
Abstract
A major concern of exposure to low doses of radiation is the risk of cancer induction. Epidemiologic data are rarely powerful enough to accurately discriminate this risk at doses <10 cGy. In order to gain insight into events at these low doses, laboratory-based studies of relevant endpoints are required. One such endpoint is radiation-induced neoplastic transformation in vitro. Such studies can provide quantitative dose-response data, as well as insights into underlying cellular and molecular mechanisms. Data are presented that indicate that low doses of low LET radiation can suppress neoplastic transformation in vitro to levels below that seen spontaneously. Mechanisms involved include both the death of a subpopulation of cells prone to spontaneous neoplastic transformation and the induction of DNA repair. The relative contributions of these mechanisms is dose-dependent. The relevance of these observations to radiation risk estimation is discussed.
I. INTRODUCTION
The study of the effects of low doses of radiation has long been recognized to be of great importance to the understanding of potential risk associated with such exposures. The gold standard with respect to estimation of such risks continues to be epidemiologic studies of human populations that have been exposed to radiation of one type or another. 1,2 Current regulations for radiation exposure are based on the fact that the majority of epidemiologic data conform, within statistical confidence limits, to a linear, no-threshold hypothesis. While epidemiologic studies are clearly of value to the estimation of risk following relatively high dose radiation exposure (e.g. >10 to 20 cGy), they are rarely sensitive enough to accurately estimate risks at low doses (e.g. <10 cGy). Thus, laboratory-based studies of low dose effects using relevant endpoints are required. Until very recently there has been a paucity of such data, most likely for the practical reason that low dose effects are difficult to measure. However, the advent of the Low Dose Radiation Research Program sponsored by the U.S. Department of Energy has provided a stimulus for investigators to attempt such research. Laboratory studies can take two main approaches. The first is to attempt to gain insight into possible molecular and cellular events following such low doses that in turn may help predict possible carcinogenic effects. The second is to actually attempt to perform quantitative dose-response studies in the low dose (<10 cGy) region. We have attempted to embrace both approaches using the endpoint of neoplastic transformation in vitro.
II. ASSAY OF NEOPLASTIC TRANSFORMATION IN VITRO
Neoplastic transformation in vitro simply means the conversion of cells in culture from a non-tumorigenic phenotype to a tumorigenic phenotype. Such phenotypes are identified by whether the cells will or will not grow tumors following implantation into suitable host animals. Quantitative studies of radiation effects using neoplastic transformation as an endpoint have been performed for the last forty years. These studies have almost exclusively been carried out using various rodent cell lines and at high radiation doses. They have been very useful in the examination of the effects of dose-rate, dose-fractionation, LET, and chemical modifiers where they have shown clear relevance to carcinogenesis in vivo. 3 Ideally, since we are interested in radiation-induced human cancers, principally carcinomas, such studies would be carried out using normal human epithelial cells. Unfortunately, both normal and immortalized human cells have proven to be extremely refractory to neoplastic transformation in vitro. 4 The only quantitative assay that is human cell-based is the HeLa x skin fibroblast human hybrid cells assay developed in our laboratory. 5,6 In our opinion, this assay has several advantages over the more widely used rodent cell-based assays. These are that the cells used are human-derived, that the assay relies on a molecular marker of neoplastic transformation rather than a change in morphology, that the mechanism of transformation is known to involve the loss tumor suppressor genes on chromosomes 11 and 14 7 , and that the assay takes 21 to 24 days to completion compared to around 50 days for the often used mouse cell-based C3H10T1/2 assay. This latter point is of significant practical advantage, particularly for low dose assays that necessarily involve many culture dishes and several repeat experiments. On the other hand, it should be made absolutely clear that these human hybrid cells are not normal cells. They are transformed, i.e. they are immortal and do not exhibit contact-inhibited growth. However, they are non-tumorigenic and as such should be regarded as pre-neoplastic cells. The study of radiation effects on such cells is nonetheless of importance since healthy humans harbor preneoplastic tissue.
