
Abstract
Omega-3 fatty acids (FAs) could play an important role in maintaining cognitive function in aging individuals. The omega-3 FA docosahexaenoic acid is a major constituent of neuronal membranes and, along with the other long-chain omega-3 FAs from fish such as eicosapentaentoic acid, has been shown to have a wide variety of beneficial effects on neuronal functioning, inflammation, oxidation and cell death, as well as on the development of the characteristic pathology of Alzheimer's disease. Omega-3 FAs may prevent vascular dementia via salutary effects on lipids, inflammation, thrombosis and vascular function. Epidemiologic studies have generally supported a protective association between fish and omega-3 FA levels and cognitive decline. Some of the small, short-term, randomized trials of docosahexaenoic acid and/or eicosapentaentoic acid supplementation have found positive effects on some aspects of cognition in older adults who were cognitively intact or had mild cognitive impairment, although little effect was found in participants with Alzheimer's disease. Large, long-term trials in this area are needed.
The greater life expectancy for women translates into a high lifetime risk of cognitive decline and dementia. Based on 30-year follow-up data from Framingham, MA, USA, a middle-aged woman in North America of European descent has a one in five risk of developing dementia of any type [1]. The prevalence of dementia has been estimated to be approximately 6–10% of individuals aged 65 years and older, rising from 1–2% of those aged 65–74 years, to 30% or more of those aged 85 years and older [2]. The economic impact of dementia is enormous – an estimated US$ 100 billion was spent in the USA on providing care for 3 million patients with dementia in 2005, and by 2011, costs are expected to have doubled [201].
Current therapies for treating dementia modify symptoms but not the course of the disease [3]. Preventive strategies to reduce the burden of dementia are clearly needed. It has been estimated that delaying the age of onset of dementia by just 5 years would reduce the lifetime risk of dementia by approximately half [1]. Nutritional interventions could play an important role in slowing the neurodegenration processes, which appears to take decades [4]. Omega-3 fatty acids (FAs) have been shown to play a critical role in early brain develop-ment [202]. Recent studies have found that omega-3 FAs may further protect against cognitive decline with advancing age. Increasing omega-3 FA intake is both safe and achievable. Given a sufficient body of evidence, recommendations to increase omega-3 FA intake could ultimately reduce the burden of age-related cognitive decline in the USA. This article will review the evidence for an effect of omega-3 FAs on cognition.
Aging & cognitive decline
Mild cognitive impairment (MCI) describes the intermediate phase in the continuum of cognitive decline between normal aging-related changes in cognitive function and dementia. Not all individuals with MCI progress to having dementia [5]. Dementia represents a diverse category of syndromes characterized by deficits in memory, cognitive function and behavior severe enough to affect social or occupational functioning [6]. Within the broad category of dementia, approximately 75% is of the Alzheimer's disease (AD) type, with or without vascular dementia, and 15% is vascular in origin [1,7].
Alzheimer's disease pathophysiology
In AD, subtle changes in hippocampal synaptic function occur prior to neuronal apoptosis, before progressing to the characteristic presence of neurofibrillary tangles, senile plaques and changes in neurotransmitter levels [8]. There is increased production of amyloid-β peptide, which results in numerous adverse consequences, including the overproduction of reactive oxygen species and proinflammatory cytokines, demyelination, activation of the apoptotic cell death cascade, neuotransmitter defects and neuronal death [9]. Oxidative stress appears to play a key role in the initiation and progression of AD [10,11]. A variety of mechanisms contribute to oxidative stress from the earliest of phases of AD, including lipid, protein, sugar and DNA and RNA peroxidation. Amyloid-β has been shown to induce oxidative stress-dependent disruption of the microtubule network that leads to neuronal apoptosis. The apolipoprotein ε4 (ApoE4) carrier status is a risk factor for AD [12].
Vascular dementia pathophysiology
Vascular dementia has its etiology in cerebrovascular disease, and its risk factors include hypertension, hyperlipidemia, cigarette smoking, diabetes mellitus and male sex, as well as genetics and loss of cerebral tissue [13]. The lifetime risk of stroke is one in five for women, which is similar to the lifetime risk of AD [1]. Over 60% of stroke survivors will have some degree of cognitive impairment, and a third will develop overt dementia [14]. Conversely, on autopsy, over a third of dementia cases have significant vascular pathology. Vascular cognitive impairment can occur alone or in association with AD, with the greatest cognitive impairment in those with both pathologies. Anatomic brain changes in vascular cognitive impairment include parenchymal lesions (infarcts and hemorrhages in particular) and the vascular lesions from which they arose [14].
Obesity, metabolic syndrome, insulin resistance and diabetes mellitus have been associated with an increased risk of cognitive dysfunction and neurodegeneration [15,16]. The mechanism for the acceleration in brain aging in diabetes mellitus remains unclear. Several hypotheses have been proposed for the process of cognitive dysfunction in diabetes mellitus, which include interference in the cerebral metabolism of tau and amyloid, and increased formation of reactive oxygen species via formation of advanced glycation end product (AGE). Inflammation, critical for the development of atherosclerosis [17], has also been implicated in cognitive decline.
