of dectin-1, lectin like-receptor families and several of the TLRs [9]. Dectin-1 acting together with TLR2 and TLR4 induces not only cytokine production, but also autonomously induces IL-17, IL-10 and IL-1 [10]. Dectin-1 is also responsible for the activation of two other pathways (Src–Syk–CARD9 and NF-κB) with the final outcome of transcribing very potent antifungal cytokines including IL-1β, IL-6 and IL-23 β-glucans [11]. Additionally, CARD9 contributes to signaling dectin-2 and macrophage-inducible C-type lectin, which are known to identify Candida mannans and could help to clarify the extensive range of fungal infections in women [12]. Moreover, the mannose receptor and TLR4 distinguish Candida cell wall mannans, and TLR2 discriminates phospholipomannan and cooperates with the βGR in cytokine stimulation production [13–15].
Identification of 1- and 3-linked β-glucans by dectin-1 has been described as one of the key pathways for fungal recognition. As a matter of fact, mice which are deficient in dectin-1 have a bigger predisposition to C. albicans infections [16].
Dectin-1 intensifies TLR2- and TLR4-induced cytokine production in human cells, resulting in TNF production. Dectin-1 also is responsible for signaling induction in the production of IL-17, IL-6 and IL-10 (via a spleen tyrosine kinase-dependent pathway) [16,17].
In a study by Underhill et al., the discovery of the dectin-1 polymorphism mutation in all tested women of European and African descent has fostered two interesting hypotheses. First, it proposes that this mutation most likely appeared >60,000 years ago, earlier than when modern humans separated in the late Paleolithic [18]. This hypothesis is sustained by the discovery that the site of the haplotypes encompassing the Tyr238X mutation was closely located to the ancient haplotype. Second, it also suggests that the high polymorphism prevalence in these individuals could indicate a significant genetic predisposition for mucosal fungal infection. As soon as proinflammatory cytokines are stimulated by IFN-γ or TNF-α, macrophages discharge nitric oxide, which is recognized for its cytotoxic fungal effects [19]. Additionally, VECs produce HBD-1 and HBD-2, two human hormonally regulated defensins. When HBD-2 is reduced, it increases the susceptibility to infection [1–2,5]. Naglik et al. defined a vaginal epithelial model in which cells recognize C. albicans, although only invading hyphal cell wall moieties were able to rapidly complete cytokine activation and secretion [20].
Heat shock proteins (HSPs) are essential for life preservation. They support cell survival in varied extreme ecological conditions. They are almost pristinely evolved proteins that are present in all living creatures. HSP-70 kDa (HSP70) – synthesized in reaction to inflammation and infection – was recently documented in the vagina. Intracellular HSP70 binds other proteins avoiding their degradation. Extracellular HSP70 binds TLR to pathogens and induces nitric acid release, stimulating the immune response [2].
Other researchers have confirmed that VECs inhibit C. albicans growth [21–23]. This specifically direct antifungal activity is based on a simple epithelial cell to fungal interaction. It is disparagingly reduced in women with Recurrent Vulvo Vaginal Candidiasis (RVVC), therefore, it causes an uncontrolled development of fungi in these patients [21–24].
Vaginal antibodies
In the complicated vaginal immune milieu, antibodies producing B-lymphocytes are also locally present; they produce IgG and IgA. These antibodies are transudate from the systemic circulation and bind to pathogens provoking death by a complement-dependent mechanism or opsonization [8]. They are important speedy mechanism for fighting pathogenic microorganisms without the need for systemic immune system response. This local complex vaginal immune system is under hormonal control, which allows vaginal protection when the adaptive immunity is downregulated by the sexual hormones essential for reproduction [4].
Bacterial vaginosis
BV has been recognized as a chronic condition associated with a depletion of lactobacilli and a distinctive overgrowth of not only G. vaginalis and Atopobium vaginae but also other Gram-negative and Gram-positive anaerobes (Figure 2, clue cell) [24]. Among the pathogenic bacteria reported in BV are: Coriobacterium, Bacteroides, Veillonella, Ruminococcus, Streptococcus, Prevotella, Megasphera, Leptotrichia, Clostridium-like species, and a few others. All of them support each other by their own synergistic properties [25].
Enzymes produced by these pathogen bacteria comprise glycosidases and sialidases. These enzymes are responsible for the characteristic low viscosity of vaginal discharge with the fishy odor of BV. The enzymatic activities of G. vaginalis and the other BV bacteria are halted during antibiotic therapy, but slowly restart after therapy is terminated [24,25]. It is interesting to note that G. vaginalis is also present in most women of child-bearing age with normal vaginal flora or without BV [25].