III. THE SHAPE OF THE DOSE RESPONSE CURVE FOR NEOPLASTIC TRANSFORMATION FOLLOWING LOW DOSES OF LOW LET RADIATION
Azzam et al., 8 using a clone of C3H10T1/2 cells, demonstrated that low doses of Co-60 gamma radiation could suppress transformation frequencies to levels below that seen spontaneously. This observation stimulated us to see if we could observe the same effect in the HeLa x skin fibroblast human hybrid cell system, which we subsequently did using Cs-137 gamma radiation. 9 It is important to note that both studies involved a post-irradiation holding of cells at relatively high cell density for 24 h prior to plating for the transformation assay. We then embarked on a major endeavor to attempt to define the shape of the dose response curve, encompassing several doses both below and above 10 cGy. 10 The data are shown in Fig. 1 and it can be seen that the dose-response curve shows a tendency at low doses to deviate from the linear extrapolation from high doses. Importantly, a similar phenomenon was subsequently demonstrated for 60 kVp x-rays (see Fig. 2), an energy widely used in diagnostic radiology. 11 This suppression of transformation could be the consequence of an adaptive response, 12 classically thought of as the induction of some sort of protective mechanism, e.g. DNA repair. This induction takes some hours to maximize which could explain why we see the suppressive effect more clearly with delayed plating. An alternative explanation for the suppression could be the selective killing of a subpopulation of cells already destined to become neoplastically transformed. 13,14 Indeed, both of these mechanisms could be operative, although with different dose dependencies, much along the lines of hyper-radiosensitivity and induced-radioresistance described by Joiner and colleagues, 15 and supported by the recent observation that there appears to be a threshold dose for DNA double-strand break repair to come into play. 16 Recent mechanistic studies from our own laboratory would also support this concept. 17 At the very lowest doses used (<0.1 cGy), not all cells experience an ionizing event, and bystander effects 18,19,20 may come into play. In summary, in this author's opinion it is likely that multiple mechanisms are involved and that their relative importance is dose-dependent. 17
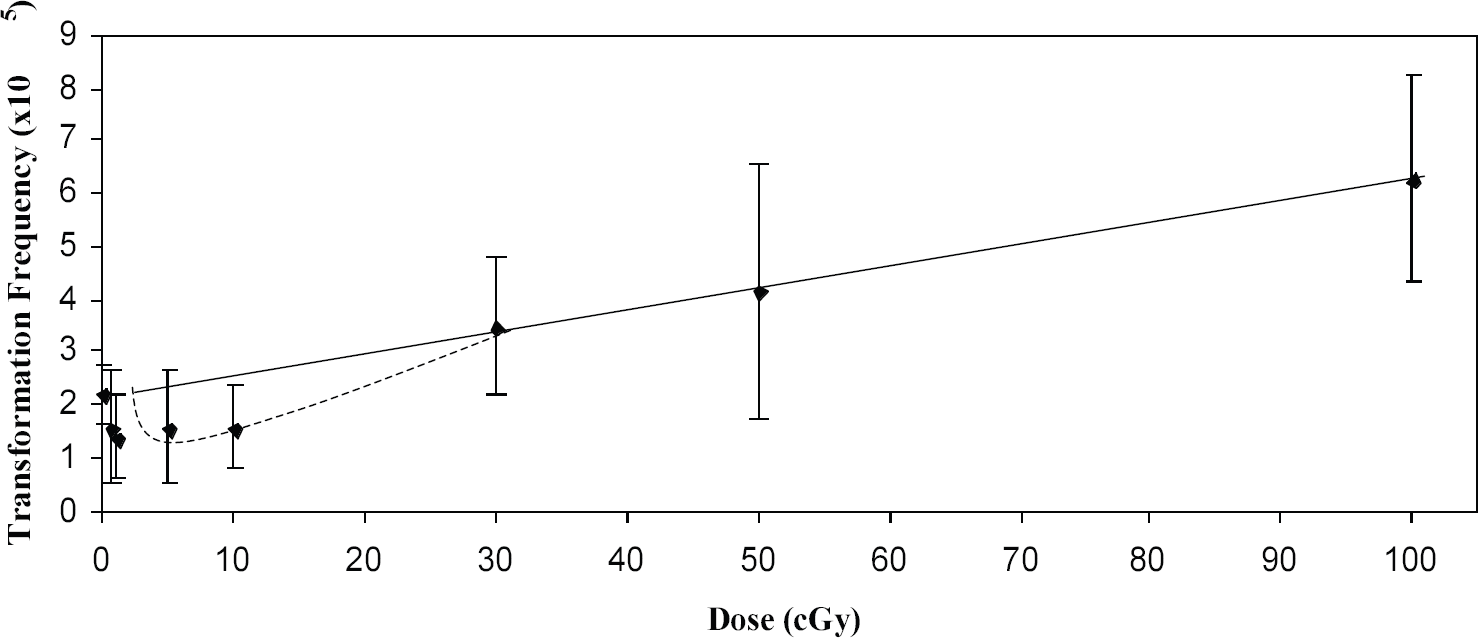
Transformants per surviving cell as a function of dose for HeLa x skin fibroblast human hybrid cells irradiated with Cs-137 gamma radiation. Error bars represent 95% confidence intervals. The straight line represents a linear regression through the high dose points (30, 50 and 100 cGy) and the zero dose point. Data are for 24h post-irradiation delayed plating. Reproduced from Ref. 10 with permission of the Radiation Research Society.