Omega-3 fatty acids
Omega-3 FAs are a family of 18–22 carbon poly-unsaturated FAs (PUFAs) with the last double bond in the n-3 position. Mammalian cells cannot synthesize omega-3 FAs. The parent omega-3 FA is α-linolenic acid (ALA), from which all omega-3 FAs are ultimately derived. The two most important and biologically active omega-3 FAs are eicosapentaenoic acid (EPA; C20:5n-3) and docosahexaenoic acid (DHA; C22:6n-3). ALA is the major omega-3 FA in the diet, with daily mean intakes of 1.6 g for men and 1.1 g for women [18]. The major dietary sources of ALA are vegetable oils such as soybean and canola oils, with flaxseed oil being one of the richest known sources. ALA is also found in a variety of plant products, including nuts (especially English walnuts), seed, vegetables, legume, grains and fruits. Conversion of dietary ALA to DHA and EPA is minimal, so the most efficient way to enrich tissue phospholipids with DHA or EPA is to consume them [19]. Fish and fish oils are the richest sources of DHA and EPA, although terrestrial meats such as beef, pork and chicken may contribute small amounts of omega-3 FAs to the diet.
Omega-3 fatty acids & Alzheimer's disease pathophysiology
Omega-3 FAs are a major constituents of neuronal membranes, with the long-chain DHA comprising 60% of the PUFAs in neuronal cell membranes, with much smaller amounts of EPA present. In the gray matter of the cortex, 30–40% of the FAs are DHA [20], with high concentrations found in synaptic membranes [21] and myelin sheaths [22]. DHA is incorporated more rapidly into gray than white matter, and has a half-life of approximately 2.5 years in the human brain [23]. DHA is essential for brain development as well as normal brain functioning. Adequate DHA levels throughout life are required for normal brain functions such as neurotransmission, synaptic plasticity and vision [24,25].
Neuronal DHA deficiency may contribute to derangements in neuronal metabolism, having specific effects on synaptic vesicle density and neurotransmitter release [26,27]. Membrane fluidity is crucial for normal neuronal functioning for transmitting axonal information, regulating membrane-bound enzymes and controlling ion channels and various receptors. Membranes become more rigid as neurons age owing to decreased levels of total PUFAs, higher levels of cholesterol, a slower rate of cholesterol turnover and greater oxidative stress in the membrane [24]. DHA infusion has been shown to increase synaptosomal membrane fluidity, and was associated with prevention of cognitive decline in amyloid-β-infused rats [28].
Alzheimer's disease patients have decreased serum, brain and neuronal DHA levels compared with age-matched controls [29–32], suggesting that a DHA deficiency could contribute to cognitive impairment. Oxidative stress appears to play a key role in the development of AD, and excessive oxidation of DHA to F4-neuroprostanes has been reported in AD patients and so could explain the reduced DHA levels in these patients [33]. PUFA deficiency in the aging mouse brain leads to progressive memory loss, cognitive alterations and learning disabilities, changes that are reversed by supplementation with omega-3 PUFAs in the form of fish oil or DHA [24,34,35]. Of particular importance for the development of AD, DHA supplementation has been shown to protect against amyloid-β production, accumulation and toxicity, as well as accumulation of hyper-phosphorylated tau as neurofibrillary tangles and cognitive impairment in mouse and rat models of AD [36–38]. Improvements in memory, learning and a reduction in spatial cognitive deficits have also been shown [37,39]. DHA pretreatment prevents membrane disorganization from amyloid-β by increasing neuronal survival following amy-loid-β infusion through preventing cytoskeletal perturbations, capsase activation and apoptosis along with promoting extracellular signal-related kinase-related survival pathways [11,40].
Early stages of AD are characterized by inflammation, upregulation of cell cycle proteins, adhesion and regeneration-related factors [41]. Activation of microglia and recruitment of astrocytes, both of which are important brain immunomodulators, are found in areas of amyloid-β deposition [42]. DHA derivatives with conjugated triene structures have immuno-regulatory and neuroprotective properties. Human cell culture studies have shown that 10–17 S-docosatrienes, or neuroprotectin D1, decreased amyloid-β production by 25% and decreased amyloid-β-associated neuroapoptosis by 50% [43]. Injection of soluble amyloid-β oligomers causes a rapid and transient accumulation/activation of the cytosolic phospholipase A2 that leads to the release of arachidonic acid (AA), an omega-6 FA that appears to be a mediator of neuronal apoptosis [44]. AA metabolism produces both pro- and anti-inflammatory prostaglandins, leukotrienes, lipoxins and thromboxanes (see Figure 1 and further discussion in the following sections of this article) and may activate apoptotic pathways, such as the sphingomyelinase pathways, for the production of ceramides [45].
