
Abstract
Intense stress can challenge tissue homeostasis and accelerate the ageing process. However, several lines of evidence indicate that repeated mild stresses can have beneficial and even life-prolonging effects. Hypersecretion of glucocorticoids (GC) represents the major hormonal response to stress. Besides its life-sustaining role, GC excess, usually due to several side-effects that promote a “catabolic” phenotype, can be detrimental for several tissues. Cushing's syndrome patients are characterized by chronic endogenous GC excess and consequently at the time of diagnosis they have an atrophic elderly-like skin. Interestingly, when Cushing's syndrome fibroblasts were removed from the high-GC milieu in vivo and cultured in vitro under standard conditions they express an “anabolic” phenotype, i.e. they restore their ability for collagen synthesis, they secrete reduced levels of metalloproteases (MMP-1 and MMP-2) and have an increased proliferative capacity and contractility. Furthermore, these cells exhibit a significant extension of their proliferative lifespan, while they respond better to exogenous stress by producing significantly higher levels of heat-shock protein-70 (HSP70). These results imply that long-term hypercortisolism in vivo can have beneficial consequences on fibroblast physiology in vitro.
INTRODUCTION
Ageing represents a failure in tissue homeostasis, in both maintenance and repair processes. At the cellular level, it is now well known that normal cells have only a limited lifespan, i.e. when cultured in vitro they can undergo only a limited number of population doublings before they stop proliferating and enter a so-called “senescent” state (Smith and Lincoln 1984; Kletsas 2003). Senescent fibroblasts exhibit an altered phenotype, i.e. they are enlarged, flattened, irregular in shape, with large lobulated nuclei, and increased number of vacuoles and lysosomal granules (Kletsas 2003). Furthermore, they display major alterations in functional characteristics, beyond their inability to proliferate. DNA microarray analysis has revealed that senescent fibroblasts exhibit a “pro-inflammatory” phenotype, e.g. they suppress collagen synthesis and they express increased levels of metalloproteases, inflammatory cytokines or other pro-inflammatory molecules, such as ICAM-1 (Shelton et al. 1999; Gorgoulis et al. 2005). Due to this phenotype, it is believed that the accumulation of senescent cells in the tissues during ageing contributes to the ageing process and the development of age-related diseases (Hornsby 2002). Especially, in the connective tissue such as in skin, ageing is leading to the thinning of the dermis (Lavker et al. 1989) and in a decreased ability for wound healing (Gerstein et al. 1993).
It is generally believed that chronic or intense exogenous stresses are able to accelerate the ageing process. At the cellular level, extreme – still non-lethal – stresses, such as treatment with H2O2, tert-butylhydroperoxide, ethanol or UV light, can lead to a stress-induced premature senescence (SIPS) (Toussaint et al. 2002). The stress-induced senescent cells are in many aspects similar, although not identical, to the cells that became senescent by successive replications (replicative senescence). On the other hand, repeated treatments with mild stresses, such as heat or cold shock, mild irradiation, hypergravity, exercise etc. has been reported to generate anti-ageing or even life-prolonging effects at the cellular or organismal level (Minois 2000; Rattan 2001), representing another example of “hormesis” (Calabrese and Baldwin 2000). However, the possibility that stress hormones-that regulate the stress response at the level of whole organism-can have such a dual role has not been studied so far.
Cortisol hypersecretion, as a result of the activation of the hypothalamic-pituitary-adrenal (HPA) axis, is one of the primary hormonal responses to stress. Although a short-term cortisol excess elicits a life-sustaining role, chronic exposure to glucocorticoids (GC) is responsible for a variety of adverse effects in several cell types (Munck et al. 1984). Indeed, chronic GC excess, either exogenous, as a part of treatment regimes, or endogenous, as a result of chronic stress, has severe detrimental effects in almost all tissues (Boumpas et al. 1993; Boscaro et al. 2001). In addition, it has been suggested that GCs may accelerate the ageing process (Sapolsky 1996). The skin is one of the tissues most profoundly affected by glucocorticoid excess, leading to atrophy and impaired wound healing (Sowers and Lippman 1985; Oikarinen and Autio 1991; Boscaro et al. 2001), a state reminiscent of the aged skin. These detrimental effects of GCs in skin are mediated primarily by interferences with skin fibroblasts' function. In particular, in vitro studies have shown that GCs induce profound alterations on several parameters of extracellular matrix (ECM) homeostasis, such as a decrease in collagen synthesis and an altered expression of matrix metalloproteinases (MMPs), leading finally to a reduction of the collagen mass (Autio et al. 1994; Slavin et al. 1994; Oikarinen et al. 1998). In general, increased (pharmacological) doses of GCs induce a “catabolic” phenotype in skin fibroblasts in vivo and in vitro.