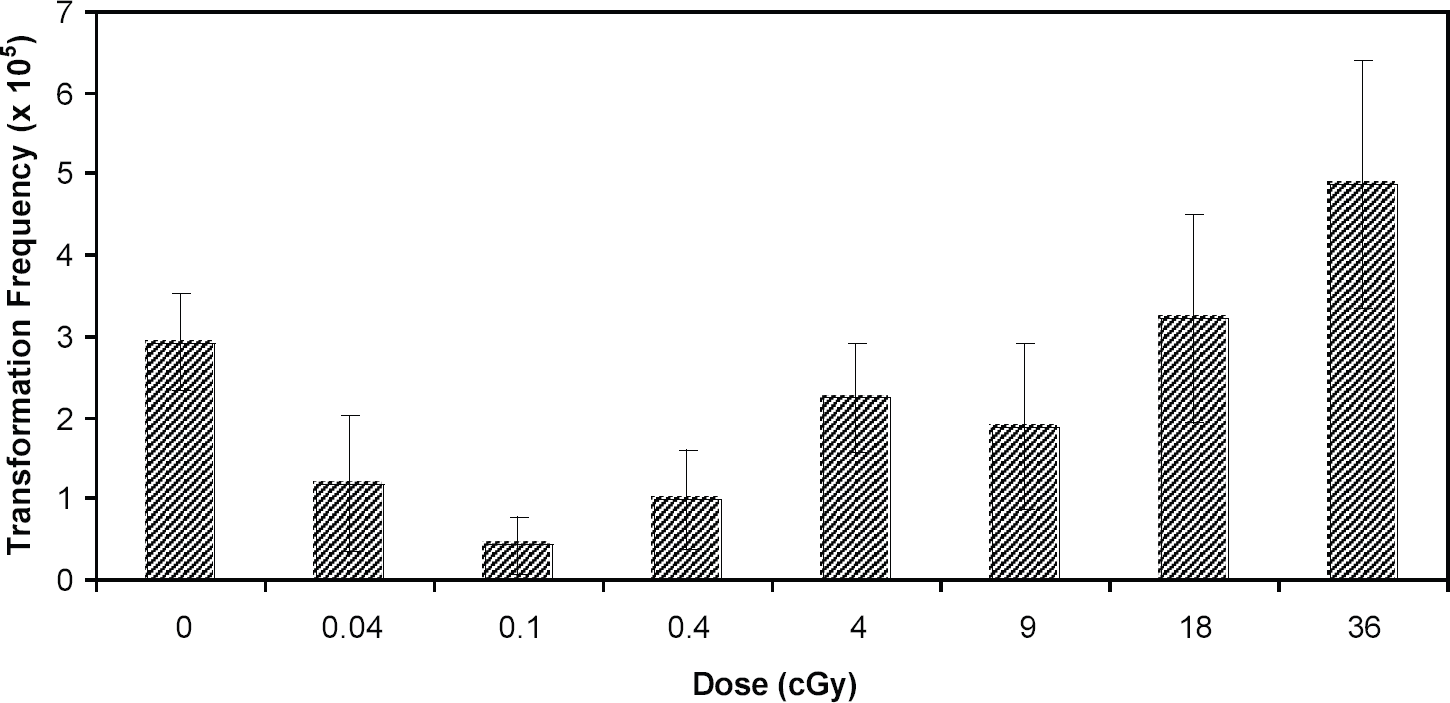
Dose-dependence for neoplastic transformation of HeLa x skin fibroblast human hybrid cells by 60 kVp X-rays. Data are for 24h post-irradiation delayed plating. Error bars represent 95% confidence intervals. Reproduced from Ref. 11 with permission of Taylor and Francis Ltd.
IV. RELEVANCE TO CANCER RISK IN HUMANS
The radiation-induction of cancer in humans is dependent on many factors, both genetic and epigenetic, that are not possible to duplicate in cells in vitro. A priori, therefore, one would not expect that studies of pre-neoplastic cells in vitro to have a direct quantitative link to risk estimates in human populations. It was, therefore, surprising to find that relative risk estimates from our in vitro data agree surprisingly well with those for the induction of breast cancer and leukemia in exposed humans at high doses, say >10 to 20 cSv (see Ref. 10 and Fig. 3). Whether this is serendipitous or not, it nevertheless begs the question as to the relevance of our low dose observations to the human situation. The in vitro data are strongly suggestive of relative risks of <1 at low doses. There is evidence that suggests that this could be the case for leukemia 21 , and in breast cancer it is at the most very close to one. 22, –27 It should be stated that this epidemiologic data could equally well be fit by linear-quadratic functions and, indeed, this is what is done for regulatory purposes. However, e.g. in case of leukemia mortality, a best estimate of a threshold dose was 0.09 Sv, 21 i.e. close to what we determine in vitro. Taken together, these observations indicate that the possibility of risks <1, or at the very least the possibility of a threshold dose, must be taken seriously, at least for these tumors. On the other hand, the latest epidemiologic analysis of all solid cancers in the A-bomb survivors28 shows no evidence for a risk of <1 at the lowest doses (<0.1 Sv), although the upper 95% confidence limit for a threshold is either 0.06 Sv or 0.10 Sv, depending upon whether survivors beyond 3,000 m from the epicenter are omitted or not. It would be desirable if the solid tumor analysis were feasible on an individual tumor basis in order to remove any confounding of the data by combining tumor types with different responses.