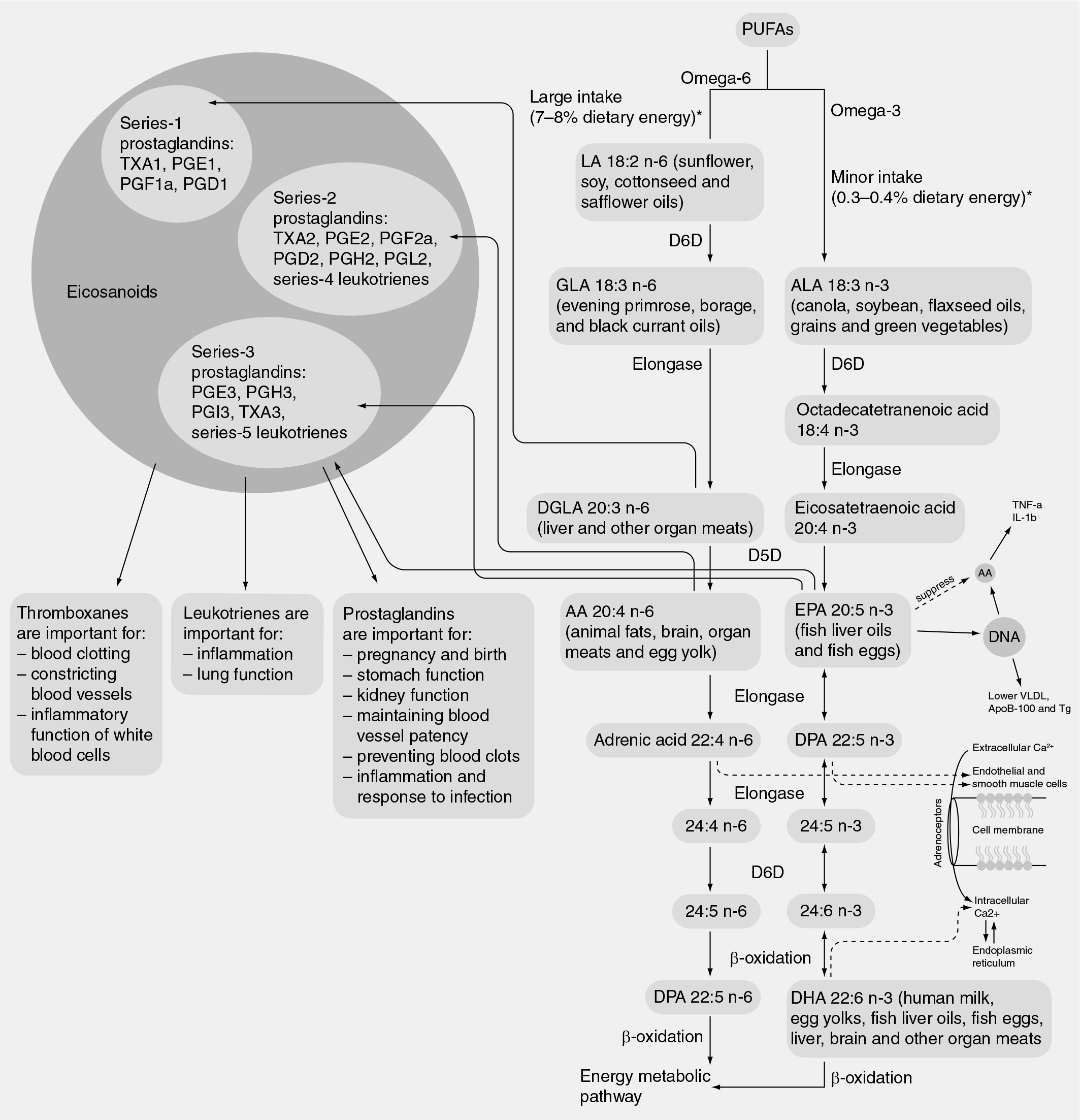
Overview of essential fatty acid metabolism from the Agency for Healthcare Research and Quality omega-3 fatty acid review.
Proinflammatory eicosanoid production from AA is partially balanced by anti-inflammatory n-3 metabolites. DHA has been shown to maintain the balance between n-3 and n-6 PUFAs in cultured astrocytes [46]. With aging, the synthesis of EPA and DHA falls while AA undergoes increased peroxidation to produce increased levels of inflammatory metabolites [47]. Higher levels of omega-6 FA–including AA– consumption have been associated with an increased risk of dementia [48]. DHA undergoes enzymatic oxidation through a 15-lipoxygenase-like enzyme, producing secondary messengers, docosanoids (17 S-resolvins, 10–17 S-docosatrienes and pro-tectins), which act as anti-inflammatory chemical lipid mediators through neuroprotectin D1 and resolving D and E receptors [49]. DHA also undergoes nonenzymatic oxidation with the production of neuroprostanes, which have anti-inflammatory effects [50]. Dietary DHA is the precursor of the 10–17 S-docosatriene neuroprotectin D1, a neuroprotective compound that decreases amyloid-β apoptosis and oxidative stress, both of which are decreased in the hippocampi of AD patients [51].
Depending on the level and duration of supplementation, omega-3 FAs can modulate gene expression in a number of tissues. Fish oil consumption alters the transcription of various genes in hippocampal cells of aged rats, including the gene encoding the amyloid-β-scavenger transthyretin [52–54]. Other PUFAs, including omega-6 FAs, have also been shown to control gene expression in the brain and other tissues [55–57], and have been shown to be ligands of brain-intranuclear peroxisome proliferator-acti-vated and retinoid X receptors [58,59]. Increased levels of saturated FAs have been shown to induce amyloidogenesis and tau hyperphosphorylation along with abnormal glucose metabolism [60].
Omega-3 fatty acids & vascular disease
Omega-3 FAs may influence the development of vascular dementia through beneficial effects on a number of cardiovascular risk factors [27,61]. The primary lipid effect of EPA and DHA is a dose-dependent reduction in plasma triglyceride levels. Omega-3 FAs lower triglycerides though a variety of mechanisms, including reduced hepatic lipogenesis, increased hepatic β-oxidation and increased intrahepatic degradation of apolipo-protein B-100, which together markedly reduce the secretion of very-low-density lipoproteins (VLDLs) [62]. They also accelerate VLDL and chylomicron triglyceride clearance by increasing lipoprotein lipase activity [63], increasing the conversion of VLDL to low-density lipoproteins (LDL) [64] and reducing postprandial lipemia [65]. Omega-3 FAs can also modestly lower blood pressure in hypertensive and older patients, as well as in diabetic patients [66]. No consistent effects on glucose metabolism have been found [61].
