
Abstract
An adaptive response may be defined as the effect of a small priming dose of radiation modifying the anticipated cellular response of the same tissues so as to alter the predicted response to a larger dose of radiation. We and others have demonstrated that at low radiation doses (less than 0.5 Gy) the lethal and mutational effect of the radiation is mainly, possibly entirely, due to the non-targeted effects. This is the dose range for priming doses in adaptive response protocols. In an associated presentation from our group, we demonstrate that the adaptive response may be explicable as a non targeted (bystander) response. In this paper we present data from exposed human patients, showing that a simple assay using blood can demonstrate variation in the extent and type of non-targeted effects and that exposure to radiation can modulate the subsequent non-targeted response to a later dose. Patients undergoing radiotherapy treatment for cancer gave blood samples immediately after the first dose, midway during and six weeks after therapy. The serum from these samples was harvested, diluted in tissue culture medium and added to reporter cells. The toxicity or growth promoting activity of the serum was measured using a clonogenic assay coupled with immunocytochemical measurement of various proteins involved in apoptosis or growth. There is already evidence that bystander effects are controlled by both genetic and epigenetic (lifestyle) factors. These data could support the development of a simple blood based assay to predict overall response of human subjects to low doses of radiation taking all the low dose factors into account.
I. INTRODUCTION
Radiation has been referred to in somewhat melodramatic fashion as the invisible killer. It is regarded as invisible because it cannot be sensed in any way. It is regarded as a killer because of the generally accepted analysis of, and extrapolation from, the fatalities of Hiroshima and Nagasaki.
This “invisibility” means that persons exposed to low doses do not necessarily know they have been exposed. Moreover, if they were exposed, they are unaware of how long they were exposed for. It would obviously be helpful to have a biological dosimeter within the patient that could be interrogated to reveal the nature and duration of the radiation exposure.
This has been worked on for a number of years, under the prevailing radiation paradigms. These paradigms are that there are target sites within the cell for radiation action, and that increasing dose causes a proportional increase in effect. These paradigms are encapsulated in the linear no-threshold model of radiation action, which extrapolates risk from high dose acute exposure.
It is implicit within this model that the primary lesion caused by radiation is the DNA double strand break, and that these breaks are produced in proportion to radiation dose. Theoretically, therefore, any dose of radiation can cause a double strand break, and therefore there cannot be a dose of radiation that is risk free.
This approach is now being challenged by the emergence of a whole body of data, which suggests that at low doses, the dose is not as important as the response to that dose. 1, –4 Bystander effects, genomic instability and adaptive responses all mean that the LNT hypothesis is too simple to explain low dose responses. The responses appear to be both genetically and epigenetically determined, and response does not seem to increase with dose in any clear way. 5,6
A whole new approach is needed where the emphasis is on response and not dose. The temporal and spatial aspects of response are also critical. The ideal biological endpoint will also distinguish between effect and harm.
Previous data from our group showed that tissue, harvested from human or mouse sources and irradiated in vitro, showed considerable variation in the degree to which bystander and genomic instability effects were expressed. 7, –10 A clear genetic component can be shown using mice, 7,8 and an epigenetic component can be demonstrated by analyzing data from smokers separately. 8,–10 These data lead us to suspect that a more general in vivo test using blood might be possible.
II. THE NEW APPROACH
As an approach towards examining these issues, we have been taking blood from radiotherapy patients. These patients are exposed daily to doses of around 2.0 Gy, to a localized treatment area. Treatment times vary according to machine type, but are typically around two minutes. During this time, circulating lymphocytes will receive part of this dose as they travel through the circulatory system. It is unusual for any areas of bone marrow production to be directly irradiated, but both they and circulating lymphocytes will receive a certain amount of scattered radiation.
