
Abstract
Normal menstruation is an inflammatory process, where the endometrial concentrations and functions of several leukocyte types can change greatly through the menstrual cycle, especially during the premenstrual and menstrual phases. These leukocytes probably have a range of functions related to mucosal protection, decidualization, embryo implantation, and the process of menstrual tissue breakdown, repair and remodeling. Some of these leukocyte changes are apparently linked to changes in the pattern of circulating leukocytes. Many immune cells have been identified in the endometrium, and those with most relevance to the processes of menstruation include uterine natural killer cells, macrophages, mast cells, neutrophils, dendritic cells and Tregs. A range of disturbances in endometrial immune cell numbers, distributions and functions, and in a range of different inflammatory and other mediators, have been identified in women with heavy menstrual bleeding or endometriosis. Sufficient evidence exists to implicate these immune changes in some of the functional disturbances and symptoms identified in these women. This field is greatly under-researched, and ripe for the wider application of modern molecular and cellular techniques in human and animal model studies.
Keywords
Menstruation is a most unusual physiological process that only occurs in a very small number of animal species – humans, great apes, most old world monkeys and, apparently, in some bats, elephant shrews and rock hyraxes. Its purpose is mysterious, although it appears to be related to the type of hemochorial placentation and the involvement of spiral arterioles. This placentation requires differentiation of the decidual tissues to such a degree that tissue necrosis and shedding occur, rather than resorption and remodeling. The spiral arterioles contribute through intense vasoconstriction resulting in localized tissue ischemia, matrix metalloproteinase (MMP) release, tissue fragmentation, necrosis and shedding.
Menstruation is a highly regulated process that follows a precise sequence of endocrine changes beginning with an unopposed stimulation of endometrial glandular and stromal proliferation by ovarian follicular estradiol (the endometrial proliferative phase) [1]. This is followed by luteal progesterone stimulation of endometrial secretory change proceeding on to predecidual change at the time when embryo implantation may be occurring (the endometrial secretory phase). As luteolysis begins, serum estradiol and progesterone levels begin to fall, and these declines trigger a further series of precisely regulated processes resulting in the onset of superficial endometrial tissue breakdown. These changes occur during a very short 48-h period before the appearance of overt menstrual bleeding (the premenstrual phase).