Inflammation, thrombosis and endothelial function are intrinsically linked processes in the development of atherosclerosis, and share several key molecular mechanisms. The anti-inflammatory effects of omega-3 FAs are thought to be mediated, at least in part, by reduced synthesis of inflammatory molecules from AA, as discussed previously. Products of AA metabolism are important regulators of cellular functions and many (but not all) of these products have atherogenic and prothrombotic effects [47]. Omega-3 FAs may also exert anti-inflammatory effects via the production of resolvin E1, an oxidized derivative of EPA produced by aspirin-blocked cyclo-oxygenase-2, which reduces inflammation by suppressing the activation of NF-κβ and consequently the synthesis of inflammatory cytokines and chemokines [67].
Increased cardiovascular risk has been asso-ciated with TNF-α, serum amyloid A and IL-1, −6 and −10, which are proinflammatory cytokines produced by monocytes and macrophages. Both EPA and DHA inhibit production of IL-6, the only cytokine that stimulates the synthesis of all the acute phase reactants involved in inflammation, as well as other inflammatory cytokines [27,64,68–76]. C-reactive protein (CRP), a hepatic acute phase reactant, is considered a summary index of inflammation, and has consistently been shown to be an independent risk factor for cardiovascular disease [77,78]. However, most studies have not shown a reduction in CRP with omega-3 FA supplementation [27,64,68–76]. Soluble adhesion factors (e.g., soluble intercellular adhesion molecule-1, soluble vascular adhesion molecule-1 and sP-selectin) are involved in platelet reactivity, endothelial activation and plaque development, and may also provide a measure of the inflammatory processes underlying atherosclerosis [70,75,79–81]. Most studies of EPA and DHA have demonstrated a decrease in soluble vascular adhesion molecules and decreased monocyte adhesion. EPA and DHA are incorporated into leukocyte and monocyte membranes. Fish oil supplementation can suppress monocyte synthesis of the prothrombotic thromboplastin and thromboxane A2 [81,82] and can decrease T-lymphocyte activation [81]. Fish oil supplementation has also been shown to suppress cyclo-oxygenase-2 expression in blood monocytes via inhibition of Toll-like receptor-mediated signaling pathways [83].
Omega-3 FAs have beneficial effects on endo-thelial function, which may also have implications in vascular dementia. Rats fed a fish oil-enriched diet were found to have increased endothelium-dependent vasodilatation in response to acetyl-choline infusion, which was associated with increased activity of endothelial nitric oxide synthase [84]. These observations have been con-firmed in human trials in which omega-3 FA supplementation improved endothelial function in patients with congestive heart failure, coronary heart disease (CHD) and dyslipidemia [85–87], and fish oil supplementation has been found to reduce systemic nitirc oxide production [88]. Epidemiological investigations in Japan in the late 1980s revealed that pulse wave velocity was significantly slower in residents of a fishing village compared with age- and sex-matched subjects from a farming village in the mountains [89]. Death rates from ischemic heart disease correlated with these differences, and were found to be 86% lower in the coastal region. Similar findings were reported in Australian fish-eaters versus non-fish eaters where arterial compliance (derived from pulse wave velocity analyses) was 25% greater in the former compared with the latter group [90]. In the Cardiovascular Health Study, an inverse relationship between the intake of oil-rich fish and systemic vascular resistance was observed [91], and a supplementation study confirmed that higher EPA and DHA intakes improved systemic arterial compliance [92].
In addition, omega-3 FAs may have antithrom-botic effects. Procoagulant factors such as fibrino-gen factors VII and VIII, von Willebrand factor and plasminogen activator inhibitor-1 have been associated with increased cardiovascular risk in several studies [93–95]. Omega-3 FAs have had variable effects compared with these factors [61]. Large doses of marine omega-3 FAs can increase bleeding times; however, no increase in clinically significant bleeding episodes has been reported in clinical trials to date [96]. Omega-3 FAs have been shown to influence gene regulation by reducing the expression of platelet-derived growth factors [97]. Omega-3 FAs suppress platelet activating factor, a potent platelet aggregator, and leukocyte activator. Nonetheless, most studies of platelet aggregation found no effect from omega-3 FAs using a variety of methodologies. Reports on the influence of omega-3 FA supplementation on homocysteine levels have been variable [98,99].
Stroke is a major contributor to vascular cognitive dysfunction. Fish intake and stroke incidence have been evaluated in several epidemiologic stud-ies. In the Zutphen study, the risk of stroke in men who consumed an average of 20 g of fish per day was 50% less than in those who consumed smaller amounts of fish [100]. In the NHANES Epidemiologic Follow-Up Study, white females who consumed fish more than once per week had an age-adjusted stroke incidence that was half that of women who did not consume fish [101]. A similar protective effect was observed in both Black women and men but not in White men. A trend (p = 0.06) towards reduced stroke risk with increasing reported fish consumption was also reported in the Nurses' Health Study [102]. The Cardiovascular Health Study reported a protective association between modest fish intake and ischemic stroke, as well as subclinical infarcts on MRI [103,104]. By contrast, both the Chicago Western Electric Study [105] and the Physicians' Health Study [106] did not find a relationship between reported fish intake and stroke risk. Serum fatty acid levels of ALA were negatively associated with stroke incidence in the Multiple Risk Factor Intervention Trial [107].
