
Abstract
Radiations may trigger protective response within a threshold of doses applied. Exposures above an upper threshold are generally detrimental, while exposures below a lower threshold may or may not increase risks for health. We recently reported that a cellular protective response occurs in interventional cardiologists to counteract the oxidative damage caused by radiation. Here, we demonstrated in an in vitro model represented by whole blood of healthy donors γ-irradiated with 220–440 mGy, that haemolysis of erythrocytes induced by hypochlorous acid was reduced by 40%. The protection triggered by γ-radiations made erythrocytes more resistant to oxidative insult caused by hypochlorous acid which was induced 3 h after irradiation and involved biochemical changes in the synthesis and turnover of glutathione. Overall, the biochemical remodelling induced by exposure to γ-radiations might contribute to generate new guidelines in professionally exposed workers.
INTRODUCTION
According to BEIR VII report (BEIR VII 2006), the maximal permissible levels that are recommended in the United States by the National Council on Radiation Protection and Measurements (NCRP) for people exposed to radiation other than background radiation and from medical applications are 1 mSv/year for the general population and 50 mSv per year for professionally exposed workers. Considering that much of the epidemiology of low dose exposures includes people who in their lifetime received up to 500 mSv, the BEIR VII committee evaluated the biological effects at low doses of low-LET (linear energy transfer) radiations in the range not exceeding 100 mGy (BEIR VII 2006). Accordingly, in relation to stochastic health risks, for sparsely ionising radiations, a low dose is defined as <100 mGy delivered acutely and a low dose-rate as <5 mGy per hour (Wakeford and Tawn 2010). In terms of “risk models”, the linear no-threshold model (LNT) predicts that approximately 1 person in 100 would be expected to develop cancer from a dose of 0.1 Sv above background, while lower doses would produce proportionally lower risks, e.g. one individual per thousand would develop cancer from an exposure to 0.01 Sv. The LNT hypothesis is strongly debated since a wealth of data from experiments using single and multi-cellular organisms, including mammals, reached different conclusions (reviewed in Calabrese 2009, Calabrese et al. 2008, Mitchel 2006, Vaiserman 2010). Following this view, it has been proposed that damage or protection induced by radiations in mammalian cells and mammals operates within an upper and lower dose thresholds, typically between about 1 and 100 mGy for a single low dose rate exposure. Exposures above the upper threshold are generally detrimental, while exposures below the lower threshold may or may not increase either cancer or non-cancer disease risk (Mitchel 2010).
Recently, correlations between the redox state of cells and protection against radiation damage have been increasingly reported. Constantly increased levels of reactive oxygen species (ROS) or nitric oxide (NO) have been observed in adapted cells and both factors may play a role in the maintenance process (Matsumoto et al. 2007, Tapio and Jacob 2007). In this context, we demonstrated that a cellular biochemical adaptation occurs in interventional cardiologists to counteract the oxidative damage caused by radiation (Russo and Picano 2012, Russo et al. 2012). We reported that the 3-fold increase in circulating ROS in exposed subjects was balanced by an increased GSH concentration in erythrocytes. A similar event probably occurred in lymphocytes of the same cardiologists, where an increased basal level of caspase-3 was measured and interpreted as a predisposition to a more efficient apoptotic response aimed to remove potentially malignant somatic cells (Russo and Picano 2012, Russo et al. 2012). This protective mechanism may avoid that prolonged radiation exposure could induce genetic damage (Andreassi et al. 2005). In the cardiologists’ study, the lifetime exposure ranged between 20 to 100 mSv (from 1,000 to 5,000 chest X-rays), but the population of exposed subjects was healthy, despite the level of exposure, suggesting that interventional cardiologists might activate protective mechanisms triggered by radiation themselves (Russo and Picano 2012, Russo et al. 2012).
To further explore at molecular level this recent finding, we used human erythrocytes to established an in vitro model to assess the biological properties of radiations administered at doses not much different from those received following professional exposure. As a consequence of their physiological role, erythrocytes are exposed to continuous oxidative stress deriving from endogenous (Cimen 2008), or exogenous sources, as in the case of professional exposure to radiation. In this respect, it has been suggested that erythrocytes and their precursor cells may represent a lifelong, wide-range radiation biodosimeter (Gong et al. 1999). For these reasons, red cells have developed efficient enzymatic and not-enzymatic antioxidant defences to preserve themselves from oxidative damage. In addition, due to their mobility, erythrocytes can be considered as ideal antioxidant scavengers acting throughout the circulation and reducing the damaging mass effect of large quantities of ROS on different tissues (Russo et al. 2012).