Additionally, G. vaginalis strains obtained from women with BV demonstrate a different genomic pattern than those obtained from patients without BV in regards to adherence, aggregation and cytolysin productions [26,27]. At wet mount preparations, BV presents with typical clue cells. Vestraelen et al. identified these clue cells as having been desquamated squamous epithelial cells derived from the biofilm [28].
Box 1. Bacterial vaginosis therapy Center for Disease Control 2015 guidelines
Oral metronidazole 500 mg twice a day for 7 days or
Topical 0.75%, 5 g once a day for 5 days or
Clindamycin 2%, 5 g vaginally for 7 days
Alternatives
Tinidazole 2 g orally daily for 2 days
Tinidazole 1 g orally daily for 5 days
Clindamycin 300 mg orally twice daily for 7 days
Clindamycin 100 mg ovules vaginally for 3 days
Relapse
Metronidazole, extended oral followed by vaginal maintenance therapy twice weekly for 4–6 months
Swidsinski et al. reported, by means of the FISH technique, that clue cells adhered in two different patterns: cohesive (in groups of highly concentrated bacteria) and dispersed (solitary and/or intermixed with other bacteria) [29].
One of the unanswered vaginal mysteries is the absence of noticeable inflammation in patients with BV. The reasoning being that in BV fluid there is considerable overgrowth of Gram-negative and Gram-positive anaerobes; also several proinflammatory cytokines like IL-6 and IL-8 are present [30,31]. Since the majority of vaginal microbiota originates from the gut microbiota, the lack of inflammation could be attributed to immune tolerance acquired through evolution [32–35]. Other contributing factors associated with the absence of inflammation include the presence of short-chain fatty acids. These fatty acids are the by-products of anaerobes and they modulate immune responses. They inhibit proinflammatory cytokine production and immune cell migration, and they induce several cells apoptosis including neutrophils [36].
Recently, Vitali et al. reported on the vaginal microbiome and metabolome from patients with BV and healthy controls using quantitative PCR and proton nuclear magnetic resonance (1H-NMR). The authors correlated the clinical condition with vaginal microbiome bacterial by principal component analysis. They concluded that L. crispatus was prevalent in the healthy vagina, while Prevotella, Atopobium and Mycoplasma hominis were dominant in BV patients [37]. Through 1H-NMR analysis, the researchers linked BV with nicotinate, malonate and acetate while maltose, kynurenine, and NAD+ were mainly found in the healthy vaginas. Therefore, the authors proposed a combined molecular approach for the diagnosis of BV based on quantitative PCR and (1)H-NMR [37].
Bacterial vaginosis therapy
The 2015 Center for Disease Control guidelines for bacterial vaginosis therapy are summarized in Box 1.
Biofilm
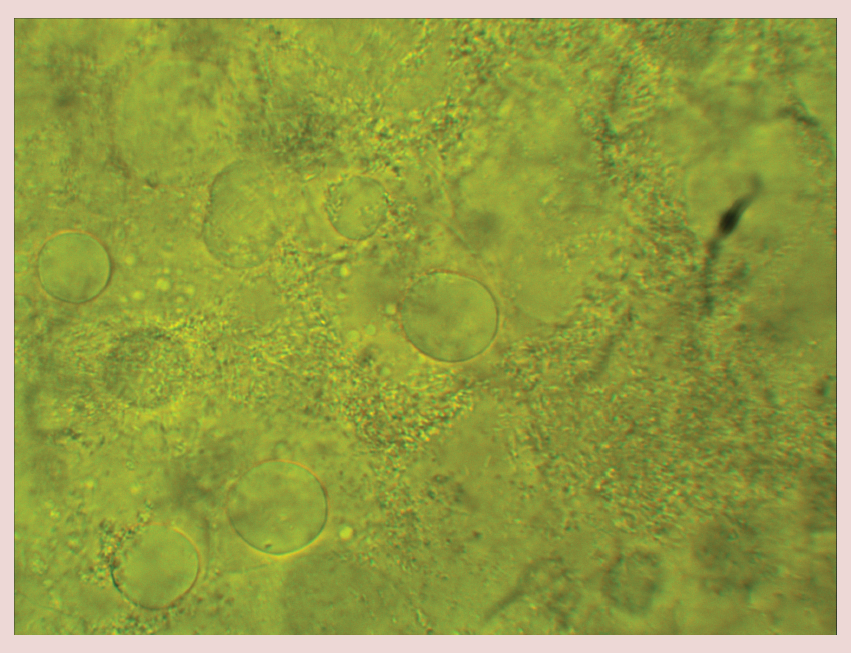
An additional important activity of lactobacillii is to produce biofilm. Biofilms are complex biological structures that facilitate bacteria in adhering to one another, to surfaces, and/or to interfaces, enabling the bacteria to thrive and protect themselves. Biofilm formation by lactobacillii is a refined process involving aggregation and recognition of surface-related stimuli that allows them to adhere to a surface. There, they are able to produce and stockpile the biofilm matrix. Ventolini et al. recently reported the in vivo formation of such biofilm by L. jensenii in fresh wet-mount microscopic preparations (Figure 3) [38].

