
Abstract
Almost all of our knowledge about the mutational effect of radiation has come from high dose studies which are generally not relevant to public exposure. The pKZ1 mouse recombination mutagenesis assay enables study of the mutational effect of very low doses of low LET radiation (μGy to cGy range) in a whole animal model. The mutational end-point studied is chromosomal inversion which is a common mutation in cancer. We have observed 1) a non-linear dose response of induced inversions in pKZ1 mice exposed to a wide dose range of low LET radiation, 2) the ability of low priming doses to cause an adaptive response to subsequent higher test doses and 3) the effect of genetic susceptibility where animals that are heterozygous for the Ataxia Telangiectasia gene
INTRODUCTION
It is known that as the dose of ionising radiation increases then the number of mutations resulting from that exposure increases. The relationship between dose of ionising radiation and DNA damage appears to be linear at high doses. The linear no threshold (LNT) model predicts that this relationship is also linear at low doses and that any dose above zero will result in some DNA damage. Almost all of the data on mutation in response to ionising radiation comes from studies of high doses. However, the population is normally unlikely to be exposed to high doses of radiation. There is little evidence that low doses of radiation (less than 0.1 Gy) are mutagenic and there is some suggestion that low doses of radiation may even be beneficial (Pollycove and Feinendegen 2003). It is important to determine whether there is a threshold dose at which no DNA damage occurs. There is, however, a paucity of
The pKZ1 transgenic recombination mutagenesis model has proven to be a sensitive model for detecting chromosomal inversions in spleen in response to very low doses of three known chemical carcinogens, cyclophosphamide (Sykes et al. 1998), etoposide (Hooker et al. 2002) and mitomycin C (unpublished results). In the case of cyclophosphamide the model has been shown to be 4 orders of magnitude more sensitive at detecting inversions than other mouse models are at detecting point mutations (Kohler et al. 1991; Sykes et al. 1998). The mice contain a transgene which has the
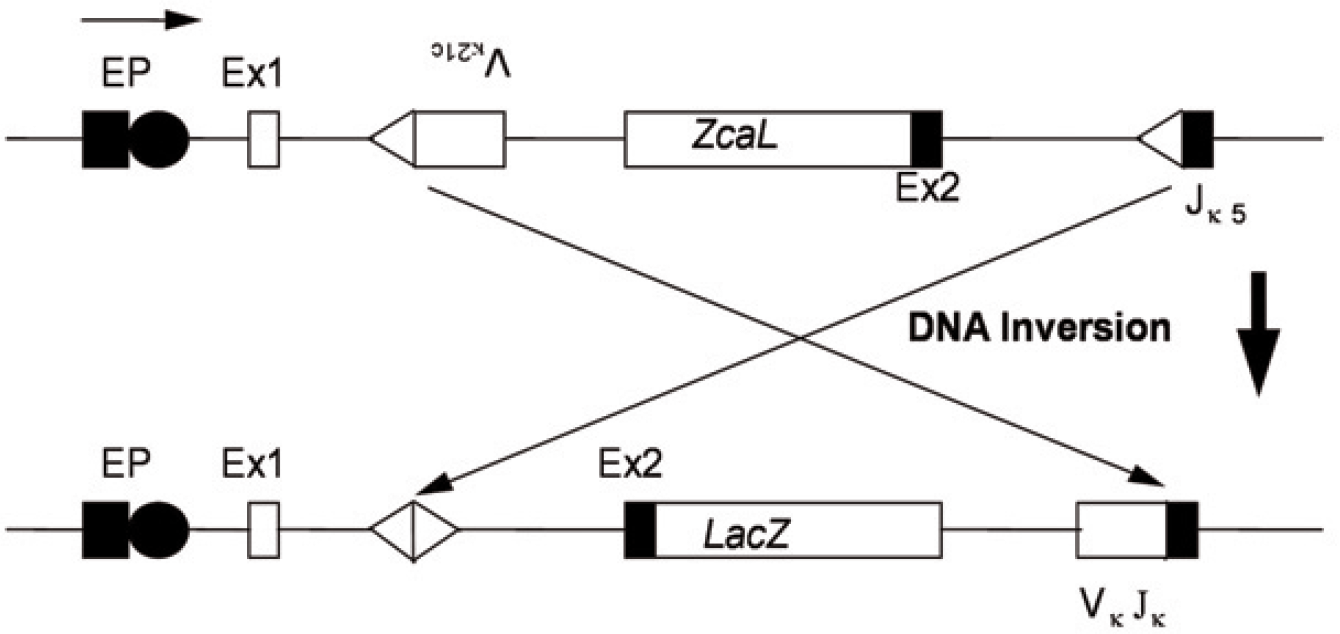
The pKZ1 transgenic mouse construct.
We set out to determine if the pKZ1 recombination mutagenesis model would provide a sensitive mutation assay in response to single whole body doses of low dose, low LET radiation in order to determine if the LNT model applies at doses < 0.1 Gy. We also utilised the pKZ1 model to study adaptive response and genetic susceptibility in response to low dose radiation exposure.
METHODS
Mice
A colony of pKZ1 mice was maintained by breeding pKZ1 mice to C57/BL6J mice. Double transgenic mice were bred by crossing pKZ1 heterozygous mice with
Irradiation of Mice
The mice were irradiated using a Philips deep X-ray unit. The machine was operated at 250 kV and 12 mA, HVL of 3 mm Cu, Source to Surface Distance (SSD) ranged from 80–291 cm and field size was 25 cm x 25 cm. The X-ray machine output was calibrated according to IPEMB protocol (Institution of Physics and Engineering in Medicine and Biology 1996). This dosimetry system has a calibration factor that is traceable to the Australian Radiation Protection and Nuclear Safety Agency.