In the present work, whole blood samples collected from healthy donors were in vitro γ-irradiated at doses relatively low to evaluate the effects on hypochlorous acid-induced haemolysis in isolated erythrocytes.
MATERIALS AND METHODS
Reagents
Glutathione (GSH); phtaldialdehyde, trichloroacetic acid (TCA); hydrogen peroxide (H2O2); phenazine methosulfate (PMS); nitro blue tetrazolium (NBT); β-nicotinamide adenine dinucleotide 2′-phosphate reduced tetrasodium salt hydrate (β-NADPH); 2′,7′-dichlorofluorescein diacetate (DCFH-DA); N-acetyl-L-cysteine (NAC); iodoacetamide (IAC) were from Sigma-Aldrich Chemical Co (Milan, Italy); sodium hypochlorite (NaOCl) was from Carlo Erba (Milan, Italy); phosphate-buffered saline (PBS) tablets were purchased from Life Technologies (Monza, Italy). All other chemicals used were of research highest purity grade.
NaOCl was diluted with PBS, and the pH of the solution was adjusted to 7.4 immediately before use. At this pH, the solution contains approximately equimolar amounts of HOCl (hypochlorous acid) and NaOCl, and is referred to hereafter as HOCl (Vissers et al. 1998).
Sample preparation and treatment condition
Whole blood samples were obtained from healthy donors, collected in EDTA-treated tubes and immediately used. Aliquots (1 ml) were γ-irradiated under air at a final dose of 220 or 440 mGy. The source of irradiation was a 137Cs-γ-radiation at room temperature, in the atmosphere of air (Gamma Cell Elite 2.7 Gy/min, MDS Nordion Canada). Irradiated and non-irradiated samples were incubated for 3 h at 37° C. After incubation, 0.1 ml of each sample was immediately incubated with 5 mM of HOCl at 37° C for 15 min.
To verify the role of GSH in erythrocyte protection after γ-irradiation, whole blood samples were pretreated with IAC (10 mM) for 30 min and exposed to 440 mGy; after 3 h, HClO (5 mM) was added to samples for 15 min before measurement of hemolysis and GSH concentration. Samples treated with different concentration of NAC (10=80 mM) were incubated for 3 h before determination of GSH.
Determination of haemolysis
After γ-irradiation, aliquots of whole blood samples (0.1 ml), were diluted to 1 ml with PBS and centrifuged at 4000 rpm for 4 min. After centrifugation, supernatants were collected and diluted with PBS. Absorbance was spectrophotometrically measured at 540 nm and the percent of haemolysis calculated as reported (Tedesco et al. 2000).
GSH measurement
GSH determination was evaluated by a spectrofluorimetric method. After treatment, erythrocytes were isolated from whole blood by centrifugation. Supernatants containing plasma and “buffy coat” were eliminated; subsequently, three consecutive washes with PBS allowed removal of residual plasma and leukocytes (Tedesco et al. 2001, Tedesco et al. 2000). Pellets were washed with PBS and proteins were precipitated with TCA (5% v/v final concentration in 0.1 M HCl and 10 mM EDTA). Using phthaldialdehyde as a substrate, the fluorescence of the supernatant was measured at 340 nm (excitation wavelength) and 460 nm (emission wavelength) (Russo et al. 2012). The concentration of GSH was extrapolated from a standard curve for pure GSH and it was expressed as percent of untreated (non-irradiated samples).