The blood from the patients is centrifuged at 1500 rpm in a bench centrifuge, and the serum is aspirated off. 0.5 ml of this is added to flasks (in triplicate) containing 300 human keratinocyte cells, pre-seeded in 5 ml of tissue culture medium, six hours previously. Controls are unirradiated human serum samples and culture medium from irradiated and control keratinocytes. The flasks are incubated at 37°C in a humidified incubator, in an atmosphere of 5% CO2 in air for nine days. The flasks are then stained with carbol fuchsin stain, and colonies are counted using the Puck and Markus clonogenic assay technique. The clonogenic survival is the number of cells that form colonies and is determined by calculating the number of colonies divided by the original number of plated cells. This is corrected for the control plating efficiency.
This assay effectively measures the toxicity of the serum from irradiated patients towards unirradiated cells and is a bystander toxicity index (BTI), which we define here as the toxicity of the treated sample versus the control. An index less than one means the overall effect is toxic. If the index is greater than one, the bystander effect causes growth or enhanced clonogenic survival.
III. PRELIMINARY RESULTS AND DISCUSSION
Figure 1 shows the preliminary results for the first 19 patients. It is clear there is very large individual variation in response. But this probably is due to the low patient numbers at this stage. At present, since the patient details are coded to preserve objectivity, we have not analyzed all the data regarding clinical radiosensitivity of these patients, but have accumulated these data together with fatigue scores for the patients as a generic measure of systemic response.

Toxicity of serum from radiotherapy patients Samples were taken at the start of radiotherapy after the first dose had been administered. “N” is a mean control value for ten normal unirradiated volunteers without cancer.
Figure 2 shows the results obtained for a small subset of patients with very advanced cancers. Three patients had primary colon tumours and one had primary breast. They are linked by all having advanced metastatic tumour spread. Clearly, their BTI is greater than one, meaning that the bystander effect in this instance is promoting proliferation.

Lack of toxicity of serum harvested from four patients who had very large tumours.
