
Abstract
Chlorophyll and carotenoid content (ChCar), lipid peroxidation (LP) and growth parameters (GP) in plants are often used for environmental pollution estimation. However, the nonmonotonic dose-response dependences (hormesis and paradoxical effects) of these indices are insufficiently explored following exposure to different pollutants. In this experiment, we studied nonmonotonic changes in ChCar, LP, GP in wheat seedlings (Triticum aestivum L.) upon exposure to lead, cadmium, copper, manganese, formaldehyde, the herbicide glyphosate, and sodium chloride in a wide range from sublethal concentration to 102−105 times lower concentrations. 85.7% of dose-response dependences were nonmonotonic (of these, 5.5% were hormesis and paradoxical effects comprised 94.5%). Multiphasic dependences were the most widespread type of paradoxical effect. Hormesis was a part of some multiphasic responses (i.e. paradoxical effects), which indicates a relationship between these phenomena. Sublethal pollutant concentrations significantly increased LP (to 2.0–2.4 times, except for manganese and glyphosate) and decreased GP (to 2.1–36.6 times, except for glyphosate), while ChCar was reduced insignificantly, normalized or even increased. Lower pollutant concentrations caused a moderate deviation in all parameters from the control (not more than 62%) for hormesis and paradoxical effects. The seedling parameters could have different types of nonmonotonic responses upon exposure to the same pollutant.
Keywords
INTRODUCTION
At the present time, evidence has accumulated in toxicology that, apart from classical monotone dose-response dependences (S-shaped, exponential), nonmonotonic responses, which include hormesis (Cedergreen et al. 2007; Calabrese and Blain 2009) and paradoxical effects (Schatz 1999; Batyan et al. 2009; Smith et al. 2012), are also found rather often.
Hormesis is a biphasic dose-response phenomenon characterized by low-dose stimulation and high-dose inhibition (Calabrese 2008). It is known that the manifestation of paradoxical effects consists of the following: as the dose or concentration of the toxic agent is reduced, its toxicity increases, and vice versa, such that with an increase in the dose, its effect is reduced (Schatz et al. 1964; Batyan et al. 2009).
Chlorophyll and carotenoid content, lipid peroxidation in the plant, and growth parameters of various plant species are widely used to estimate the level of environmental pollution (Carreras et al. 1996; Pignata et al. 1999; Madkour and Laurence 2002; Wahsha et al. 2012). However, the nonmonotonic dose-response dependences of these indices have been insufficiently explored under the influence of different pollutants. In the conditions of ecosystems of contaminated territories, a whole range of pollutants exert an impact on a plant, which hampers analysis of the nature of influence of separate substances. Experimental models are more adequate when studying this issue compared with the conditions of contaminated territories.
Therefore, the aim of this study was to study nonmonotonic changes in wheat seedling (Triticum aestivum L.) parameters such as photosynthetic pigment content, lipid peroxidation, and growth parameters upon exposure to different environmental pollutants (heavy metals: lead, cadmium, copper, manganese, as well as formaldehyde, herbicide glyphosate and sodium chloride) in wide range from sublethal concentrations to 102−105 times lower concentrations.
In this study, we asked how often the pollutants induced nonmonotonic responses in the studied plant parameters, what type of nonmonotonic dose-response dependences occurred most frequently (hormesis or paradoxical effects) for the studied plant parameters, if there was relationship between hormesis and paradoxical effects, whether nonmonotonic dose-response dependences were similar for different plant parameters, and in what dose ranges nonmonotonic responses were seen. In addition, we explored the magnitude of the maximum stimulatory (maximum inhibitory) response and the width of the stimulatory (inhibitory) zone of the studied plant parameters upon hormesis and paradoxical effects.
We use the term hormesis to indicate a biphasic dose-response dependence in which the studied plant parameter undergoes an improvement at low doses and an impairment at high doses under the influence of a pollutant. We consider an antioxidant effect, an increase in the photosynthetic pigment content and increased growth as parameters indicating an improvement in the plant. A prooxidant effect, a decrease in the photosynthetic pigment content and reduced growth are parameters considered to indicate an impairment in the plant.