Fluorescent measurement of intracellular ROS
Irradiated and non-irradiated blood samples were incubated for a maximum of 3 h at 37° C in the atmosphere of air. At the end of incubation 0.1 ml of blood was centrifuged at 4000 rpm for 10 min and the erythrocytes washed with PBS. Red blood cells were suspended with PBS and incubated 30 min in the presence of 10 μM DCFH-DA, a non-fluorescent compound that freely permeates cells. DCFH-DA is hydrolyzed to DCFH, whose interaction with peroxide gives rise to 2′,7′-dichlorofluorescin (DCF) detected spectrofluorimetrically. Then erythrocytes were centrifuged and washed with PBS. Aliquots were transferred in a multiwells (Costar) and the ROS production was measured using a microplate fluorescence reader (FL 500 Bio-Tek Instruments Milan, Italy) with excitation and emission settings at 485 and 530 nm, respectively (Tedesco et al. 2000).
RESULTS AND DISCUSSION
It has been extensively reported that free electrons generated by γ-radiations can hit and damage cellular structures both directly, or indirectly, by interacting with water molecules and producing ROS (Feinendegen 2002), such as the high reactive hydroxyl radical, OH• which, in turns, produces hydrogen peroxide. In the attempt to reconstitute in vitro the potential protective effects of γ-radiations on the redox homeostasis of human erythrocytes, we treated whole human blood from healthy donors with HClO, which results in rapid oxidation of reduced glutathione, increase in cell osmotic fragility and formation of transient membrane pores with the consequent haemolysis (Zavodnik et al. 2002). We applied a relatively high concentration of HClO (5 mM) which caused an average haemolysis of about 15–20% (15.74 ± 2.22; Fig. 1). The rationale for using such concentration was the following: i. we used whole blood instead of isolated erythrocytes; therefore, the HClO concentration applied was in the range required to bypass the antioxidant capacity of human plasma (data not shown); ii. hyperproduction of HClO