It used to be thought that menstruation was just an endocrine process driven by changes in hormone secretion from the ovary, but it is becoming clear that it is actually much more complex. It has been recognized for many decades that there is a ‘leukocyte infiltration’ into the endometrium premenstrually [2–5]; however, it was Finn who linked a number of pieces of evidence to recognize that menstruation is also a complex ‘inflammatory’ process [6]. This inflammatory process involves a series of major changes in endometrial composition and in the functions of several different leukocyte types through the menstrual cycle, to such an extent that approximately 40% of all stromal cells in the late ‘luteal/secretory/predecidual’ phase are leukocytes.
Endometrial immune response during the normal menstrual cycle
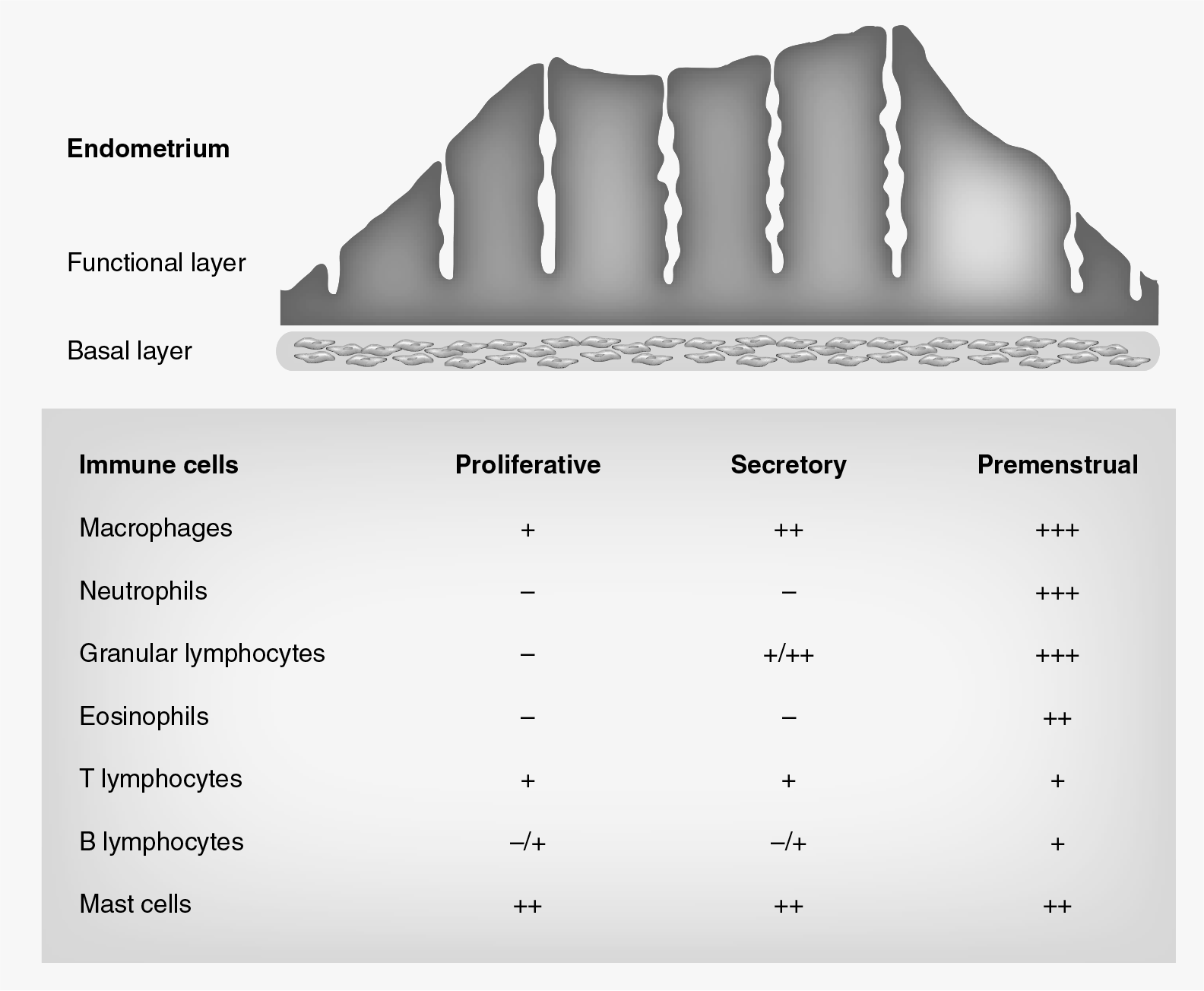
Uterine immune cell populations undergo substantial changes throughout the normal menstrual cycle. These leukocytes provide immune protection for the uterine mucosal surfaces and play crucial roles in endometrial remodeling, decidualization, and embryonic implantation, and in the absence of pregnancy, facilitate the process of menstruation [7,8]. Immune cells, including macrophages, neutrophils and natural killer (NK) cells, have been implicated in this response and it has been shown that these cells are recruited into the eutopic endometrium during the secretory phase of the cycle, in preparation for the onset of menstruation (Figure 1) [9]. However, it is becoming increasingly clear that immune cells also play important roles during other phases of the menstrual cycle. A number of distinct immune cell populations appear to be implicated in the menstruation process itself, while others play important roles in endometrial clearance following the menstrual breakdown. The following sections explore the role of the immune system in preparing the uterus for the process of menstruation, as well as in facilitating subsequent clearance of the uterine cavity from menstrual debris.
Recruitment of immune cell populations during the menstrual cycle.
Preparation for onset of menstruation
Prior to the onset of menstruation, falling levels of estrogen and progesterone, in absence of pregnancy, are believed to trigger recruitment and in situ expansion of leukocyte populations. Increased numbers of macrophages, eosinophils, neutrophils, granular lymphocytes, B and T lymphocytes, as well as mast cells, are evident in the late secretory endometrium and, at this time of the cycle, these cells comprise up to 40% of the total cells within the stromal compartment (Figure 1) [2]. Premenstrual withdrawal of progesterone increases the expression of inflammatory mediators, including IL-8 and MCP1 [10], which are believed to drive endometrial leukocyte recruitment during this time of the cycle [4,5,11,12]. The immunological responses occurring premenstrually are largely innate in nature [10]. Marked increases in the numbers of macrophages and neutrophils are evident during this time of the cycle [13], and these cells are a rich source of natural antimicrobial proteins, including defensins and whey acid protein motif proteins, which play key roles in ensuring microbial protection at the time when the epithelial barrier is disrupted [10,14].