We considered that a paradoxical effect is a biphasic or multiphasic dose-response dependence characterized by a reduction in a stimulatory or inhibitory pollutant effect to the control level (or a switch from inhibition to stimulation) with an increase in concentration (dose) at one or several sites along the curve. A stimulatory pollutant effect is an improvement in a plant parameter, while an inhibitory effect is an impairment of a plant parameter.
MATERIALS AND METHODS
Pollutant selection
Widespread pollutants with a variety of chemical characteristics were selected for this study. At present, heavy metals and formaldehyde are priority pollutants of the atmospheric air and soil in large cities, caused by the rapid growth in the number of cars (Belkina, 2008) and industrial emissions (Cheng 2003). We selected heavy metals which are known xenobiotics (lead, cadmium) and microelements (copper, manganese) for plants.
We used the herbicide glyphosate (trade name Glifos, produced by Cheminova A/S, Denmark). Glyphosate is also known as Roundup. Glyphosate (N-(phosphonomethyl)glycine) is a broad-spectrum systemic herbicide. It is used to control weeds in a wide variety of agricultural, urban, lawn and garden, aquatic, and forestry situations (Cox 1998). Glyphosate kills plants by interfering with the synthesis of the aromatic amino acids phenylalanine, tyrosine, and tryptophan. It does this by inhibiting the enzyme 5-enolpyruvylshikimate-3-phosphate synthase, which catalyzes the reaction of shikimate-3-phosphate and phosphoenolpyruvate to form 5-enolpyruvyl-shikimate-3-phosphate (Steinrücken and Amrhein 1980). Glyphosphate is usually absorbed through the foliage. Its absorption through the root system is insufficiently explored.
In northern countries, sodium chloride is used as a winter de-icing agent on roads (Belkina, 2008; Cunningham et al. 2008). There is a relationship between sodium chloride accumulation, in leaves and in soil, and injury to roadside plants (Bryson and Barker 2002).
Plant parameter selection
For our study, we selected plant parameters that are widely used to estimate the level of environmental pollution, such as photosynthetic pigment content, lipid peroxidation and growth parameters (Carreras et al. 1996; Pignata et al. 1999; Madkour and Laurence 2002; Wahsha et al. 2012). In addition, we selected parameters with variable significance for the phenotypic adaptation of the plant. It is known that some products of lipid peroxidation, such as MDA, can regulate the function of proteins and gene activity, and always take part in phenotypic adaptation to stress factors in plants (Brand et al. 2004; Weber et al. 2004). Photosynthetic pigments and growth parameters do not have a similar biological activity and depend on the energy status and plastic plant metabolism.
Plant material and growth conditions
The effect of each pollutant was studied in a separate experiment. Seven experiments were carried out. The wheat variety Moscow-39 was used for this study. Plants were grown in Petri dishes on a filter paper substrate moistened with solutions of pollutants. We studied the following concentration ranges of pollutants: 3.13·10−4−1.28 g L−1 lead nitrate, 3.13·10−4−0.08 g L−1 cadmium sulfate, 7.8·0−5−0.32 g L−1 copper sulfate, 1.25·10−3−5.12 g L−1 manganese sulfate, 1.56·10−4−0.08 g L−1 formaldehyde, 2.44·10−6−0.01 g L−1 glyphosate, and 0.25·10−2−5.12 g L−1 sodium chloride.
Neighboring pollutant concentrations differed by two-fold. The range of concentrations was selected based on previous experiments so that the maximum concentration of each toxicant was sublethal, i.e., it had a considerable toxic effect on the growth of the seedling, but did not cause the death of plants. Plants in the control groups were grown with distilled water. Solutions of the pollutants for treatment were also prepared using distilled water. We used distilled water for plant culture to exclude competitive and chemical interactions between pollutants and compounds in a nutrient solution. This is particularly important for the study of low pollutant concentrations. It is known that heavy metals compete with some macro and microelements for absorption by plant roots (Yang et al. 1996; Kabata-Pendias and Pendias, 2000).
Every treatment or control had five Petri dishes with 60 seedlings in each. The plants were grown in a plant culture chamber with a 17 h light period at a light intensity of 40 W·m−2, 22/17°C day/night temperature, and 75±3% relative humidity. Every day, the solutions were replaced with fresh ones. On day 8, the photosynthetic pigment content, lipid peroxidation, and growth parameters of wheat seedlings were evaluated.