in the millimolar range is normally generated by neutrophils and monocytes following inflammatory tissue injury and microbial killing (Jesaitis and Dratz 1992). As reported in Fig. 1, γ-irradiation (220 – 440 mGy) protected in a dose dependent manner human erythrocytes from HClO-induced haemolysis. Blood samples irradiated with 440 mGy, showed a protection of about 40% (9.56 ± 1.70 vs 15.74 ± 2.22). It is worthwhile to note that the maximal effect was observed after 3 h from γ-irradiation (data not shown), confirming the need of a lag time to allow the erythrocytes to develop radiation-induced resistance to the oxidative insult caused by HClO.
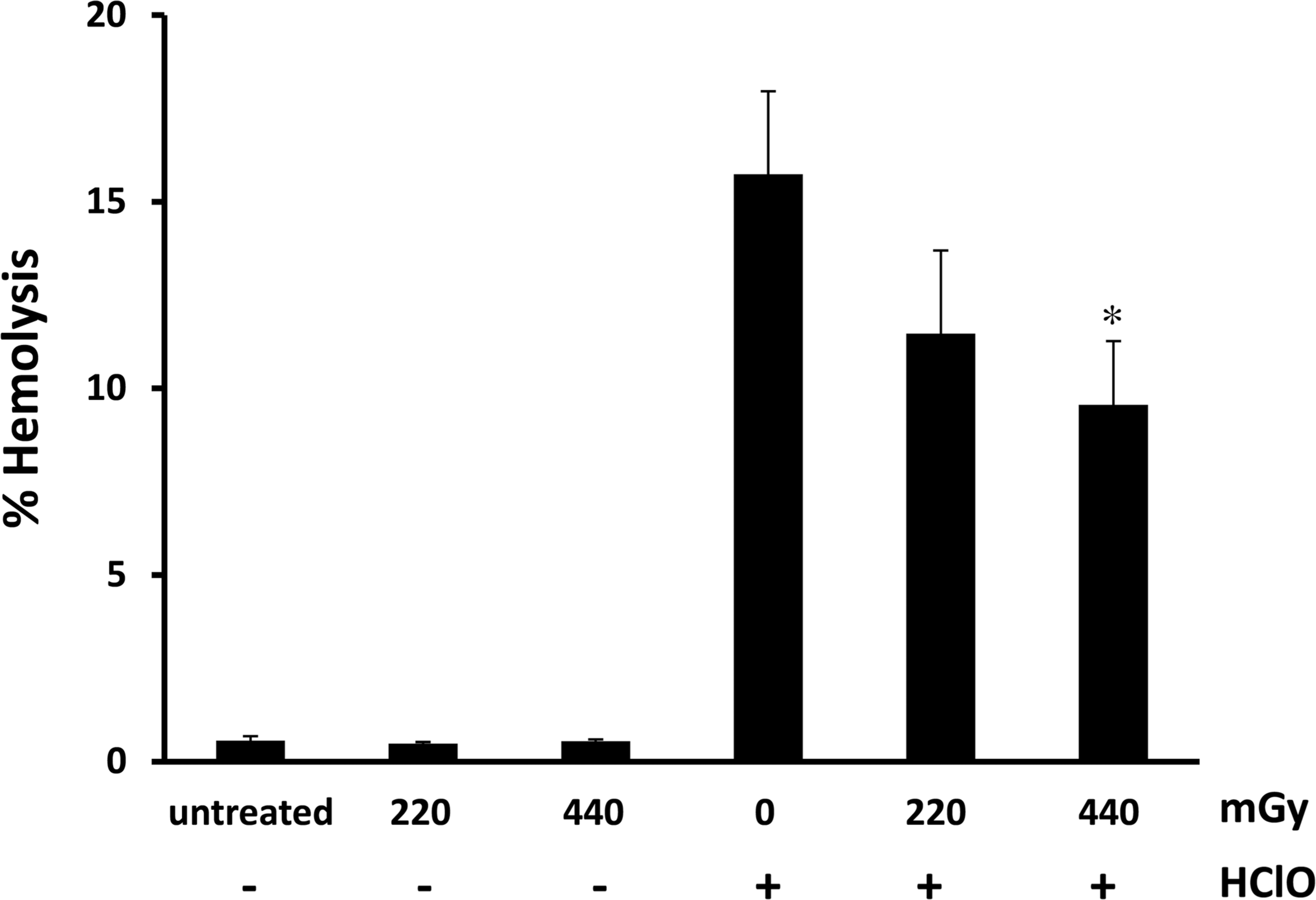
γ-Irradiations attenuates haemolysis in erythrocytes treated with HClO. Whole blood samples of 9 donors were irradiated at indicated doses and incubated for 3 h at 37° C. Subsequently, samples were treated with HClO (5 mM) for 15 min at 37° C and percentage of haemolysis measured as reported in method section. Bar graphs represent the mean ± s.e. Symbols indicate significance: p<0.05 (∗) vs HClO mono treatment.
Since ionizing radiation generates the production of ROS, such as superoxide, hydrogen peroxide and hydroxyl radical in a variety of cells, including erythrocytes (Klucinski et al. 2008, Riley 1994), we verified if this was also the case in our experimental model. As shown in Fig. 2, percentage of ROS production almost doubled compared to control (196.56 ± 18.21), soon after irradiation of whole blood with 440 mGy and returned to basal level after 3 h. At this time, maximal protection by γ-radiations was observed (Fig. 1). Therefore, we postulated that the transient ROS increase following γ-irradiation triggered antioxidant defences leading to an increased cellular resistance to a second oxidative damage induced by HClO.
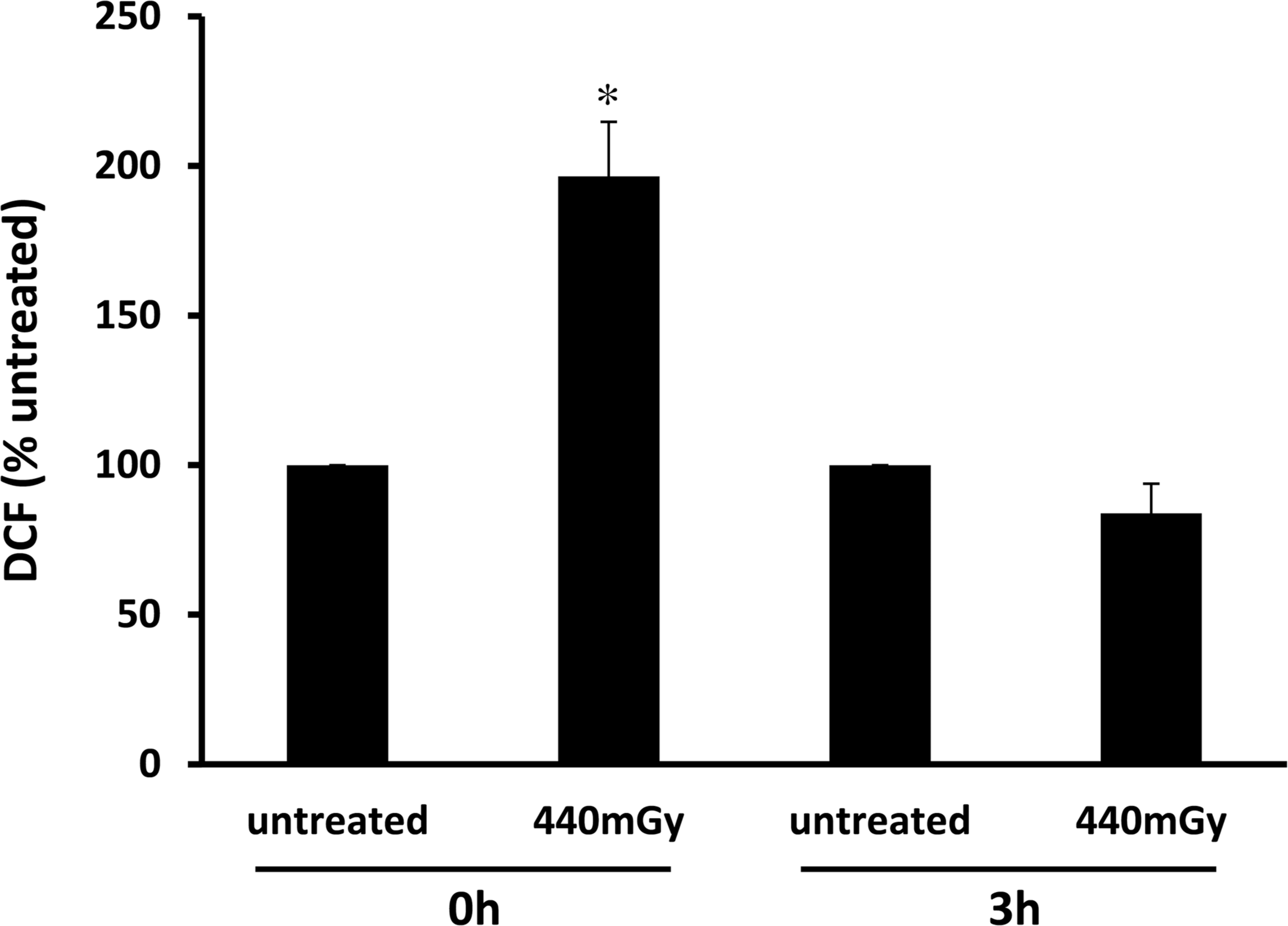