Endogenous Cushing's syndrome is a rare disease due to cortisol overproduction by the adrenal glands (Orth 1995). Most patients have atrophic skin and impaired wound healing. Skin fibroblasts from these individuals therefore represent a model suitable for the investigation in vitro of the cumulative effects of prior exposure to elevated (non-pharmacological) glucocorticoid levels in the absence of an exogenous stress. In an effort to understand if this exposure may affect – negatively or positively – several aspects of tissue homeostasis, primary cultures of skin fibroblasts obtained from patients suffering from endogenous Cushing's syndrome (Cushing Fibroblasts: CF) have been developed and their functional characteristics when grown under standard culture conditions in vitro in the absence of exogenous GC administration were studied. In particular, possible long-lasting alterations concerning central fibroblast activities involved in skin homeostasis and wound repair, such as proliferation, contraction, production and degradation of extracellular matrix components, as well as cellular longevity, were investigated – in comparison to skin fibroblasts from normal donors (Normal Fibroblasts: NF).
COLLAGEN SYNTHESIS AND CATABOLISM IN CUSHING'S SYNDROME FIBROBLASTS
Thinning of the dermis is one major feature of the aged skin (Lavker et al. 1989). Collagen is the most abundant protein in the dermis and consequently inhibition of its synthesis is a primary cause of dermal atrophy. Furthermore, an increase in matrix metalloproteases (MMPs) secretion, enhancing collagenolysis, could play equally important role in the deregulation of skin homeostasis. Indeed, a reduction in collagen synthesis and an increase in the expression of matrix metalloproteases, has been found during senescence (Khorramizadeh et al. 1999). Also treatment with glucocorticoids has a “catabolic” effect in skin, similar to age-related alterations. It is well known that exposure to glucocorticoids grossly diminishes type I procollagen synthesis at the mRNA and protein level in human dermal fibroblasts (Cockayne et al. 1986; Slavin et al. 1994). This reduction is due to the coordinate action of GCs through both glucocorticoid-and TGF-β- responsive elements (Meisler et al. 1995). When skin fibroblasts from Cushing's syndrome patients were removed from the high cortisol milieu in vivo and cultured in vitro in the absence of cortisol they expressed comparable amounts of α1-procollagen mRNA with fibroblasts from normal donors (authors' personal unpublished data). Furthermore, collagen synthesis, measured by the incorporation of tritiated proline in newly secreted collagen was restored to normal levels (Table 1). In addition, when treated with TGF-β - a potent inducer of collagen synthesis - Cushing's and normal skin fibroblasts exhibited similar stimulation of 3H-proline incorporation over control levels (Zervolea et al. 2005). Even more, these cells were found to secrete significantly lower levels of the metalloproteases MMP-1 and MMP-2, compared to normal fibroblasts (Table 1). Regarding the tissue inhibitors of MMPs (TIMPs), CF were found to secrete lower levels of TIMP-1 than NF, while there was no significant difference in TIMP-2 levels (Table 1).