Understanding of the conditions involved in biofilm formation has significantly progressed. The in vivo biofilm formation by lactobacillii has recently been reported [38]. Gardnerella vaginalis and A. vaginae play a major role in the development of vaginal biofilm. The production of biofilm by both bacteria appears to be responsible for the continuance and high recurrence rates of BV [39,40]. Gardnerella vaginalis produces an adherent biofilm which is still present even after recommended metronidazole therapy is completed [28,31]. Moreover, in vitro experiments using G. vaginalis biofilms have concluded that gardnerella can tolerate five-times the normal vaginal hydrogen peroxide concentration and four- to eight-times the normal lactic acid concentration [41–43].
Swidsinski et al. reported that a compact biofilm in layers was attached to 50% of epithelial cells' surfaces in 90% of patients with BV, compared with only 10% in uninfected controls (p < 0.001) [28].
Frequently, atopobium has been reported to be found enclosed in biofilm associated with BV. Atopobium is resistant to metronidazole, and biofilms typically repel antibiotic treatments [27,32,39].
Molecular biology studies regarding the type, number and relative proportion of vaginal bacteria colonization have revealed that there is a great difference between patients with BV and unaffected patients. It seems that this difference most likely reflects each individual woman's anogenital colonization [27,31,42,44–45].
Vaginal candidiasis
Vaginal candidiasis is a highly prevalent infection found all throughout the world that has dramatically increased in recent years. The primary species involved is Candida, and C. albicans is by far the most commonly reported fungus (Figure 4) [46,47].

Fresh wet mount preparation showing pseudohyphae with chlamydospores.
The non-albicans candida's species that are also implicated in vaginal fungal infections include Candida glabrata, Candida krusei, Candida tropicalis, Candida stelloidea and Candida dubliniensis. In rare cases, Saccharomyces cerevisiae and Aspergillus nigrans have also been reported [48].
Excessive vulvovaginal care with products including soaps, bubble baths, douches and creams may be contributing to the disruption of the vaginal milieu and predispose to fungal infections. Likewise, numerous risk factors have been stated to be involved in recurrent vaginal candidiasis. Among them are local vaginal immune defense deficiencies, allergic factors, gene polymorphisms, glucose serum levels, antibiotic therapy, psychosocial stress and estrogens [49,50].
Genetic factors in both the fungus and in the host must be significant to comprehend who gets an infection and why. Very little is known about detailed human genetic predispositions to fungi.
Smeekens et al. reported that genetic dissimilarities in the host are likely responsible for the susceptibility to acquire fungal infections. For instance, severe infections are mostly linked to a single gene of immune deficiency [49]. And recently, such gene deficiencies including STAT1, STAT3 and CARD9 have been reported [14,51]. Mutations in the gene encoding transcription factor STAT3 damage several pathways, including the generation of IL17-committed T lymphocytes (Th17 cells). Th17 cells produce IL-22, which synergizes with IL-17 in the epithelial synthesis of defensins. STAT3 also signals itself, IL-17 and IL-22. [12] On the other hand, gene polymorphisms related to immune deficiencies have typically been linked with recurrent vulvovaginal candidiasis [51].
Ferwerda et al. described a family in which four women with either recurrent vulvovaginal candidiasis or onychomycosis displayed the early-stop-codon mutation Tyr238X in their βGR. Their dectin-1 mutated forms were not well expressed, did not mediate β-glucan binding, and led to defective production of IL-17, TNF and IL-6 after they were stimulated with β-glucan or C. albicans [52]. However, fungal killing and phagocytosis were normal in these women. According to the authors, dectin-1 deficiency was not related to invasive fungal infections but had a definite role in human mucosal antifungal defense [52]. In conclusion, Ferwerda et al. proposed that mucosal candidiasis, which is generally seen in women with T-cell-mediated immunity deficiency, (Th17) may also be caused by a genetic defect of the βGR. Therefore, dectin-1 may be specific for mucosal antifungal defense [52].
Dectin-1 is the cell-surface receptor for β-glucan, a major component of the budding yeast cell wall, and its signaling travels through a series of molecules, including CARD9, leading to the activation of NF-κB [12].