Clinical trials of omega-3 FA supplementation have inconsistently found evidence of a protective effect of omega-3 FAs on stroke. The Lyon Diet Heart Study (testing a Mediterranean-style diet enriched with ALA) [108] and the GISSI- Prevention study (testing the effects of 850 mg of supplemental omega-3 FAs), found no significant effect of omega-3 FAs on the incidence of stroke, although the risk of CHD was reduced [109]. The Japan EPA Lipid Intervention Study reported a 20% reduction in nonfatal CHD and recurrent stroke, but not in the primary prevention of stroke from 1800 mg of EPA daily [110]. Some evidence suggests the associations could differ if the data were analyzed for type-specific stroke incidence. Fish intake has the most consistent inverse relationship to ischemic stroke [111].
A reduced risk of ischemic stroke from increased omega-3 FA intake could be mediated by changes in plaque morphology that reduces the risk of acute plaque rupture. In a study of 188 patients awaiting carotid endarterectomy, those who received fish oil had higher levels of omega-3 FAs incorporated into their carotid plaques and an increase in plaque-stabilizing characteristics, such as thicker fibrous caps, absence of inflammation and fewer macro-phages [112]. EPA has been shown to reduce the progression of carotid intima-media thickness in diabetic subjects [113], and fish intake and higher levels of plasma DHA are associated with slower progression of coronary atherosclerosis in women with CHD [114,115].
Menopause & hormone therapy
Estrogen receptors are present in the hippocampus, which plays a role in memory and is especially affected by aging and AD [116,117]. Estrogens have a number of neuroprotective effects including decreased accumulation of amyloid-β peptide, antioxidant activity, inhibition of neuronal apoptosis, modulation of neurotransmitter systems important in memory formation, modulation of ApoE gene expression, increased blood flow and reduced ischemia [118–120]. As women age and enter menopause, there is a reduction in blood estrogen levels owing to a loss of ovarian follicles.
Although observational data have suggested that early initiation of postmenopausal oral estradiol therapy had a protective effect on the development of AD [121], the Women's Health Initiative – Mental Status studies found a negative impact of conjugated equine estrogen (CEE) alone and with medroxyprogesterone acetate (MPA) on domains of cognitive function and the development of cognitive impairment in women aged 65 years and older [122–126]. The mechanisms whereby postmenopausal hormone therapy exerts harmful effects on cognition have yet to be elucidated. In the WHI hormone trials as a whole, both the CEE and CEE plus MPA groups had a higher risk of stroke compared with placebo, although only CEE plus MPA conferred an excess risk of CHD [127,128]. In a subgroup of women who underwent MRI, mean ischemic lesion volumes were similar in the hormone therapy and placebo groups [129]. However, those receiving hormone therapy had greater brain atrophy, especially if cognitive deficits were present [130].
Several studies suggest the possibility that hormonal status is one of the factors influencing omega-3 FA status. Young women convert the plant omega-3 FA, ALA, to EPA and DHA with a greater efficiency than do young men [131–133]. Endogenous estrogen seems to stimulate the bio-synthesis of long-chain omega-3 FAs from common dietary precursors, presumably via stimulation of the δ-5 and δ-6 desaturases, the rate limiting enzymes in PUFA biosynthesis (Figure 1). Postmenopausal women who take exogenous estrogen may not enjoy the same benefit. A study comparing red blood cell EPA plus DHA levels in healthy women over the age of 50 years, who were or were not on postmenopausal hormone therapy, found no evidence for an effect on membrane omega-3 FA levels owing to exogenous hormone status. Some studies have demonstrated that hor-mone therapy attenuates the conversion of ALA to DHA and DHA to EPA and the accumulation of EPA in serum phospholipids [134].
No data are available regarding the interaction between omega-3 FAs and postmenopausal hormone therapy on the pathophysiology of AD and vascular dementia. Several adverse effects of estrogen and/or progestin administration have been associated with increased cardiovascular risk, and some of these may be ameliorated by omega-3 FAs. For example, the addition of MPA to CEE has been shown to ameliorate the beneficial changes of CEE on LDL-cholesterol, high-density lipoprotein cholesterol and endothelial function [135,136]. Both CEE and CEE plus MPA have been shown to markedly increase inflammatory markers, including CRP, and to increase triglyceride levels [135,137].
Epidemiological studies
Table 1 summarizes epidemiologic studies of the association between omega-3 FAs and cognitive decline and/or dementia. Cross-sectional studies have found variable relationships between omega-3 FA levels and intake [29,138–144]. The largest was from Norway and included 2031 subjects, approximately half of whom were women [141]. This study used a food frequency questionnaire to estimate exposure to omega-3 FAs, and found a significant inverse relationship between the reported intake of fish (only 2% consumed no fish or fish products) and cognitive status.
Epidemiologic studies of omega-3 fatty acids and cognition.