Plant growth parameters
Previously, we found that seven-day wheat seedlings have a strong correlation (r=0.80; p<0.05) between shoot (root system) length and the dry biomass of these organs. Therefore, the length of the shoot and the root systems was used as an indicator of growth. These parameters were measured for 30 seedlings in each treatment or control. Six plants were taken from each Petri dish (six seedlings × five Petri dishes for that treatment).
Photosynthetic pigments
The photosynthetic pigment content in the first plant leaf was determined in ten biological replicates (n = 10) for each treatment as well as the control. Leaves from 10 plants in the same group were pooled for one biological replicate.
Fresh leaves from 10 plants (0.2 g) were cut into pieces and homogenized in 80% acetone in the presence of quartz and calcium carbonate. The homogenate was filtered and then diluted to 25 mL with 80% acetone. The absorbance of the mixture was measured at 470, 646, and 663 nm using a UFV-vis spectrophotometer (SF 2000, OKB Spectrum, Russia). The concentrations of chlorophylls and carotenoids were calculated according to the formulae of Lichtenthaler (1987).
Lipid peroxidation
The level of lipid peroxidation was determined in terms of malondialdehyde (MDA) concentration according to the method of Heath and Packer (1968) with the modifications of Kamyshnikov (2002). The MDA content in the first plant leaf was determined in ten biological replicates (n = 10) for each treatment as well as the control. Leaves from 10 plants in the same group were pooled for one biological replicate. Approximately 0.2 g of fresh leaves from 10 plants was homogenized in 2 mL of 3 mM ethylenediaminetetraacetate (EDTA). Then, 2 mL of 20% trichloroacetic acid (TCA) and 2 mL of 0.75% 2-thiobarbituric acid (TBA) were added to 2 mL of the homogenate. The mixture was incubated at 96ºC for 30 min and then transferred into an ice bath to stop the reaction. The tubes were centrifuged at 10 000 g for 15 min and the absorbance of the resulting supernatant was measured at 532 nm. Measurements were corrected for unspecific turbidity by subtracting the absorbance at 600 nm. The MDA concentration was calculated using the extinction coefficient of 155 mM−1 cm−1. The concentration of MDA was expressed as nmol g−1 FW.
Statistical analysis
Statistical analyses were executed using the programs Statistica 6.0. and Primer of Biostatistics 4.03. Parametric criteria were used since the Shapiro-Wilk's criterion showed that the sampling distribution in all treatments and control did not differ from the Gaussian distribution. One-way ANOVA and parametric Newman-Keuls tests were used for multiple comparisons of studied parameters. Sampling means with standard errors were used for graphical data presentation. The least significant difference was used for multiple comparison at the p<0.05 level between treatment and control means.
RESULTS
Lipid peroxidation
The sublethal concentrations of all pollutants, except for manganese sulfate and glyphosate, caused a significant linear increase in MDA accumulation in the first seedling leaf. In comparison with the control, the MDA content increased by a maximum of 54% (5.12 g L−1 sodium chloride) or 2.0–2.4 times (0.08 g L−1 formaldehyde; 0.08 g L−1 cadmium sulfate; 1.28 g L−1 lead nitrate; 0.32 g L−1 copper sulfate) (Fig. 1a-c, e, g). It is known that many environmental stress factors increase the lipid peroxidation rate in plants (Mittler 2002; Foyer and Noctoc 2005). It is possible that glyphosate and manganese sulfate have a prooxidant effect at concentrations exceeding those studied here.