Extracellular Matrix Production & Catabolism by Cushing Fibroblasts
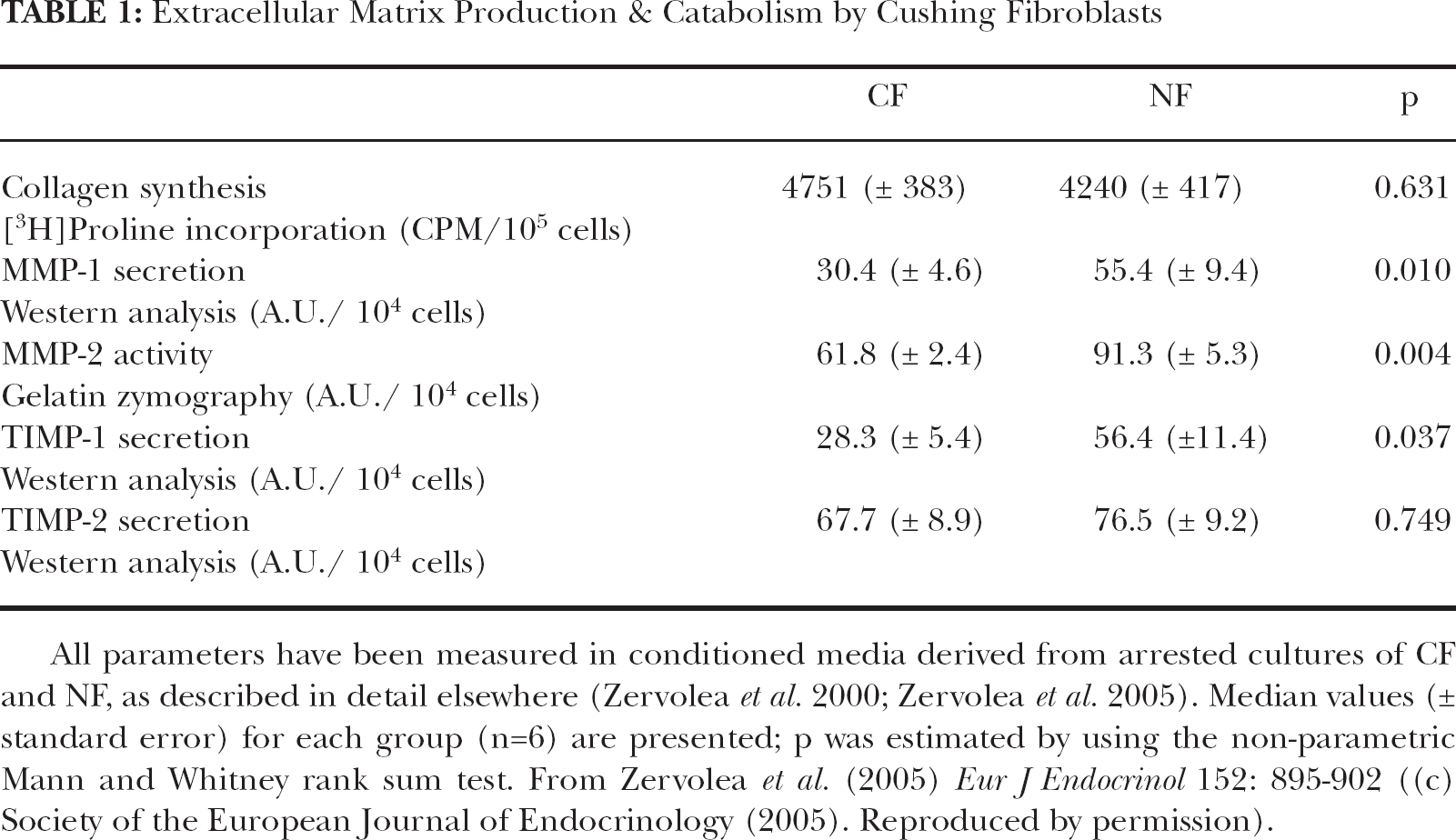
All parameters have been measured in conditioned media derived from arrested cultures of CF and NF, as described in detail elsewhere (Zervolea et al. 2000; Zervolea et al. 2005). Median values (± standard error) for each group (n=6) are presented; p was estimated by using the non-parametric Mann and Whitney rank sum test. From Zervolea et al. (2005) Eur J Endocrinol 152: 895–902 ((c) Society of the European Journal of Endocrinology (2005). Reproduced by permission).
CUSHING'S SYNDROME FIBROBLASTS EXHIBIT AN INCREASED PROLIFERATIVE CAPACITY AND CONTRACTILITY IN VITRO
It was further discovered that Cushing's syndrome patients' skin fibroblasts can proliferate much faster, compared to cells from control normal donors. This has been shown both by their increased ability to synthesize DNA, a prerequisite for proliferation, and their shorter population doubling time (Table 2). In addition, Cushing's fibroblasts were found to achieve much higher final cell densities than fibroblasts from normal donors (Table 2).