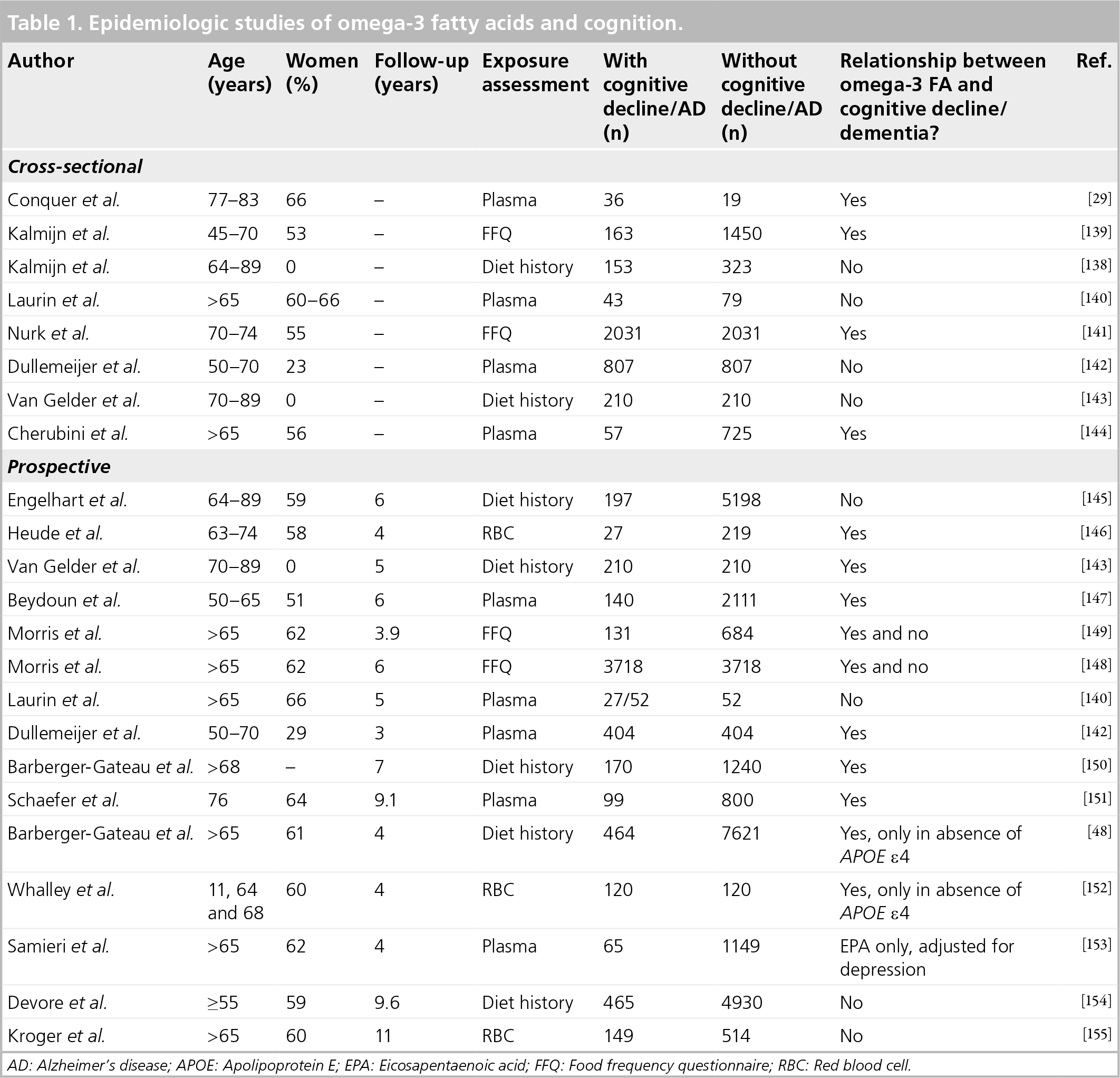
AD: Alzheimer's disease; APOE: Apolipoprotein E; EPA: Eicosapentaenoic acid; FFQ: Food frequency questionnaire; RBC: Red blood cell.
Prospective cohort studies have generally been more supportive of the protective relationship between omega-3 FAs and cognitive status [48,140,142,143,145–155]. Follow-up periods ranged from 3–11 years. Approximately 60% of par-ticipants in the prospective studies were women.
Subgroup analyses by gender were not performed. The largest, the Three City Cohort study, assessed the development of AD or cognitive impairment over 4 years as a function of reported fish intake in approximately 8085 men and women over the age of 65 years [48]. A significant protective effect of fish intake was observed, but only in those subjects who were noncarriers of APOE ε4. Benefit in the absence of the APOE ε4 allele has been replicated in some but not all studies [156]. The Framingham Heart Study (FHS) had a follow-up at 9 years, with a mean age of 76 years at baseline [151]. After adjusting for age, education, APOE ε4 status and plasma homocysteine, those subjects in the highest quartile of plasma DHA were 47% less likely to develop dementia (p = 0.04) and 59% less likely to develop AD (p = 0.14). These subjects had an average fish intake of at least three servings per week.
Clinical trials
Seven randomized, controlled trials evaluating the effects of omega-3 FAs on cognition have been published; two included cognitively intact individ-uals [157,158], two included individuals with MCI [159,160], two included individuals with AD [160,161] and two enrolled only patients with Huntington's disease, a hereditary, premature-onset dementia (
Randomized, controlled trials of omega-3 fatty acids on cognition and brain morphology.