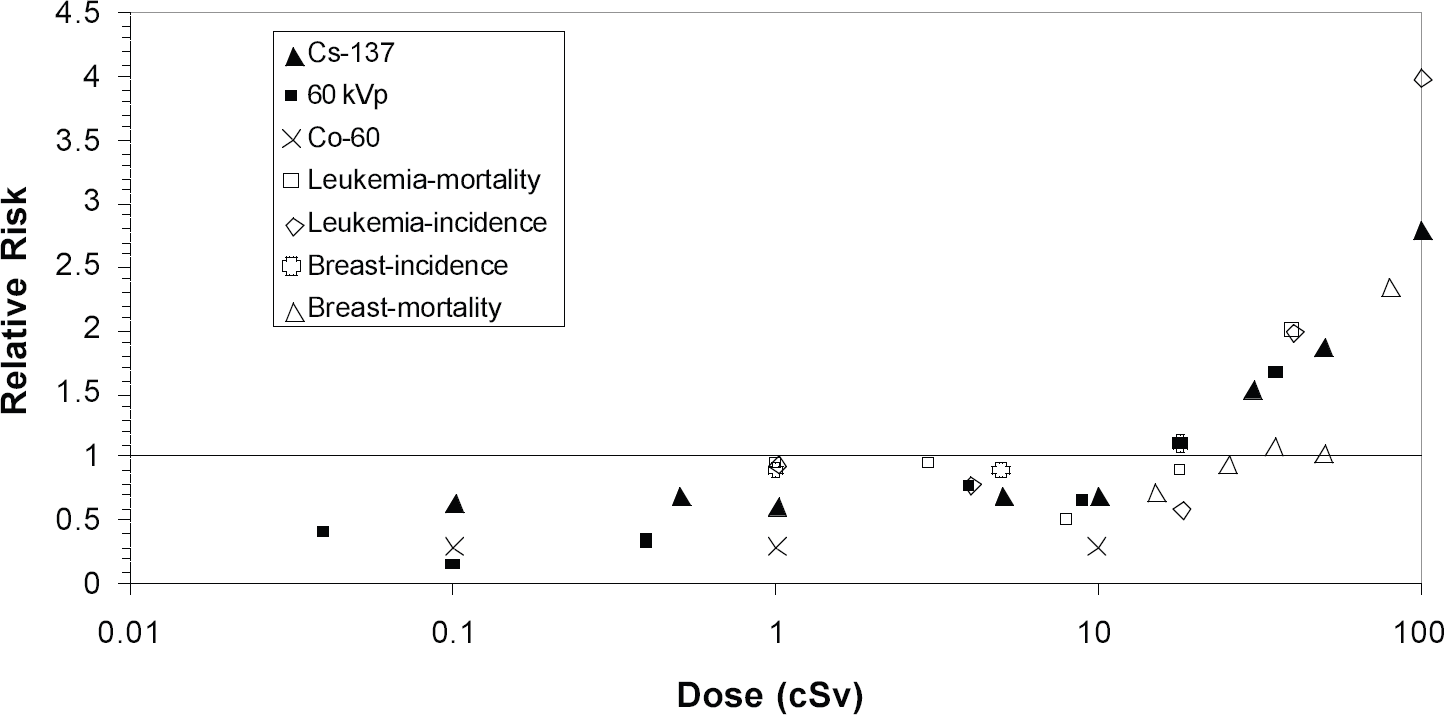
Relative risk as a function of dose for neoplastic cell transformation in vitro and leukemia and breast cancer induction in humans.
It is of interest to speculate that those tumors for which there may be a relative risk of <1, or a true threshold dose, may be the tumors that are more readily inducible at high dose (e.g. breast and leukemia), i.e. where the tissue at risk has more of a genetic predisposition to genomic instability. The rationale for this being that normal tissues that may be more susceptible to carcinogenesis at high doses because they have a relatively low level of endogenous protection, and thus may be the very tissues that are more influenced by an induced increase in this protection at low doses. Further laboratory studies with appropriate cell lines may be able to address this question.
Footnotes
ACKNOWLEDGMENTS
Supported by the DOE Low Dose Radiation Research Program Grant Numbers DE-FG07–99ER62876 and DE-FG03–02ER63309.