Proliferative & Contractile Characteristics of Cushing Fibroblasts

Proliferative characteristics of CF and NF cultured in the presence of 10% Fetal Bovine Serum, and cell-populated collagen gel contraction were studied as described in detail elsewhere (Zervolea et al. 2005). Median values (± standard error) for each group (n=6) are presented; p was estimated by using the non-parametric Mann and Whitney rank sum test. From Zervolea et al. (2005) Eur J Endocrinol 152: 895–902 ((c) Society of the European Journal of Endocrinology (2005). Reproduced by permission).
Contraction represents an important fibroblast function in normal skin homeostasis, as well as in the wound healing process. Gels of polymerized collagen populated by fibroblasts provide a functional in vitro model of tissue contraction (Grinnell 2000). Glucocorticoids, such as dexamethasone or hydrocortisone, can reduce skin fibroblast-mediated contraction, and this effect may play a role in the delay in the wound-healing process (Gabbiani 1979; Coulomb et al. 1984). This inhibition is reversed when Cushing's fibroblasts are transferred in vitro and cultured in the absence of glucocorticoids: collagen matrices populated by an equal number of Cushing's and normal fibroblasts were more intensely contracted by the former compared to the latter (Table 2).
All the data presented above indicate that when skin fibroblast from Cushing's syndrome patients where removed from the high-cortisol milieu and cultured in vitro under standard conditions, they restore their ability for collagen production, they secrete lower levels of metalloproteases, and they exhibit an increased proliferative capacity, a higher final cell culture density and an increased contractility. Collectively, these anabolic responses are in clear contrast to the catabolic effects of glucocorticoids observed in vivo or in vitro and, indicate that prior chronic exposure to glucocorticoid excess is not associated with a persisting adverse outcome in the functional phenotype of these cells.
FIBROBLASTS FROM CUSHING'S SYNDROME PATIENTS EXHIBIT AN INCREASED LIFESPAN IN VITRO
As mentioned above, Cushing's syndrome patients have an elderly-like skin. Given that skin fibroblasts derived from patients suffering from age-related pathologies, e.g. diabetes, exhibit a shorter lifespan in vitro (Cristofalo et al. 1998), the in vitro longevity of Cushing's syndrome fibroblasts was measured. Unexpectedly, these cells exhibit a near 40% extension of their lifespan, compared to cells from sex- and age-matched control donors (Figure 1).

In vitro proliferative life-span of Cushing's syndrome patients' fibroblasts. CF and NF were cultured until they reached replicative senescence. The values represent the cumulative population doublings achieved by each strain. The horizontal bars indicate the median value for each group; p was estimated by using the non-parametric Mann and Whitney rank sum test. Reproduced from Pratsinis et al. (2002) Exp Gerontol 37: 1237–1245, with permission from Elsevier.
The effect of increased cortisol concentrations on the longevity of human fibroblasts has been already addressed, though with controversial results. Early studies have shown a significant extension of the lifespan of embryonic lung fibroblasts when cultured in the presence of cortisol (Maciera-Coelho 1966; Grove and Cristofalo 1977; Nichols et al. 1977), however, this has not been reproduced by other investigators, using a different protocol (Didinsky and Rheinwald 1981). The discrepancy between the effect of increased cortisol concentrations in vivo and in vitro can be due to the differences between the conditions under which the cells are exposed to cortisol. For example, while in vitro the cells are proliferating in the presence of serum and are exposed in a single dose of cortisol, in the tissue in vivo they are quiescent in the absence of serum, and they are exposed to cortisol in a definite diurnal rhythm, as well as in other local or systemic factors. Indeed, other systemic factors-beyond cortisol-that are also altered in Cushing's syndrome could also be responsible for this phenomenon, such as growth hormone (GH) and Insulin-like Growth Factor-I (IGF-I), which are drastically down-regulated in the plasma of these patients (Wajchenberg et al. 1996; Unterman and Phillips 1985). In this vein, it has been reported that GH and IGF-I overexpression in transgenic mice leads to accelerated ageing (Steger et al. 1993; Gems and Partidge 2001).