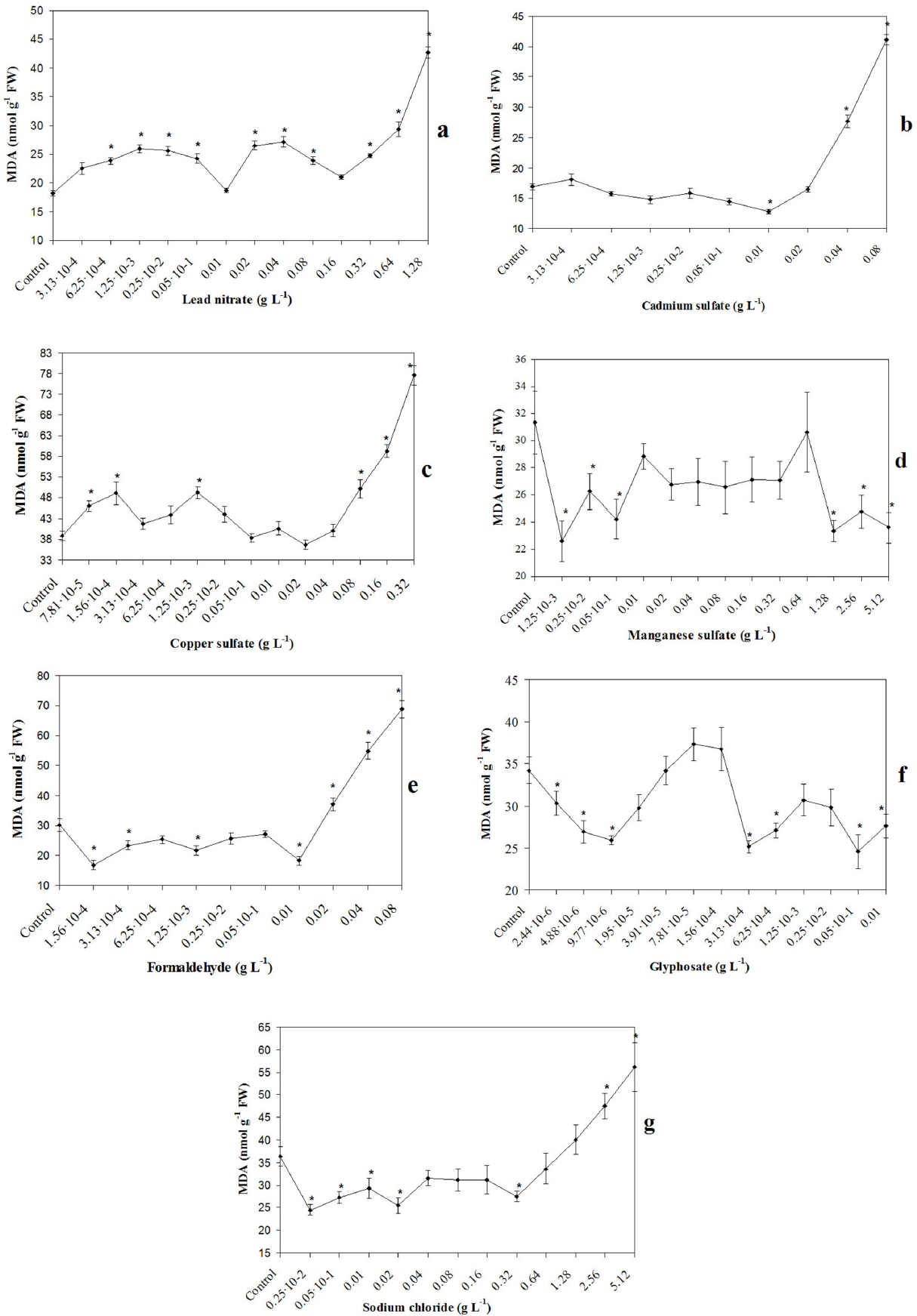
Lipid peroxidation rate in the first leaf of wheat seedlings upon exposure to different pollutants. Values represent mean±SE (n=10). * indicates significant differences between treatments and control at p<0.05.
At the same time, lower toxicant concentrations, except for cadmium sulfate, caused nonmonotonic changes in the MDA content. With an increase in such toxicant concentrations in the solution, there was an alternation in the prooxidant (lead nitrate, copper sulfate) (Fig. 1a, c) or antioxidant (manganese sulfate, sodium chloride, formaldehyde, glyphosate) (Fig. 1d-g) effect (p<0.05 compared to control) with a normalization of the MDA content in the seedling leaf (p>0.05 compared to control). The maximum difference in the MDA content between the treatment and the control was significantly lower than in the range of sublethal concentrations (by 26–48% more or less than control level) (Fig. 1). Apparently, sublethal concentrations caused a disruption in the antioxidant system and therefore an uncontrolled increase in the rate of lipid peroxidation. Regulatory mechanisms prevented a significant disruption in MDA under the action of lower toxicant concentrations.