Other cell populations that play crucial roles prior to menstruation are mast cells. While mast cell numbers remain relatively constant throughout the menstrual cycle, elevated expression of extracellular tryptase in the late secretory phase is suggestive of increased mast cell activation during this time [2]. Tryptase and other mediators released by mast cells (histamines, cytokines and growth factors) induce tissue edema and play roles in degradation of extracellular matrix through activation of proteolytic enzymes and MMPs [2]. MMPs are activated by a variety of enzymes and leukocyte proteases, which leads to activation of an MMP cascade and subsequent degradation of extracellular matrix.
Process of menstruation
Upregulated mast cell and MMP activation persists into the menstrual phase of the cycle [10]. MMPs, in particular, facilitate the process of endometrial breakdown [15]; however, most of the breakdown has occurred prior to the menstrual shedding and, at this time of the cycle, activation of endometrial repair mechanisms is well established.
At menstruation, when shed endometrial fragments and debris fill the uterine cavity, specific immunological changes are believed to be initiated with regard to the numbers and functions of uterine immune cell subsets. Dendritic cells (DCs) and other antigen-presenting cells (APCs), which function to capture antigens and initiate specific immunological responses, almost certainly facilitate clearance of the uterine cavity at the time of menstruation [16,17]. DCs undergo significant variations during the normal cycle and their numbers peak at the time of menstruation. Both immature and mature DCs are elevated during the menstrual phase [16]. Elevated numbers of DCs during menstruation suggests that these cells function to capture shed endometrial fragments and to activate subsequent immunological reactions that would facilitate endometrial clearance.
Macrophages, although not as specific as DCs, are also capable of antigen presentation. A significant increase in macrophage cell numbers is evident during the normal midmenstrual phase and, at this time, these cells are believed to facilitate endometrial clearance through antigen presentation and through initiation of apoptosis of shed fragments [18].
Under normal conditions, in response to exposure to certain antigens or other inflammatory signals, immature DCs undergo maturation and travel to the lymph nodes (LNs) via the afferent lymphatics, where antigens are presented to T cells [19]. Further supporting the role of efficient antigen presentation at the time of menstruation is the finding of elevated CD10+ endometrial stromal cells in the uterine-draining LNs during this time of the cycle [20]. When the endometrium breaks down and is shed, endometrial lymphatics are exposed to the uterine cavity, thereby providing the opportunity for endometrial products (cells and fragments) to be channeled into the regional lymphoid tissue for destruction. The inflammatory processes of menstruation are also associated with increased numbers of T cells, macrophages, NK cells, B cells and plasma cells in uterine-draining LNs during menstruation. Increased presence of both endometrial stromal cells and immune cells in regional LNs suggest that the process of menstruation is associated with specific adaptive immunological activations, which are likely to occur with every normal cycle. These concepts are summarized in

It can thus be postulated that APCs are the key immune cell subset that govern the removal of menstrual debris following endometrial breakdown. Subsequently, these immune cells are likely to be facilitating the processes of efficient endometrial repair and regeneration following normal menstruation.
Postmenstruation
Although endometrial repair mechanisms are initiated early following endometrial breakdown, lymphocyte activation becomes particularly apparent during the proliferative phase of the cycle. During the early- to mid-proliferative phase of the cycle, CD8+ cytotoxic T cells are highly activated and are believed to function in ensuring immunological surveillance [21].
It can be hypothesized that elevated cytotoxic activity results directly from efficient antigen capture and subsequent T-cell activation that occurs during menstruation. Thus, this activation may be crucial for finalizing the clearance of residual debris and potential antigens, which may persist within the uterine cavity. Increased cytotoxic activity may also play a role in protecting the uterine cavity from pathogens prior to implantation. Following menstruation, the uterine environment is more conducive to population by microbes given that menstrual blood may serve to provide an optimal environment for microbial proliferation and pathogenicity. Consequently, a study has shown that bacterial contamination is present during the normal menstrual cycle and is most evident during the menstrual and proliferative stages of the cycle [22].
With rising preovulatory estrogen levels prior to ovulation, the cytotoxic T-cell activity reduces abruptly. It is believed that this reduction serves to ensure successful implantation. Rising levels of Foxp3+ Tregs during this phase of the cycle are likely to be implicated in suppression of this cytotoxicity [17]. Tregs are immune cell populations that play crucial roles in controlling and suppressing a range of immunological responses. Evidence suggests that Tregs induce immunological tolerance that is required for successful embryonic implantation [23]. In the absence of implantation, the endometrium will prepare for the process of menstruation and the cycle will begin again (Figure 2).