Irradiation was carried out after placing mice in a perspex mouse holder. Radiation doses less than 0.01 Gy were delivered at a SSD of 291 cm and using additional lead filters to reduce the dose rate. The control mice were also placed in the perspex holder for the same period of time but without the irradiator turned on. After irradiation the mice were returned to their normal housing cages where they were housed for three days.
For adaptive response experiments, mice were exposed to a low priming dose of radiation and then a higher test dose 4 hours later. Control mice included sham-treated mice as well as priming dose only and test dose only.
Histochemical Detection of Inversion Events
The mice were euthanased by CO2 asphyxiation and dissected. The spleen was snap frozen in OCT cryoprotectant embedding compound (Tissue-Tek) and stored at −20°C. Frozen tissue sections were cut on a cryostat and stained for β-galactosidase (β-gal) expression using the method described in Sykes et al (1998). Briefly, 5 μm frozen spleen sections were fixed in glutaraldehyde, stained with X-gal and counter-stained with neutral red.
RESULTS
Threshold
At high doses of radiation (> 0.1 Gy) an increase in chromosomal inversions was observed compared with the endogenous inversion frequency. Between 0.1 mGy and 100 mGy a decrease in inversions below endogenous frequency was observed, and at very low doses (< 0.01 mGy) an increase in inversions was again detected until the frequency returned to the endogenous frequency observed in the sham-treated animals.
Adaptive Response
Mice were treated with 0.01 Gy priming dose and then 1 Gy challenge dose 4 hours later. Control groups included sham-treated, priming alone and challenge alone. No difference was observed in mutation frequency between sham-treated animals and those receiving 0.01 Gy. There was a trend towards an increase in inversion frequency for the 1 Gy treatment groups. The groups that received a priming dose of 0.01 Gy followed by a challenge dose of 1 Gy 4 hours later showed a decrease in inversions to a level below endogenous frequency.
Genetic Susceptibility
pKZ1 mice which have two normal copies of the
DISCUSSION
The results presented here indicate that the pKZ1 model will be a useful whole animal model for studying the mutagenic effect of low dose radiation. We observed an inversion response after exposure to doses of radiation that are orders of magnitude lower than have previously been reported. The dose-response observed was non-linear at doses less than 0.1 Gy. At very low doses and at high doses an increase in inversions was observed, and at intermediate doses a decrease below endogenous inversion frequency was observed. The shape of the dose-response observed is shown in Figure 2. The only report of a whole animal model exhibiting an induction of mutations at X-radiation doses of < 0.1 Gy was by Scheistl

Low dose radiation dose-response curve. Inversions were induced in pKZ1 spleen at very low and at high doses of radiation exposure. Intermediate doses of radiation caused a decrease below endogenous inversion frequency. The straight line represents the LNT theory.
The adaptive response can be defined as a change in sensitivity to radiation damage induced by a low dose, which is delivered prior to a larger challenge dose (Wolff 1998). This has the potential to affect the risk estimation of radiation exposure. The risk of damage from high dose radiation exposure may be decreased if there has been prior exposure to low dose radiation, affecting the validity of the risk predicted using the LNT theory. Our results strongly suggest that a low priming dose of 0.01 Gy is capable of reducing the frequency of inversions caused by a subsequent challenge dose of 1 Gy. The aim here was to determine if inversions could be used as an end-point to study adaptive response at doses of radiation that are lower than have previously been reported. These experiments suggest that the pKZ1 mouse mutation model is an appropriate whole animal system for measurement of the adaptive radiation response. The priming dose (0.01 Gy) used in these experiments is amongst the lowest priming dose reported to be capable of inducing an adaptive response in either
Different individuals in the population will have different genetic susceptibilities to DNA damaging agents such as ionising radiation based on their ability to repair damaged DNA. Individuals homozygous for
DNA double strand breaks are known to be the major lesion induced after X-irradiation. The pKZ1 mutagenesis model allows measurement of inversion events, which arise through the repair of DNA double strand breaks. In spleen tissue in the pKZ1 model, inversion events may be recognized and repaired using the RAG1/RAG2 mediated NHEJ machinery which is important in both repair of DNA double strand breaks and in the normal development of B- and T-cell receptors (reviewed in Lieber 1998). Investigation of the important enzymes in the NHEJ pathway (for example RAG-1, RAG-2, DNA-PK, Ku 70, Ku 86), will hopefully lead to further clues about the underlying mechanism of changes in inversions identified in the pKZ1 mouse spleen in response to low dose radiation.
The results presented here indicate that the pKZ1 mouse model has the sensitivity to enable study of low dose radiation exposure which is relevant to human population exposure. The model can also be used to study adaptive response and genetic susceptibility in response to low dose radiation exposure. Our data using DNA inversions as a mutagenic end-point indicate that the dose response to X-radiation is not linear and that the validity of the LNT model needs to be revisited. This will have an important impact when setting exposure limits and conditions for safe occupational and medical radiation exposure.
Footnotes
ACKNOWLEDGMENTS
We wish to thank Professor Martin Lavin (Queensland Institute of Medical Research) for kindly providing heterozygous