Analysis of the dose-response dependences for the entire studied range of pollutant concentrations showed the following results. A biphasic hormetic dependence was obtained only for cadmium sulfate. In the first phase of dependence, an antioxidant effect was found. There was a prooxidant effect in the second phase (Fig. 1b). The dose-response dependences of the rest toxicants were multiphasic (three to six phases) and similar to an oscillation. This can be attributed to paradoxical effects. The width of the stimulatory (inhibitory) zone of MDA dependences upon the paradoxical effects and hormesis caused by low pollutant concentrations varied significantly, from one to four points of the curve, with significant differences with compared to the control (p<0.05) & (Fig. 1a-g).
Apparently, we were not able identify oscillatory changes in the MDA content upon exposure to cadmium sulfate because of the very narrow concentration range that caused an antioxidant effect. These low antioxidant concentrations of cadmium sulfate were simply not included in the set of selected doses.
Plant growth parameters
Sublethal concentrations of the studied pollutants, except for the effect of glyphosate on shoot length, caused a significant linear decrease in the size of the shoot and root system. The most significant inhibitory effect was observed for the root system length.
In comparison with the control, the root system length decreased by a maximum of 22–27% (glyphosate, sodium chloride) (Fig. 2f-g) or by 2.1–36.6 times (the rest of pollutants) (Fig. 2a-e), and shoot length decreased by a maximum of 25–35% (salts of lead, copper, manganese, sodium) (Fig. 2a, c-d, g) or by 2.0–4.8 times (cadmium sulfate, formaldehyde) (Fig. 2b, e). It is known that various environmental stress factors can disrupt growth processes in plants (Chapin 1991).

Root system and shoot length of wheat seedlings upon exposure to different pollutants. Values represent mean±SE (n=30). * indicates significant differences between treatments and control at p>0.05.
At the same time, lower toxicant concentrations caused nonmonotonic changes in seedling size. Such an effect was not observed for glyphosate (shoot length) or the salts of cadmium (shoot and root system length) and lead (root system length) (Fig. 2a-b, f). The maximum deviation in growth parameters from the control level was significantly less under the action of low toxicant concentrations compared to a sublethal toxicant content in the solution (for root system length: 6–43%, for shoot length: 5–22%) (Fig. 2).
Analysis of the dose-response dependences for the entire studied range of pollutant concentrations showed the following results. A biphasic hormetic dependence was obtained only under the action of lead salt on the root system length. In the first phase there was a moderate stimulating effect, and in the second phase, a significant inhibitory effect (Fig. 2a). A monotonic dose-response dependence or absent changes were found for cadmium sulfate (shoot and root system length) and glyphosate (shoot length), respectively (Fig. 2b, f). Most dose-response dependences in growth parameters could be attributed to paradoxical effects and were multiphasic with three to six phases (Fig. 2).
The width of the stimulatory (inhibitory) zone of growth parameter dose-response dependences upon the paradoxical effects and hormesis caused by low pollutant concentrations varied significantly from one to five points of the curve, with significant differences with compared to the control (p<0.05) (Fig. 2).
Photosynthetic pigments
Sublethal concentrations of the studied toxicants had a variable influence on the content of chlorophyll and carotenoids. Formaldehyde (0.08 g L−1), copper (0.32 g L−1), and manganese (0.08 g L−1) salts caused a decrease in the level of all pigments (Fig. 3c-e). However, the reduction was not as significant as the change in the MDA content (Fig.1) and seedling size (Fig.2) upon exposure to a sublethal toxicant concentration (only by 24–62% with compared to control) (Fig. 3).