Dysregulated immune response & menstrual symptoms
The systemic and endometrial immune responses during the menstrual cycle appear to be tightly controlled, and dysregulations in these responses may lead to menstrual symptoms and pathology. The sections below explore how dysregulations in the immune response may lead to heavy menstrual bleeding (HMB) and may contribute to development of gynecological pathologies, such as endometriosis.
Heavy menstrual bleeding
HMB is a common gynecological problem, characterized, in research terms, by menstrual blood loss of greater than 80 ml per cycle [24]. While HMB is experienced by up to 15% of menstruating women, the exact pathophysiological mechanisms that cause HMB are largely unknown. However, it has been demonstrated that a number of disturbances at the molecular level within the endometrium may contribute to prolonged or excessive bleeding. However, no investigator has studied more than one mechanism at the same time in the same group of women. While the immune response is tightly regulated during the normal menstruation, some evidence suggests that immunological disturbances may contribute to abnormal uterine bleeding, including HMB.
With the withdrawal of progesterone during normal luteolysis and prior to the onset of normal menstruation, large numbers of immune cells are channeled into the endometrium where they secrete and promote the release of a range of inflammatory mediators. Inflammatory and other mediators, secreted by endometrial immune cell populations, play key roles in endometrial breakdown, regeneration and repair. A range of these mediators are disturbed in women with HMB. In 2006, Malik et al. demonstrated that endometrial VEGF-A, which plays a role in neovascularization, is significantly reduced in women with HMB [25]. VEGF-A levels are normally elevated during menstrual and proliferative phases of the cycle. During this time, VEGF-A induces endothelial cell expression of PDGF and heparin-binding EGF, both of which promote migration and differentiation of vascular smooth muscle cells. Furthermore, VEGF-A has the potential to activate tissue factor – a major cellular initiator of coagulation [25]. In the endometrium, various immune cells, particularly macrophages, give rise to VEGF-A [26]. Reduced levels of VEGF-A in women with HMB may account for prolonged, heavy bleeding through defective vascular regeneration and endometrial repair.
Other important endometrial immunological mediators, including MMPs and TNF-α, are also altered in women with HMB. MMP-2 and MMP-9 are significantly reduced in HMB, while TNF-α expression is significantly elevated in these women [25]. MMP-2, in particular, strongly promotes vasoconstriction through mediation of platelet aggregation [27]. It also plays a role in activating neutrophils and promoting leukocyte–endothelial cell adhesion [27]. Reduced immune cell activation and adhesion in HMB may contribute to a dysfunctional local immunological environment in which regeneration and repair mechanisms do not occur efficiently. Similarly, overproduction of inflammatory mediators, such as TNF-α, in women with HMB is likely to further delay the ability of the endometrium to repair. One of the cardinal features of inflammation is vascular dilatation, which leads to hyperemia, a process that could directly account for disturbed bleeding patterns. Furthermore, TNF-α is secreted by a large number of endometrial leukocytes, predominantly macrophages, and significantly contributes to menstrual shedding through induction of apoptosis [25]. In a mouse model, administration of TNF-α leads to endometrial hemorrhage through induction of vascular damage [28]. Through similar mechanisms, increased TNF-α secretion by the inflammatory endometrial leukocytes may contribute to excessive menstrual bleeding, as well as delay in endometrial regeneration and repair.
Furthermore, it has been postulated that migratory endometrial leukocytes may directly influence vascular permeability [29]. Macrophages and other leukocytes may lead to excessive bleeding through secretion of potent vasodilators, platelet-activating factor and prostaglandin E, while uterine NKs, through secretion of perforins, could increase vessel permeability through degradation of the endometrial vascular and cellular structures. Other immune cells, especially mast cells, are also likely to be dysregulated in HMB. Mast cell degranulation, which normally occurs premenstrually, leads to release of various preformed mediators including heparin, an anticoagulant. Increased heparin-like activity has been reported in women with HMB [30,31]. Other mast cell products, including COX-1 and −2 and prostaglandins, are also increased in the endometrium of women with HMB. Mast cells synthesize arachidonic acid, an intermediate that leads to the production of prostaglandins, prostacyclins and thromboxane. Prostaglandin production plays an integral role in the inflammatory process of normal menstruation [32,33]; however, significantly elevated levels in HMB may alter the function of surrounding endometrial leukocytes. Certainly, it has been reported that elevated levels of COXs and overexpression of prostaglandins may adversely affect the numbers and function of other leukocytes, including T-cell and DC populations [34].