It is still unknown if the primary mechanism in severe candidiasis involves compromised dectin-1 signaling at the epithelial level or epithelium compromised leukocyte activation mediated by IL-17. A comparable defect has been reported in women with defective CARD9. The CARD9 mutation is rare but its infrequency is proportionate with its severity. CARD9 is involved in other cell-surface and intracellular signals, among them are p38 MAPK and Jun N-terminal kinase pathways [12]. Pinke et al. recently reported that mast cells were observed to phagocytose and produce harmful nitric oxide to C. albicans by way of TLR2/dectin-1 [53]. These current reports explain the severity of infections by Candida as compared with isolated dectin-1 deficiency [14]. The 2015 guidelines from the Center for Disease Control USA for the treatment of vaginal candidiasis are summarized in Box 2.
Genetic components play a significant role in the susceptibility to RVVC; among them, a number of polymorphisms in IL-4, NALP3 and MBL have been identified [52]. For instance, NLRP3 defects more often found in RVVC patients have facilitated the identification of the inflammasome as a critical constituent, although there may be many more involved [51].
Candidiasis therapy
Candidiasis is not a sexually transmitted disease. Chronic vulvar/vaginal pruritus, clumpy white vaginal discharge, burning, irritation, discomfort and/or pain are common complaints from women with RVVC [54].
RVVC is a very important theme that is challenging to understand and to treat. The vast majority of women diagnosed with RVVC develop the infection without a history of any identifiable risk factors. RVVC is correlated with vaginal immune deficiency. Hence, the successful treatment of an acute episode does not prevent recurrences. This is the reason why an acute episode treatment should be followed by a weekly preventive treatment for at least 6 months and in some cases, many years [54].
Conclusion
The most important scientific advances regarding the immunology and pathophysiology of human vaginal microflora, including BV and Candidiasis, were addressed.
Box 2. Vaginal candidiasis therapy Center for Disease Control 2015 guidelines
Prescription intravaginal agents
Butoconazole 2% cream (single-dose bioadhesive product), 5 g iv. in a single application or
Terconazole 0.4% cream 5 g iv. daily for 7 days or
Terconazole 0.8% cream 5 g iv. daily for 3 days or
Terconazole 80 mg vaginal suppository, one suppository daily for 3 days
Over-the-counter intravaginal agents
Clotrimazole 1% cream 5 g iv. daily for 7–14 days or
Clotrimazole 2% cream 5 g iv. daily for 3 days or
Miconazole 2% cream 5 g iv. daily for 7 days or
Miconazole 4% cream 5 g iv. daily for 3 days or
Miconazole 100 mg vaginal suppository, one suppository daily for 7 days or
Miconazole 200 mg vaginal suppository, one suppository for 3 days or
Miconazole 1200 mg vaginal suppository, one suppository for 1 day or
Tioconazole 6.5% ointment 5 g iv. in a single application
Oral agent
Fluconazole 150 mg orally in a single dose
Recurrent infections
7–14 days of topical therapy or
Oral fluconazole: 100, 150 or 200-mg dose every third day for a total of three doses
Oral fluconazole (i.e., 100, 150 or 200-mg dose) weekly for 6 months is the first-line maintenance regimen
iv.: Intravenous.
Future perspective
The pathogenesis of BV is still under investigation and an animal model has not been identified yet. BV presents several more challenges, among them are: the need to develop more effective treatments, to find ways to re-establish the vaginal milieu with lactobacilli after treatment, and to tackle recurrences [36,55].
We now have a better knowledge of the pathophysiological mechanisms essential for vaginal mucosa antifungal defense. Preliminary genetic susceptibility association studies should be extended to larger multiethnic cohorts of RVVC patients.
Additionally, it is necessary to comprehend the intricate function that probiotics provide with regard to fungal infection prevention, treatment and recurrences [55,56].
A better understanding of the role that lactobacillii biofilms exert in the vaginal milieu could provide an additional natural biological weapon on the fight against intrusive pathogenic bacteria and fungi in the vagina.
Moreover, lactobacilli products like bacteriocins and biofilm could be the alternative to antibiotics, for not only to maintain a healthy vaginal milieu but also to treat recurrent vaginal infections. As well, the role that lactobacilli play in term and preterm labor mechanisms is still in early investigation [57,58]. All these important research challenges may finally lead to an enhanced understanding of the pathogenesis of BV and RVVC as a result to propose innovative therapies.
Footnotes
Acknowledgements
The author would like to thank Melissa Waggoner, Senior Editor, for their assistance in English editing.
G Ventolini has no relevant affiliations or financial involvement with any organization or entity with a financial interest in or financial conflict with the subject matter or materials discussed in the manuscript. This includes employment, consultancies, honoraria, stock ownership or options, expert testimony, grants or patents received or pending, or royalties.
No writing assistance was utilized in the production of this manuscript.