In Figure 3, data for a group of patients where three blood samples were tested are shown. Samples were taken immediately after the first dose, midway during therapy and six weeks after radiotherapy. Again there is great individual variation but the trend is for the bystander effect either to decrease during therapy in some patients or to decrease during therapy in others. A bell shaped response only occurred in one patient. Clearly again, more patient data are needed. The aim of the paper is to show the possibilities of this assay not to draw conclusions from this limited amount of data. The data are, however consistent with other data from our group suggesting an adaptive response to this endpoint of radiation exposure. 11
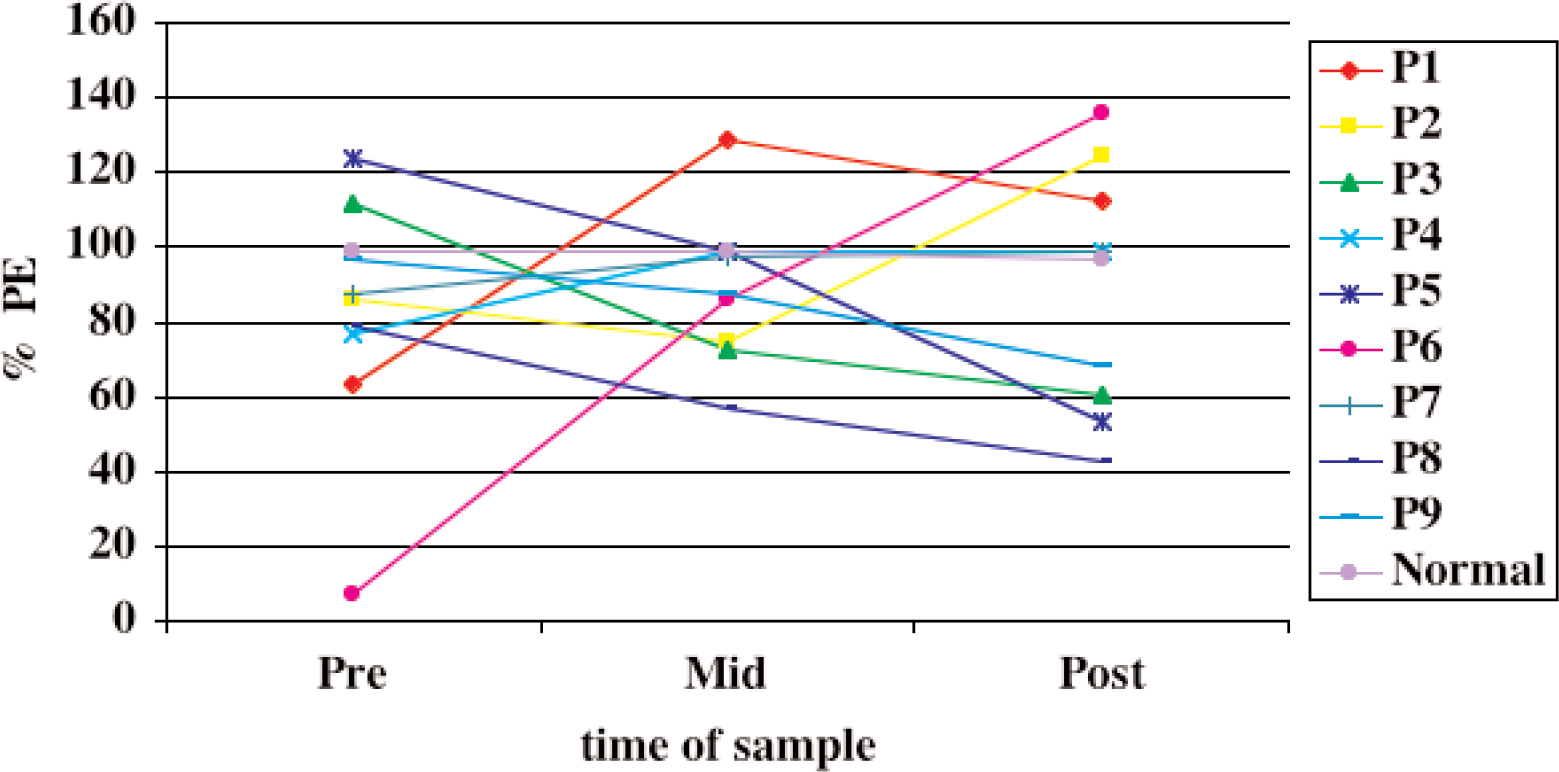
Toxicity of serum obtained from radiotherapy patients before, during and six weeks after treatment. All data were normalized to 100% for the control. The actual mean control-plating efficiency for the three time points was 24.5 + 3.7.
These results, although very preliminary, do give hope that a new biological dosimetric test, which gives useful data concerning low dose response, can be developed.
Most tests for radiation exposure concentrate on measuring chromosome damage in lymphocytes, 12, –15 assuming that if the lymphocytes were circulating at the time of irradiation they would have received a number of double strand breaks proportional in some way to the absorbed dose. If there was no proportionality between dose and double strand breaks this may have been attributable to repair of the breaks, although repair of some lymphocytes may be restricted. Physiology is also relevant here, as the number of lymphocytes circulating at the time irradiation will decrease through natural wastage. In other words, any values obtained are useful only if the time of irradiation is known.
It would appear from our preliminary data that an ‘adaptive model’ corrects the excesses of the initial response and can be thought of as a stabilizing mechanism. In other words, patients that produce an initial large bystander effect produce an adaptive response that mitigates this. In patients with a small bystander effect, the adaptive response produces the opposite effect. The magnitude and direction of any adaptive response is thus dependent on initial conditions and so might well fall into a chaotic model.
Footnotes
ACKNOWLEDGMENTS
We acknowledge St. Luke's Institute of Cancer Research, the Science Foundation of Ireland and the Irish Cancer Research Advancement Board for funding.