Another growth factor possibly implicated in the increased longevity of CF is transforming growth factor-beta (TGF-β), a crucial regulator of cellular homeostasis, which has been recently shown to be also involved in the regulation of cellular lifespan. In particular, it has been shown that stress-induced premature senescence is mediated via TGF-β (Frippiat et al. 2001). The levels of TGF-β secreted by Cushing's and normal fibroblasts were measured, by using a highly sensitive and specific bioassay. In particular, conditioned media (CM) from these cell strains were collected, and the amount of total and active TGF-β secreted was estimated. In agreement with the extended lifespan of CF, these cells secrete lower levels of active TGF-β in their milieu (Pratsinis et al. 2002), and – maybe, more importantly – the average active TGF-β as a percentage of the total is significantly lower in CF than in NF (Table 3). In addition, as reactive oxygen species (ROS) have been inversely related with cellular longevity and ageing in general (Finkel and Holbrook 2000), the internal ROS content in both CF and NF was estimated, after incubation of the cells with the fluorescent substrate DCFH-DA. Although no significant difference between the two groups was found (Table 3), there was a trend for a lower concentration of ROS in CF compared to NF, as expected considering the increased longevity of CF.
Parameters of Stress Resistance and Longevity in Cushing Fibroblasts

TGF-β1 secretion, and HSP70 inducibility of CF and NF cultures were studied as described in detail elsewhere (Pratsinis et al. 2002). Reactive oxygen species was estimated as described in Athanasas et al. (2004). Median values (± standard error) for each group (n=6) are presented; p was estimated by using the non-parametric Mann and Whitney rank sum test. Adapted from Pratsinis et al. (2002) Exp Gerontol 37: 1237–1245, with permission from Elsevier.
RESPONSE OF FIBROBLASTS FROM CUSHING'S SYNDROME PATIENTS TO EXOGENOUS STRESS
It has been long suggested that longevity is inversely linked to stress resistance. In particular, the expression of Heat Shock Proteins (HSP), a group of molecular chaperones, is reduced during in vivo and in vitro ageing (Gutsmann-Conrad et al. 1998; Luce and Cristofalo 1992). It was discovered that HSP70 basal levels (in the absence of any stress application) of CF are relatively lower than that of NF (Pratsinis et al. 2002). After induction with sodium arsenite, a known stressor, the levels of HSP70 levels are dramatically increased in both cell types. However, while the percentage of activation for NF was 4.5-fold (over basal levels), in Cushing's syndrome-patients' fibroblasts an approx. double activation, i.e. 10.7-fold (Table 3) was observed. This highly enhanced response of CF is in agreement with their extended lifespan in culture.
CONCLUSIONS
Intense stresses are known to be detrimental for tissue homeostasis and furthermore they may also accelerate the ageing process. At the cellular level such a stress can provoke modifications in its structure and function, e.g. in its redox status, protein metabolism and stability, DNA synthesis, etc. (Rattan et al. 2004). Moreover, these stresses affect cellular longevity. Probably, the best example in this direction is the application of an exogenous oxidative stress. When early-passage cells are exposed to sublethal concentrations of H2O2 or tert-butylhydroperoxide they enter a permanent arrested state. This phenomenon is called “stress-induced premature senescence” (SIPS) (Toussaint et al. 2000), as these cells resemble those that have undergone senescence after serial subculturing in vitro. In addition, SIPS is provoked when cells are exposed in hyperoxic conditions, i.e. a 40% oxygen partial pressure, compared to the standard condition of 20%. Finally, when cells are cultured under hypoxic conditions (3% oxygen tension) or in the presence of free radical scavengers a considerable extension in their lifespan is observed (Toussaint et al. 2002). In contrast to the above, it has been shown that exposure to mild stress can counteract the negative effects of ageing (Minois 2000). Several types of stress can provoke this anti-ageing effect, e.g. calorie restriction, temperature shock, hypergravity, exercise or irradiation. In a series of studies, Rattan and collaborators have shown that repeated mild heat stress (exposure to 410C for 1h twice a week) has anti-ageing effects. In particular, this treatment reduces the accumulation of oxidatively and glycoxidatively damaged proteins, stimulates the proteasomal activities for the degradation of abnormal proteins, enhances the cellular antioxidant ability and its resistance to other stresses (Rattan et al. 2004). Calorie restriction represents another type of mild stress. When rodents were fed with a balanced diet containing approx. 30% less calories, they were found to live significantly longer that the control – ad libitum fed-mice (Masoro 2000). A similar lifespan extension was found also in fibroblasts derived from these animals (Pendergrass et al. 1995). All these examples collectively indicate that repeated mild stress can have beneficial and even life promoting effect, a phenomenon called “hormesis” (Calabrese and Baldwin 2000).