γ-Radiations transiently increase intracellular ROS. Whole blood from healthy donors was exposed to 440 mGy and intracellular ROS were measured at different time as reported in methods section. Bar graphs represent the mean (expressed as percent of untreated) ± s.e. Symbols indicate significance: p<0.05 (∗) versus untreated.
Based on previous observation carried out on workers of X-ray departments occupationally exposed to long-term low levels of ionizing radiation where a significant decrease of erythrocyte enzymatic antioxidant defences (SOD and CAT) was observed (Klucinski et al. 2008), we measured SOD and CAT activities to verify if increase in the activities of these antioxidant enzymes was able to counteract ROS burst observed in Fig. 2. However, none of them showed a significant variation in after treatment with γ-radiations (data not shown). On the opposite, we observed a significant and dose-dependent increase in the percentage of GSH compared to untreated samples following treatment at both 220 and 440 mGy (202 ± 76 and 247 ± 65, respectively; Fig. 3). Therefore, we tried to investigate the role of GSH in triggering biochemical protection induced by γ-radiations in erythrocytes using IAC and NAC, two compounds able to regulate GSH concentration in erythrocytes. IAC is an alkylating agent that bind covalently with the thiol group of cysteine inducing a depletion of GSH (Palmen and Evelo 1996). NAC is a synthetic precursor of intracellular cysteine and GSH and its administration replenishes intracellular GSH levels (Atkuri et al. 2007). Fig. 4A reports that treatment of blood samples with 10 mM IAC strongly decreased basal level of GSH in not irradiated (29.90 ± 29.89) and irradiated erythrocytes (3.09 ± 3.10 vs 247 ± 65). As a consequence of GSH depletion, HClO-induced haemolysis in the presence of IAC almost doubled (28.66 ± 5.57 vs 15.07 ± 1.93) and the protective effect of γ-irradiation disappeared (29.19 ± 5.06 vs 9.42 ± 0.70) (Fig. 4B). On the opposite, increasing concentrations of NAC were able to increase GSH production in a dose dependent manner (Fig. 5A) and, consequently, to lower haemolysis induced by HClO (Fig. 5B).