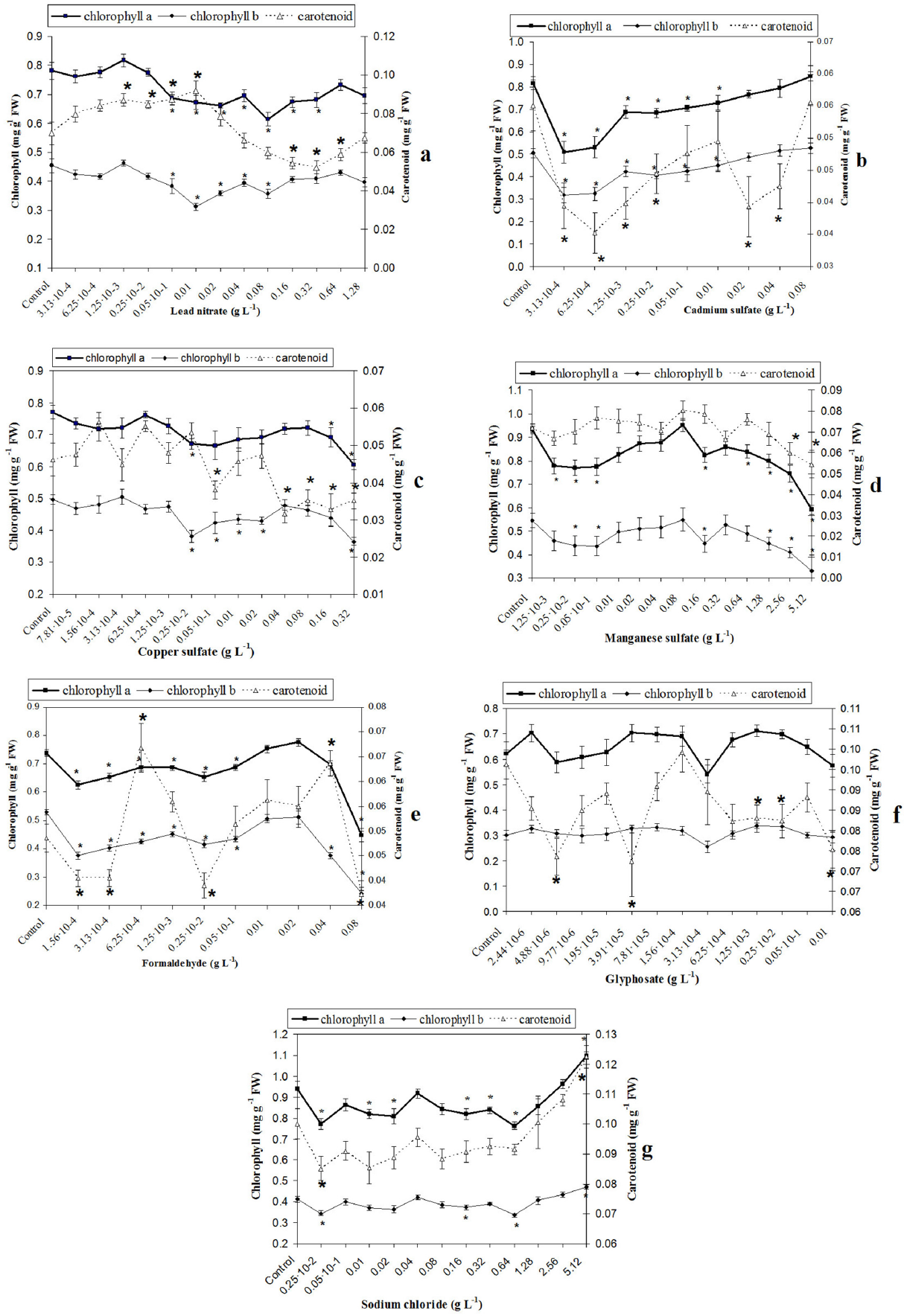
Chlorophyll and carotenoid content in the first leaf of wheat seedlings upon exposure to different pollutants. Values represent mean±SE (n=10). * indicates significant differences between treatments and control at p<0.05.
Glyphosate (0.01 g L−1), lead nitrate (1.28 g L−1), and cadmium sulfate (0.08 g L−1) in sublethal concentrations did not influence on the level of pigments (Fig. 3a-b, f), and sodium chloride (5.12 g L−1) even increased the chlorophyll and carotenoid content (Fig. 3g).
Analysis of dose-response dependences for the entire studied range of pollutant concentrations showed the following results. As a whole, dose-response dependences of the pigment content were biphasic or multiphase (three to five phases) in most cases. At the same time, biphasic dependencies differed from hormesis. In the first phase, the pigment content was less than in the control (p<0.05), while in the second phase, the pigment content increased to the control level (p>0.05). Most dose-response dependences in pigment content could be attributed to paradoxical effects and were multiphasic with three to six phases (Fig. 3).
Only a tendency was found for the existence of multiphase dependences for the chlorophyll and carotenoid content upon glyphosate and manganese sulfate exposure, respectively (p>0.05) (Fig. 3d, f). There was a moderate maximum deviation in the pigment content from control level in the entire range of pollutant concentrations (18–62%).