AA: Arachadonic acid; ADAS-Cog: Alzheimer's Disease Assessment Scale - Cognitive; CDRS: Clinical Dementia Rating Scale; CIBIC+: Clinician's Interview-Based Impression of Change Scale; DHA: Docosahexaenoic acid; EPA: Eicosapentaenoic acid; FA: Fatty acid; MCI: Mild cognitive impairment; MMSE: Mini Mental State Examination
In the largest trial in cognitively intact subjects, van de Rest et al. randomized 302 participants to higher dose (1.8 g) EPA plus DHA, lower dose (0.4 g) EPA plus DHA, or placebo for 26 weeks [157]. The mean age was 70 years and 45% were female. Omega-3 FA supplementation had no effect on the cognitive domains at either 13 or 26 weeks, as measured by an extensive neuropsychological test battery that included the cognitive domains of attention, sensorimotor speed, memory and executive function. Very few subjects were lost to follow-up (1.8 g omega-3 FA group: 1/96; 0.4 g omega-3 FA group: 0/100; placebo: 4/106 at 26 weeks). Nor were any effects found on wellbeing or depression, as measured by a number of tests [164]. By contrast, a smaller trial that randomized 49 women to one of four groups found that 0.8 g DHA with or without 12 mg lutein improved verbal fluency after 4 months compared with placebo [158]. The combination of DHA and lutein improved memory scores, with a trend toward more efficient learning. No effects on mental processing speed, accuracy or mood were found.
Kotani et al. studied 21 subjects (9 female) with MCI, who were randomized to either 240 mg/day of the omega-6 FA, AA and DHA supplementation or placebo for 90 days [159].
At the end of the study, an improvement was observed in immediate memory and attention in the AA plus DHA group (p < 0.01). Visuospatial/construction skills and language showed no improvement in either group after supplementation. The authors postulated that DHA might not improve cognitive function directly, but via improving membrane function and regional cerebral blood flow.
Chiu et al. performed a pilot study of 46 subjects evenly divided between MCI or AD, who were randomized to either 1.8 g omega-3 FAs (EPA + DHA) or olive oil placebo, and were followed for 24 weeks [160]. The mean age was 75 years and 46% were women. In the MCI group, omega-3 FAs improved cognition (Alzheimer Disease Assessment Scale – Cognitive: p < 0.03; Clinician's Interview-Based Impression of Change Scale: p = 0.008). No cognitive improvement was observed in AD subjects receiving omega-3 FAs. Interestingly, a greater proportion of EPA incorporated into red blood cell membranes was associated with better cognitive outcome (p = 0.003). Omega-3 PUFAs were well tolerated. However, 17% of the omega-3 FA group and 32% of the placebo group were not included in the final analysis, limiting conclusions from this trial.
In a trial in AD patients, Freund-Levi et al. enrolled 204 subjects with mild-to-moderate AD and a Mini Mental State Examination (MMSE) of greater than or equal to 15 whose condition was stable on acetylcholine esterase inhibitors treatment [161]. The mean age was 73 years and 51% were women. Subjects were randomized to double-blind DHA 1.7 g, EPA 0.6 g or placebo for 6 months, followed by greater than or equal to 6 months of open-label omega-3 FAs. No difference in cognitive decline was observed over the first 6 months of treatment using the MMSE, the Alzheimer's Disease Assessment Scale – Cognitive and the Clinical Dementia Rating Scale. However, in a subgroup of subjects (n = 23) with very mild cognitive decline (MMSE >27), there was a significant reduction in MMSE decline rate in the omega-3 FA group compared with placebo (p < 0.05) with improve-ment over time in delayed word-recall and attention in this subgroup of participants on omega-3 FAs compared with placebo. The placebo group with very mild cognitive decline experienced a similar arrest in MMSE decline after subsequent treatment with omega-3 FAs. No significant treatment effects on activities of daily living or on caregivers' burden were observed, but in APOEε4 carriers, there was a beneficial effect on agitation and depression. Omega-3 FA treatment was safe and well tolerated, with a positive effect on weight and appetite, especially in the absence of the APOE ε4 allele [165]. The drop-out rate during the double-blind treatment period was 13% (12 in the omega-3 FAs and 14 in the placebo group) and 2% during the open-label treatment period (2 in each group).
Puri et al. found that 1 year of ethyl-EPA treatment in 30 subjects with Huntington's disease reduced global brain atrophy (as seen by MRI), with the greatest benefits observed in the caudate nucleus and posterior thalamus, compared with placebo [163]. Women accounted for 50% of the subjects. By contrast, the TREND-HD study did not show any cognitive benefit in ethyl-EPA diet compared with placebo in subjects with Huntington's disease followed-up over 6 months [162]. Cognition was measured using the MMSE and 50% of participants were women. A total of six of 158 subjects in the EPA group and two of 158 subjects in the placebo group were excluded from the analysis.
It should be noted that depression, which may influence cognitive function, has not been shown to consistently respond to omega-3 FA supplementation [164,166,167,203].
Summary
The greater life expectancy for women translates into a high lifetime risk of cognitive decline and dementia, with a middle-aged woman in North America of European descent having a one in five risk of developing dementia of any type. Omega-3 FAs could play an important role in maintaining cognitive function in aging individuals. The omega-3 FA DHA is a major constituent of neuronal membranes and is essential for normal brain development. Blood levels of DHA (and, in some studies, EPA) are decreased in individuals with AD, suggesting that deficiency could contribute to cognitive impairment. In animal and in vitro studies, omega-3 FAs have been shown to have a wide variety of beneficial effects on neuronal functioning, inflammation, oxidation and cell death. They have also been shown to protect against the development of cognitive impairment in AD animal models and to decrease the characteristic pathology of AD, including a reduction in amyloid-β production, accumulation and toxicity, as well as slowing neurofibrillary tangle formation.