At the organismal level, systemic responses to stress are of primary importance, as they affect nearly all tissues. In this context the activation of the hypothalamic-pituitary-adrenal (HPA) axis, one of the immediate hormonal responses to stress, is crucial for the maintenance of tissue homeostasis. Cortisol hypersecretion, as a result of this activation, has clearly a life-sustaining role. However, due to several side-effects, chronic exposure to pharmacological doses of glucocorticoids (GC) can be detrimental for almost all tissues and even lead to accelerated ageing (Boumpas et al. 1993; Sapolsky 1996; Boscaro et al. 2001). Cushing's syndrome, a rare disease due to cortisol overproduction by the adrenal glands, represents a good model for studying the cumulative effects of GCs, since patients with this syndrome are exposed in excessive albeit non-pharmacological doses of cortisol for a long period of time, in the absence of an exogenous stress, and consequently they have an elderly-like skin (atrophic and with impaired wound healing) due to the catabolic effects of cortisol. Interestingly, Cushing's syndrome fibroblasts, when removed from the high-GC milieu of in vivo and cultured in vitro under standard conditions, were found to restore their ability for collagen synthesis and to secrete reduced levels of metalloproteases (MMP-1 and MMP-2). Even more, these cells are more vivid in culture than those derived from normal, control donors, as they proliferate faster, they reach higher final culture densities and they have increased contractile ability, all important parameters for the maintenance of tissue homeostasis and a proper wound healing process. Although it has been reported that glucocorticoids can accelerate the ageing process, the data from the in vitro phenotype of CF are pointing to an opposite direction, having in mind that senescent cells lose their ability for proliferation, collagen synthesis and contraction, while they secrete increased amounts of metalloproteases. In agreement, it has been found that Cushing's fibroblasts have a remarkable extension of their lifespan in vitro. Furthermore, it has been shown that CF produce less reactive oxygen species and secrete less TGF-β, two factors that have been inversely related to senescence. More importantly, they are able to respond much better to exogenous stress, by producing nearly double concentrations of inducible HSP70.
As mentioned above, one of the best examples of an extension of lifespan as a result of a long-lasting mild stress is the case of calorie-restricted rodents. Reduction of food intake markedly increases their longevity, retards age-associated physiological decline and delays age-associated diseases (Masoro 2000). Furthermore, calorie restriction leads to increased replicative lifespan, cell proliferation and improved wound repair in vivo and in vitro (Pendegrass et al. 1995; Wolf et al. 1995; Reed et al. 1996). Interestingly, in an analogy to Cushing's syndrome these animals exhibit mildly elevated plasma glucocorticoid levels (Sabatino et al. 1991). Another well known paradigm of hormesis is exercise. This activity although stressful and deleterious in biochemical terms – due to the production of free radicals and stress hormones (Rattan 2005) – has clearly beneficial effects. Interestingly, plasma cortisol was found to be higher in athletes, in agreement with their lower anabolic/ catabolic balance (Grasso et al. 1997).
In conclusion, the data from the in vitro behaviour of Cushing's syndrome fibroblasts indicate that a long-term exposure to high cortisol concentrations in vivo is not associated with persisting adverse changes in skin fibroblasts and/or acceleration of the aging process of these cells. Instead, an unexpectedly positive effect on the physiology of these cells when cultured in vitro has been found, as well as an extension of their lifespan. Further work is needed to explain these anti-ageing effects of the exposure to elevated cortisol concentrations. One possible explanation for this phenomenon can be grounded on the ability of cortisol to control metabolism. It has been hypothesized that optimal allocation of metabolic resources between the maintenance and repair and other functions is crucial for a proper somatic maintenance and for maximizing longevity (Kirkwood and Franceschi 1992). Accordingly, the catabolic action of cortisol leads to severe alterations in tissue homeostasis in Cushing's syndrome patients, such as dermal atrophy and impaired wound repair. However, when these cells are removed from the high-cortisol milieu they express in vitro an “anabolic” phenotype (characterised by increased proliferation, collagen accumulation or contraction) and an extension of their lifespan. This “hormetic” reaction can provide a basis for understanding the beneficial effect of several stresses at the organismal level.