The width of the stimulatory (inhibitory) zone of pigment dependences upon paradoxical effects caused by low pollutant concentrations varied significantly, from one to eight points on the curve, with significant differences with compared to the control (p<0.05) (Fig. 3).
DISCUSSION
The most frequently seen dose-response dependences of the studied plant parameters on toxicant concentrations were nonmonotonic (85.7% of all dependences). At the same time, the typical biphasic hormetic dependences characterized by low-dose stimulation and high-dose inhibition were only 5.5% of all nonmonotonic responses. These responses were observed under the effect of cadmium sulfate on MDA content (Fig.1b), as well as with salt of lead on the length of the root system (Fig.2a). The rest of the nonmonotonic dose-response dependences could be attributed to paradoxical effects. We identified two types of such responses:
Non-hormetic biphasic dependences (13.9% of all nonmonotonic responses). In the first phase of the dependence, there was an inhibitory effect, while in the second phase, normalization of the parameter or stimulatory effect was seen. This phenomenon was observed under the effect of cadmium and lead salts on the chlorophyll content, as well as with sodium chloride on the level of carotenoids (Fig. 3a-b, g).
Multiphasic dependences similar to oscillations (80.6% of all nonmonotonic responses). These responses were observed the most frequently under the action of toxicants on MDA accumulation, carotenoid content, and growth parameters and had three phases or more (Fig. 1 –3).
Hormesis could be part of some multiphasic dose-response dependences (i.e. part of the paradoxical effect). For example, some parts of the multiphasic dependences of the MDA content to sodium chloride (0.16−5.12 g L−1) and formaldehyde (0.05·10−1−0.08 g L−1) concentrations were hormesis (Fig. 1e, g). In the first phase, toxicants induced an antioxidant effect (0.16–0.32 g L−1 sodium chloride; 0.05·10−1−0.01 g L−1 formaldehyde), while in the second phase a prooxidant effect was seen. Hormesis was also observed under the effect of glyphosate (1.56 10−4−0.01 g L−1) and sodium chloride (0.16−5.12 g L−1) on the root system length (Fig. 2f-g). At first, these toxicants had a stimulatory effect, and then an inhibitory one. In general, these dose-response dependences were multiphasic.
Thus, paradoxical effects and hormesis, apparently, are interrelated phenomena as they can be parts of the same complex oscillatory dependence. Perhaps researchers usually identify mainly biphasic hormetic dependences because a relatively narrow range of toxicant doses is studied. We analyzed a range from sublethal concentrations of toxicants to values in 102–105 times smaller, so mainly multiphasic dependences were identified (i.e. paradoxical effects).
Nonmonotonic changes in different seedling parameters were not always synchronous under the influence of a certain toxicant. The same toxicant concentrations caused a normalization of some parameters and a disruption of others. For example, high concentrations of cadmium sulfate (0.08 g L−1), lead nitrate (1.28 g L−1), and sodium chloride (5.12 g L−1) caused a significant destabilization of the seedling state (a significant increase in the lipid peroxidation rate and inhibition of growth) and normalized or even increased the pigment content (Fig. 1a-b, g; Fig. 2a-b, g; Fig. 3a-b, g). Apparently, the adaptation of seedlings to high toxicant concentrations required considerable energy expenditure. Therefore, the maintenance of photosynthesis (including the content of photosynthetic pigments) as an energy source was necessary for the survival of plants in extreme conditions.
The studied parameters can have different type of dose-response dependences under the action of the same toxicant. For example, lead nitrate induced a biphasic change in the content of chlorophyll (non-hormetic dependence) and the root system length (hormesis), and a multiphasic change in MDA and carotenoid content (Fig. 1a; Fig. 2a; Fig. 3a).
An asynchronous change in studied seedling parameters may be caused by the specificity of the toxicant, as well as by its significance for the phenotypic adaptation of plant. For example, the MDA content had the most multiphasic dependences as compared with the rest of the parameters (Fig.1). The follow medians for phase number were found: Me(MDA)=5; Me(chlorophylls)=3; Me(carotenoids)=3; Me(length of root system)=4; Me(shoot length)=3. It is known that some products of lipid peroxidation such as MDA are biologically active and take part in phenotypic adaptation to stress factors in plants (Brand et al. 2004; Weber et al. 2004). It is possible, therefore, that the MDA content is the most labile parameter of wheat seedlings.