Vascular dementia constitutes approximately 15% of dementia and also contributes to further cognitive decline in the 75% of patients with AD. Omega-3 FAs may inhibit vascular dementia via salutary effects on lipids, inflammation, thrombosis and vascular function. Fish intake has generally been associated with a lower risk of stroke, which is a major contributor to cognitive decline. Although most clinical trials of omega-3 FA supplementation have been too underpowered to detect an effect on stroke, the largest study to date (a subanalysis of the Japan EPA Lipid Intervention Study) found a 20% reduction in the risk for recurrent stroke in patients who were treated with EPA. Estrogen can increase the conversion of dietary ALA to EPA, and women have been reported to have higher levels of EPA than men prior to menopause. Following meno-pause, women may be at increased risk of cognitive impairment owing to the loss of estrogen's effects on the hippocampus (which plays a role in memory), as well as to the loss of estrogen's ben-eficial effects on neuronal function, blood flow and inhibition of amyloid-β formation. Whether the loss of estrogen also adversely affects omega-3 FA tissue levels is not known.
Epidemiologic studies have generally supported a protective association between fish and omega-3 FA levels and cognitive decline. Some studies found a beneficial association from omega-3 FAs only in those subjects who were not carriers of APOE ε4. Randomized trials of omega-3 FA supplementation on AD/dementia end points have been small (≤300 participants) and short (≤12 months). Omega-3 FA supplementation had some positive effects on certain aspects of cognition, such as memory and attention, in older adults who were cognitively intact or had MCI. Little effect was found on cognition in patients with AD or Huntington's disease in studies using a variety of assessment tools.
Future perspective
Further elucidation of the role of long-term levels of DHA and EPA from epidemiologic studies may help to clarify the role of omega-3 FAs on the development of early cognitive dysfunction. Brain imaging and functional studies may also be helpful. Subgroup analyses of existing studies and large long-term studies of women may also help to determine changes in DHA and EPA levels during the menopause, as well as longer-term associations with gradual cognitive changes leading to later cognitive impairment. Recommendations for consistent assessment of cognitive function are needed to permit comparison and pooling of study populations.
Executive summary
Cognitive impartment and dementia increase with advancing age.
A total of 30% of those aged 85 years and older have dementia.
Owing to aging demographics, expenditures associated with dementia are expected to increase to more than US$ 1 trillion by 2050.
Therapies to prevent or slow cognitive decline are needed.
Omega-3 fatty acids (FAs) are major constituents of neuronal membranes.
Omega-3 FAs have a wide variety of potential beneficial effects on neuronal functioning, inflammation, oxidation and cell death.
Omega-3 FAs prevent development of the characteristic pathology of Alzheimer's disease in animal models.
Vascular dementia constitutes approximately 15% of dementia and contributes to further cognitive decline in the 75% of patients with Alzheimer's disease.
Omega-3 FAs may prevent vascular dementia via salutary effects on lipids, inflammation, thrombosis and vascular function.
Omega-3 FAs and fish intake have protective associations for stroke.
Some evidence for stroke prevention has emerged from clinical trials of omega-3 FA supplementation.
Endogenous estrogen increases omega-3 FA levels.
Postmenopausal estrogen therapy may negatively influence omega-3 FA levels.
Omega-3 FA levels and fish intake are consistently associated with reduced risk of cognitive decline in epidemiologic studies in women and men.
APOE alleles influenced the association between omega-3 FAs and dementia in some but not all studies.
Three of the four small, short-term trials of omega-3 FAs have generally shown positive effects on some aspects of cognition in older adults who were cognitively intact or had mild cognitive impairment.
Two small, short-term trials of omega-3 FAs found little effect on cognitive decline in patients with Alzheimer's disease in small trials.
Large, long-term trials are needed to evaluate the effect of omega-3 FA supplementation on preventing or slowing cognitive decline with advancing age.
Several ongoing trials with greater than or equal to 10,000 participants are evaluating the long-term (≥5 years) effect of omega-3 FA supplementation on cardiovascular disease, although none have cognitive changes as a major end point [204–206]. Since the potential public health impact of preventing progression to dementia is great, it is hoped that cognitive function studies ancillary to these trials are planned. Such studies should utilize tests that are sufficiently sensitive to detect more gradual decline in cognitive function in cognitively intact older adults as well as in those with MCI. In addition, studies that are adequately sized to assess a treatment interaction with APOEε4 would be desirable, since epidemiologic studies suggest that the absence of the APOEε4 allele may influence whether omega-3 FAs have beneficial effects.
Footnotes
This project was supported (or supported in part) by an Agency for Healthcare Research and Quality (AHRQ) Centers for Education and Research on Therapeutics cooperative agreement #5 U18 HSO16094. Jennifer G Robinson has received in the past year: grants from Abbott, Aegerion, Astra-Zeneca, Bristol-Myers Squibb, Daiichi-Sankyo, Glaxo Smith Kline, Hoffman La Roche, Merck and Merck Schering-Plough. William Harris is a consultant to Monsanto Co. and GlaxoSmithKline; both companies have interests in omega-3 fatty acids. He is also the founder and chief scientific advisor to Omega Quant Analytics, a company that offers blood omega-3 testing. The authors have no other relevant affiliations or financial involvement with any organization or entity with a financial interest in or financial conflict with the subject matter or materials discussed in the manuscript apart from those disclosed.
No writing assistance was utilized in the production of this manuscript.