Whether or not a dysregulated immunological response is a primary cause for HMB, and indeed other abnormal uterine bleeding symptoms, remains uncertain. However, large numbers of inflammatory cells that are recruited into the endometrium in preparation of menstruation play crucial roles in orchestrating a range of events that lead to endometrial breakdown, as well as subsequent regeneration and repair. In women with HMB, factors that directly promote bleeding through inflammation and vasodilatation, or through delayed or impaired vascular regeneration and anticoagulation, are likely to be implicated in the process of excessive blood loss at the time of menstruation.
Endometriosis
Endometriosis, characterized by presence of endometrial-like glands and stroma, outside the uterine cavity has been widely documented to have an inflammatory component [35,36]. Numerous studies have shown that peritoneal leukocytes and their inflammatory mediators exert local effects, creating a microenvironment that may facilitate the development and progression of endometriotic lesions. Increased endometrial aromatase activity in endometriosis enhances local estrogen biosynthesis at lesion sites and, in doing so, plays a role in supporting the proliferation and growth of these estrogen-dependent endometriotic fragments [37,38]. Macrophages, in particular, have been shown to express aromatase enzyme, which can stimulate local estradiol production at the lesion site [39], and in doing so encourage disease progression [35]. Alongside aromatase expression, immune cells at and around endometriotic lesion sites secrete a wide range of proinflammatory mediators and growth factors, which have been shown to promote survival and facilitate establishment and growth of lesions [40].
The immune system appears to be significantly disturbed not only at lesion sites, but also within the uterine cavity of women with endometriosis [35,36,41,42]. In both these situations, the demonstrated changes in immune cell numbers and functions are also strongly suggestive of effects on development and activation of local nerve fibers sufficient to influence mechanisms responsible for triggering of pain signals [43]. Not only does the aberrant immune response contribute to mechanisms of pain in endometriosis, it is also believed to contribute to infertility. Alterations in the local environment and immune function in women with endometriosis have been shown to interfere with successful embryonic implantation, leading to increased rates of implantation failure and infertility in women with endometriosis [44,45].
The role of the immune system in endometriosis is believed to be complex. Disequilibrium between neuroendocrine and immune function is believed to be at play in endometriosis [46]. Increased stress response, characterized by elevated corticotrophin-releasing hormone, and urocortin secretion in endometriosis affects the local immune environment and is believed to contribute to altered immunological profiles and immune cell function in women with endometriosis [47].
Unlike normal menstruation, the ability of endometrial APCs to capture and present antigens appears to be compromised in women with endometriosis. Ineffective targeting of shed endometrial fragments in endometriosis is suggested by the observation of suppressed DC maturation [48]. CD83+ mature DCs, which play vital roles in initiating appropriate immunological responses following antigen capture, were highly significantly reduced in numbers not only during the menstrual phase, but across all phases of the menstrual cycle in women with endometriosis. These findings suggest that components of antigen capture and/or presentation are likely to be disturbed in the endometrium from women with endometriosis, where inefficient clearance of endometrial fragments may favor establishment and progression of disease at ectopic locations.
Furthermore, in women with endometriosis, the numbers of various immune cell populations, including macrophages, uterine NK cells and immature DCs were persistently increased following menstruation (proliferative phase) [18,42,48]. This is in contrast to the normal menstrual cycle. These immune cells may reflect a prolonged attempt by the immune system following menstruation to clear the uterine cavity, subsequent to failure of antigen-presenting mechanisms. Similarly, in contrast to the normal cycle, where T-cell activation occurs during the proliferative phase, T-cell activation persists into the secretory phase in women with endometriosis. In concordance with this, increased numbers of Tregs during the secretory phase in endometriosis are likely to be crucial for immunological suppression of this overly inflamed environment [17,49]. Furthermore, a secretory rise in Tregs may directly interfere with premenstrual recruitment of immune cell populations and reduce the ability of newly recruited DCs to efficiently present antigens in women with endometriosis [50]. Overall, the observations of the endometrial immune environment in endometriosis indicate inefficient targeting of shed endometrial antigens, factors that allow survival of shed fragments and their consequent ectopic establishment as endometriotic lesions.
In addition, while the inflammatory processes of menstruation are associated with increased numbers of T cells, macrophages, NK cells, B cells and plasma cells being channeled into the uterine draining LNs, the numbers of these cell populations were consistently reduced in LNs from women with endometriosis [20]. The mechanism(s) responsible for the decline in leukocyte numbers in the LNs of women with endometriosis are unclear; however, recruitment of large numbers of immune cells to ectopic endometriotic sites is one possible explanation. Regardless of the mechanisms involved, these findings suggest that efficient immunological responses may be required to contain shed endometrial fragments within the draining-uterine LNs in order to prevent their further dissemination and establishment at ectopic sites. Such findings raise a possibility that endometriosis may be a disease in which primary dysregulations of the immune system (inherited or acquired) may contribute to disease pathophysiology.