It should be noted that significant deviation in studied parameters from the control level occurred only under the action of sublethal concentrations of toxicants. For example, such concentrations increased the MDA content to 2.0–2.4, and the length of the root system was reduced to 2.1–36.6 times as compared with the control. At lower concentrations causing oscillatory responses, the deviation from the control level up or down was moderate (not more than 62%). Other authors have also shown that a moderate stimulatory response (exceeding control not more than 60%) is most typical for hormesis (Calabrese 2008). Apparently, this is a universal regularity for any nonmonotonic responses under the influence of low toxicant doses.
Significant disruptions in the organism's state are harmful for its normal functioning. Therefore, significant disruptions are prevented by regulatory systems. So long as the organism is able to normalize its parameters, changes will be moderate. Under the action of sublethal toxicant concentrations, the compensatory capacity of the organism is depleted and significant impairment of its parameters is observed.
The width of the stimulatory (inhibitory) zone of pigment, MDA, and growth parameter dependences upon hormesis and paradoxical effects caused by low pollutant concentrations varied significantly, from one to eight points on the curve, with significant differences compared to control. Significant variation in the width of the stimulatory zone was shown for hormesis by Calabrese (2008).
At present, the causes of a nonmonotonic response in living organisms are insufficiently explored. Previously, to explain this phenomenon, we proposed a hypothesis (Erofeeva et al. 2011) that was based on the concept of the gradual involvement of different adaptive mechanisms in the process of phenotypic adaptation to a damaging factor and the existence of several regimes (levels) of functioning in living organisms (Veselova et al. 1998; Garkavi et al. 1999). Each level has its own adaptive capacity. If the intensity of the damaging factor exceeds the adaptive capacity of this functioning level, homeostasis is disturbed, and additional protective mechanisms are activated. As a result, the organism transits to the next level of functioning. This transition allows for a relative normalization of homeostasis, but markedly increases resource expenditures for adaptation. With a further increase in the intensity of the factor, a disturbance in the organism's state will be observed again, which will cause a transition to the next level with more active adaptive mechanisms. Therefore, increasing the intensity of a factor can cause an oscillatory change in organism parameters, in which the deviation of the parameter from the control will alternate with normalization.
Under the influence of sublethal factor values, the adaptive capacity of the organism will be depleted, so a significant increase in parameter disturbance will be observed and the dose-response dependence will be linear.
If our hypothesis is correct, all the parameters of the organism related to phenotypic adaptation must have oscillatory dependences on the intensity of a stress factor. Such oscillations can have a different period, amplitude and direction of the effect (stimulatory or inhibitory effect), depending on the significance of the plant parameter for the adaptation process and the specificity of the stress factor.
We can draw the following conclusions based on this study:
Under the action of different pollutants (salts of cadmium, copper, manganese, lead as well as formaldehyde, the herbicide glyphosate and sodium chloride) in a wide concentration range from sublethal values to 102–105 times lower concentrations, the majority the dose-response dependences of MDA, pigment content, and growth parameters were nonmonotonic (85.7% of all dependences). Of these, 5.5% were hormesis and paradoxical effects comprised 94.5%. Multiphasic dose-response dependences similar to an oscillation were the most widespread type of paradoxical effects.
Hormesis was a part of some multiphasic responses (i.e. paradoxical effects), which indicates a relationship between these phenomena.
Sublethal pollutant concentrations significantly increased lipid peroxidation (by 2.0–2.4 times, except for manganese and glyphosate) and decreased growth parameters (by 2.1–36.6 times, except for shoot length upon glyphosate exposure), while the photosynthetic pigment content reduced insignificantly, normalized or even increased. Lower pollutant concentrations caused a moderate deviation in all parameters from the control (not more than 62%) for hormesis and paradoxical effects.
The studied seedling parameters could have different types of nonmonotonic dose-response dependences (hormesis or paradoxical effects) upon the same pollutant exposure.
The width of the stimulatory (inhibitory) zone of pigment, MDA, and growth parameter dependences upon hormesis and paradoxical effects caused by low pollutant concentrations varied significantly from one to eight points of the curve, with significant differences compared to control (p<0.05).
Footnotes
ACKNOWLEDGMENTS
Three anonymous reviewers gave valuable comments on earlier versions of the manuscript.