Effects of γ-radiations on GSH concentration. GSH content was measured after exposition of whole blood from 9 donors to 220–440 mGy and compared to untreated samples. After 3 h of incubation, haemolysates were prepared and GSH concentration was measured spectrofluorimetrically at 340±20 nm of excitation and 460±20 nm emission using phthaldialdehyde as a substrate. Bar graphs represent the mean (expressed as percent of untreated) ± s.e. Symbols indicate significance: p<0.05 (∗) respect to untreated.

IAC sequesters GSH making erythrocytes more sensitive to haemolysis induced by HClO. Samples in panel A were prepared as follow: whole blood was pretreated with IAC (10 mM) for 30 min and exposed to 440 mGy. After 3 h, haemolysates were prepared and GSH concentration was measured spectrofluorimetrically. Bar graphs represent the mean (expressed as percent of untreated) ± s.e. Symbols indicate significance: p<0.05 (∗) respect to untreated and p<0.001 (∗∗) respect to untreated and 440 mGy irradiation. In panel B, the whole blood was treated as indicated. IAC incubation (30 min) preceded exposition to 440 mGy. After 3 h, samples were treated with HClO (5 mM) for 15 min at 37° C followed by measurement of haemolysis. Bar graphs represent the mean ± s.e. Symbols indicate significance: p<0.05 (∗) respect to HClO mono-treatment, p<0.01 (∗∗) respect to 440 mGy plus HClO and p<0.005 (∗∗∗) respect to HClO mono-treatment.

NAC enhances GSH levels protecting erythrocytes from haemolysis induced by HClO. In panel A, whole blood was treated with different concentration of NAC (10=80 mM) and GSH content was determined after 3 h of incubation. Bar graphs represent the mean (expressed as percentage of untreated) ± s.e. Symbols indicate significance: p<0.05 (∗) and p<0.005 (∗∗) versus untreated samples. In panel B, whole blood was treated as reported in panel A before addition of 5 mM HClO for 15 min at 37° C. Pellets were extensively washed with PBS to remove NAC before GSH measurement. The percentage of haemolysis was measured as reported in methods section. Bar graphs represent the mean ± s.e. Symbols indicate significance: p<0.05 (∗) versus HClO mono treatment.
We summarized the data obtained in the present work in the scheme reported in Fig. 6. Shortly, our hypothesis suggests that treatment of whole human blood with a HClO concentration not far from that reachable in vivo following an inflammatory process, induces large haemolysis in erythrocytes caused by a burst of intracellular ROS. Pre-treatment of whole blood with ionizing radiation at doses < 500 mGy generates a protective effect against HClO-induced haemolysis. We postulated that the initial and transient increase in intracellular ROS caused by γ-irradiation triggers downstream events leading to increased GSH concentration which, in turns, protects erythrocytes from the subsequent oxidative damage caused by HClO (Fig. 6). How ROS-generated by γ-irradiation positively regulate de novo GSH synthesis is actually under investigation in our laboratory. A model for GSH synthesis and turnover in the human erythrocyte has been recently published and represents the starting point for future studies (Raftos et al. 2010).