Conclusion & future perspective
Large fluctuations in the numbers and functions of immune cells occur throughout the menstrual cycle. These fluctuations are likely to be attributed to a number of factors, including leukocyte recruitment and migration from the peripheral circulation; apoptosis and cell loss during endometrial breakdown; and cellular proliferation within the tissue [51,52].
In preparation for the process of menstruation, carefully coordinated recruitment of immune cell populations precedes the process of endometrial breakdown, during which time increased numbers of APCs are likely to be actively capturing endometrial fragments and activating cytotoxic T-cell responses (which predominate after the process of menstruation is complete). The cytotoxic responses are precisely controlled and suppressed prior to the implantation period during a normal conception cycle. Dysregulated immunological regulation may create an inflammatory environment that is likely not only to interfere with successful embryonic implantation, but also contribute to other pathologies, such as HMB and endometriosis.
Future studies should aim to better define specific day-to-day changes in the immune cell composition and function during the menstrual phase of the cycle, as well as to determine which APCs are likely to be most implicated in antigen recognition following endometrial breakdown at menstruation. In addition, studies into the functions of DC and Treg subsets should be conducted, with a focus on determining if specific endometrial antigens are present at the time of menstruation, and if so, how they might function to recruit APCs into the endometrium. It is thus a matter of priority that future studies in this area explore how the immune cell populations recognize and target shed fragments during menstruation, and how this recognition and targeting may be altered in pathological states. Such studies almost certainly require a multicenter-collaboration approach in order to ensure a larger sample size along with well-characterized clinical data. Understanding the role of the immune response in both the presence and absence of disease has considerable potential for developing better therapeutic options through the prospect of manipulating apoptosis and the mechanisms of clearance of shed endometrial fragments [53]. Therefore, the prospects of better defining the immune response during menstruation and understanding how this immune response may be modulated is knowledge that will potentially allow for earlier detection, prevention and management of menstrual and endometrial disorders.
Financial & competing interests disclosure
The authors have no relevant affiliations or financial involvement with any organization or entity with a financial interest in or financial conflict with the subject matter or materials discussed in the manuscript. This includes employment, consultancies, honoraria, stock ownership or options, expert testimony, grants or patents received or pending, or royalties.
No writing assistance was utilized in the production of this manuscript.
Executive summary
Menstruation is an inflammatory process.
Embryo implantation is an inflammatory process.
Endometrial immune cell populations undergo a sequence of well-coordinated changes during the normal menstrual cycle.
Very specific changes in leukocytes are recognized leading up to and following menstruation.
Some endometrial leukocyte changes are linked to changing patterns of circulating leukocytes.
Endometrial leukocytes have important roles in mucosal protection, decidualization, embryo implantation, menstrual tissue breakdown and endometrial repair and remodeling.
Leukocyte types involved include uterine natural killer cells, macrophages, mast cells, neutrophils, dendritic cells, eosinophils, occasionally B cells and a variety of T-cell types, especially Tregs. Tregs are probably the main controlling cell type.
Much is now known about changes in different leukocyte numbers and densities related to the menstrual process; however, little is known about their changes in function at these times.
Much less is known about the disturbances in immune cell types and functions in relation to heavy menstrual bleeding; however, there are enough fragments of evidence to support meaningful speculation. There is sufficient evidence to say that inflammatory and other mediators, secreted by endometrial immune cell populations, play key roles in endometrial breakdown, regeneration and repair. A range of these mediators are disturbed in heavy menstrual bleeding. Evidence of immune cell activation and adhesion in heavy menstrual bleeding may contribute to a dysfunctional local immunological environment in which regeneration and repair mechanisms do not function effectively. Vascular dilatation and permeability may be similarly disturbed by changes in immune cell function.
Much more is known about endometrial immune cell changes in women with endometriosis and the ways in which they may relate to the generation of pain symptoms, and to the ability of shed endometrial fragments to survive and contribute to peritoneal endometriotic lesions.
Macrophages, uterine natural killer cells, dendritic cells and Tregs may all play important roles in the disturbances of endometrial function in endometriosis.
Much remains to be learned in these under-researched fields.