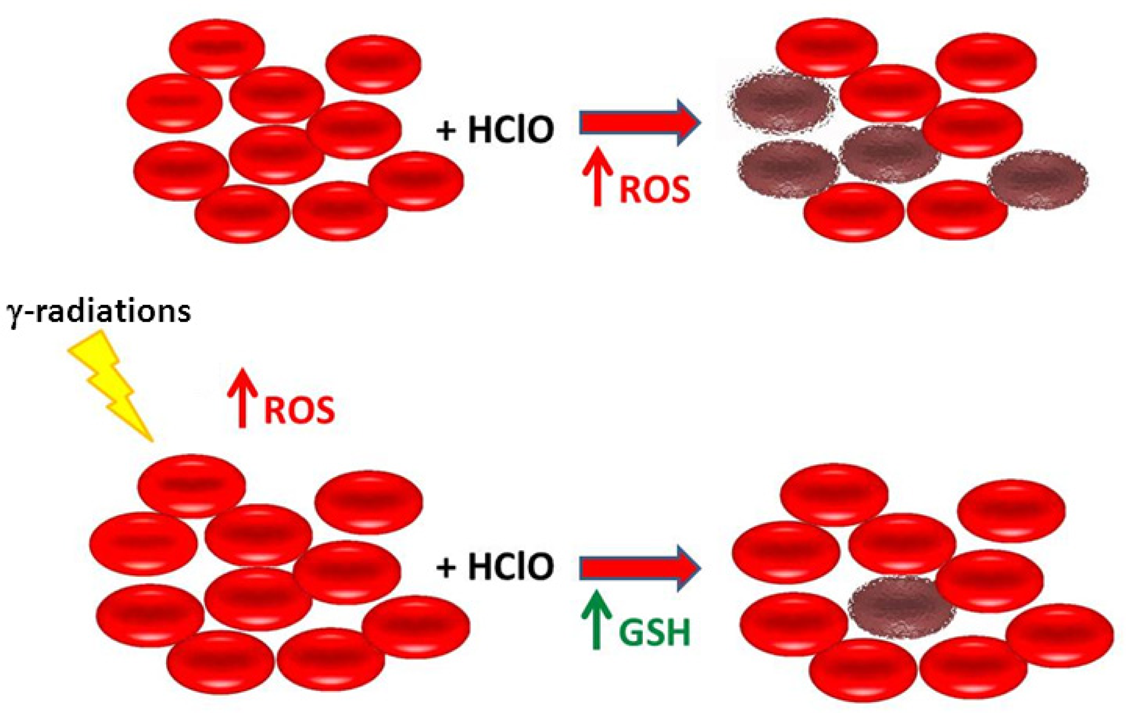
Schematic representation of the effect of γ-radiations in erythrocytes treated with HClO accordingly to data reported in the text.
A previous study indicated that NaOCl-induced haemolysis of erythrocytes pretreated with X-radiation at a dose of 400 Gy was lower than haemolysis of erythrocytes treated only with NaOCl. In contrast, slightly higher haemolysis was observed for erythrocytes pre-irradiated with lower (40 or 200 Gy) doses of radiation (Krokosz 2003). However, substantial differences exist between the present work and the Krokosz's study (Krokosz 2003): i. we applied a dose of radiation at least three orders of magnitude lower (440 mGy versus 400 Gy); ii. we irradiated whole blood which appears a more physiological exposure to radiations than isolated erythrocytes; iii. we demonstrated that GSH plays an important role in the protective effect of γ-irradiation against oxidative damage induced by HClO. In addition, the in vitro model investigated in the present work strongly supports the observation recently published on the capacity of erythrocytes isolated from a group of interventional cardiologists to generate a biochemical protective response able to compensate the potential damage associated to professional exposure to ionizing radiations (Russo and Picano 2012, Russo et al. 2012). In fact, in interventional cardiologists: i. the increase in circulating ROS was balanced by an increased GSH concentration in erythrocytes, while no significant changes in SOD and catalase activities were measured; ii. the total serum antioxidant power in exposed cardiologists and control groups did not show any significant change, suggesting the generation of a possible protective mechanisms. Similarly, in our model, 3 h after irradiation and before HClO addition, no change in serum antioxidant power was measured (data not shown).
In summary, the present work suggests that relatively low doses of ionizing radiation may induce biochemical changes in the synthesis and turnover of GSH which makes erythrocytes more resistant to further oxidative insult. Overall, the biochemical remodelling induced by γ-irradiation might contribute to generate new guidelines in professionally exposed workers, which are currently based, almost exclusively, on minimizing radiation exposure by implementing safety measures and protection devices.
Footnotes
ACKNOWLEDGMENT
We thank the healthy subjects who gave their consent for blood samples. Conflict of interest: none declared.