
Abstract
We evaluated the IGF1 system in cholangiocytes of primay biliary cirrhosis (PBC) patients and investigated the relationships with apoptosis. Biopsies of PBC patients (n=32) and normal subjects (n=5) were investigated by immunohistochemistry for expression in cholangiocytes of IGF1, IGF1-R, pAKT, terminal deoxynucleotide transferase end labeling (TUNEL), Bax (proapoptotic protein), and Bcl2 (antiapoptotic protein). Whereas normal cholangiocytes were almost negative, cholangiocytes of PBC patients showed strong IHC staining for IGF1, IGF1-R, and pAKT, which increases from stage I to stage IV, where >70% of cholangiocytes were positive. Bax/Bcl2 ratio reached the highest value (4.6) in PBC stage III when apoptosis is maximal (24% TUNEL positivity), whereas it declines in stage IV (1.4) when only 7.8% cholangiocytes were TUNEL positive. In PBC stages III and IV, expression of IGF1, IGF1-R, and pAKT in cholangiocytes was directly correlated with the antiapoptotic Bcl2 and inversely correlated with proapoptotic Bax, Bax/Bcl2 ratio, and TUNEL positivity. In conclusion, cholangiocytes of PBC patients showed a marked increase in IGF1, IGF1-R, and pAKT expression involving most cholangiocytes surviving in the terminal ductopenic stage. This was associated and correlated with a balance of pro- and antiapoptotic proteins favoring survival rather than apoptosis, suggesting a major role of IGF1 system in promoting cholangiocyte survival.
P
The aim of this study was to evaluate IGF1, IGF1-R, and PI3K/AKPT pathway in cholangiocytes of patients with different stages of PBC and to correlate their expression with markers of apoptosis.
Materials and Methods
Liver Samples and Patient Characteristics
The study was carried out on liver biopsies from 32 post-menopausal female patients with diagnosis of PBC based on standard, internationally accepted criteria (Boyer 1997). Four of eight biopsies of PBC stage IV were obtained from the explanted liver. As controls, we investigated five liver biopsies with a normal histology (from patients submitted to laparotomy). PBC specimens were staged according to Ludwig et al. (1978). All PBC patients and controls were negative for hepatitis B and C markers. All PBC patients were under treatment with ursodeoxycholic acid. The study protocol was in accordance with the ethical guidelines of the 1975 Declaration of Helsinki.
Liver Histology and Immunohistochemistry
Common liver histology was performed in 3- to 4-μm sections stained with hematoxylin-eosin and Masson's stains. For immunohistochemistry (IHC), endogenous peroxidase activity was blocked by a 30-min incubation in methanolic hydrogen peroxide (2.5%). Endogenous biotin was blocked by Biotin Blocking System (Dako; Glostrup, Denmark). Sections were hydrated, rinsed in PBS (pH 7.4), and incubated overnight at 4C with antibodies for cytokeratin 19 (CK-19, 1:100; Dako), proliferating cellular nuclear antigen (PC10, mouse anti-PCNA reacting with all vertebrate species, 1:100; Dako), IGF1 (sc-7144, goat polyclonal, 1:100; Santa Cruz Biotechnology, Santa Cruz, CA), IGF1-R (sc-9038, rabbit polyclonal, 1:80; Santa Cruz Biotechnology); pAKT (sc-16646-R, rabbit polyclonal, 1:100; Santa Cruz Biotechnology), Bax (sc-7480, mouse monoclonal, 1:200; Santa Cruz Biotechnology), Bcl2 (sc-7382, mouse monoclonal, 1:100; Santa Cruz Biotechnology). Samples were rinsed with PBS for 5 min, incubated for 10 min at room temperature with secondary link biotinylated antibody (biotinylated anti-rabbit, anti-mouse, anti-goat, ABC system; DAKO LSAB+), incubated with streptavidin-peroxidase for 10 min, and finally developed with DAB. Negative controls were performed by applying specific non-immune serum. At least six slides were used per patient.
For detection of apoptosis on the single cell we used the terminal deoxynucleotide transferase end labeling (TUNEL) method (ApopTag; Oncor, Gaithersburg, MD).
For each sample, more than five portal spaces limited to interlobular bile ducts (<100 μm in diameter) were studied. Findings were expressed as the percentage of IHC-positive cells over the total number of cholangiocytes by counting >100 nuclei in bile ducts. Histological and IHC determination were performed independently and blindly by two pathologists (PO, AF) from two different institutions, with an agreement always >90%.
Analytical Methods
Liver function tests and hepatitis markers were assessed by routine laboratory methods. Antimitochondrial antibodies status was evaluated by indirect immunofluorescence and by ELISA, as previously described (Alvaro et al. 2004).
Statistical Analyses
Data were expressed as mean ± standard error (SE). Statistics were performed using the Mann-Whitney test and Spearman's correlation coefficient. A p value <0.05 was considered to be statistically significant.
Results
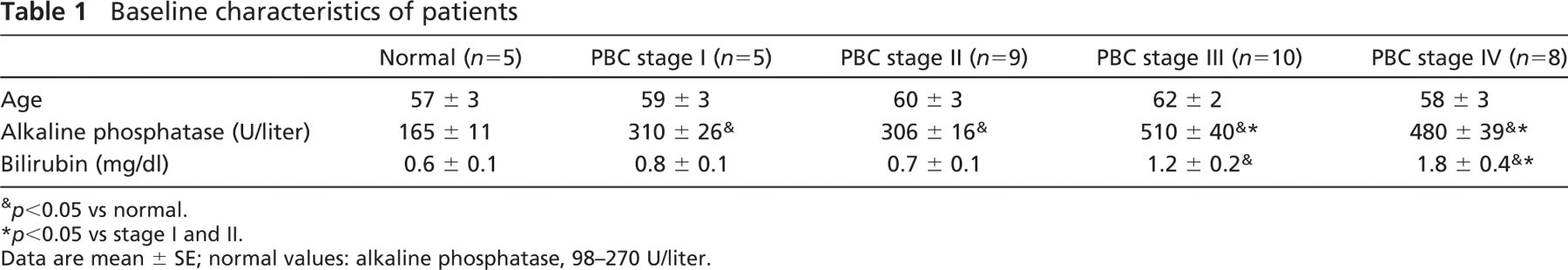
Characteristics of PBC patients and normal controls are reported in Table 1. No significant differences in age exist between patients with different PBC stages and normal controls.
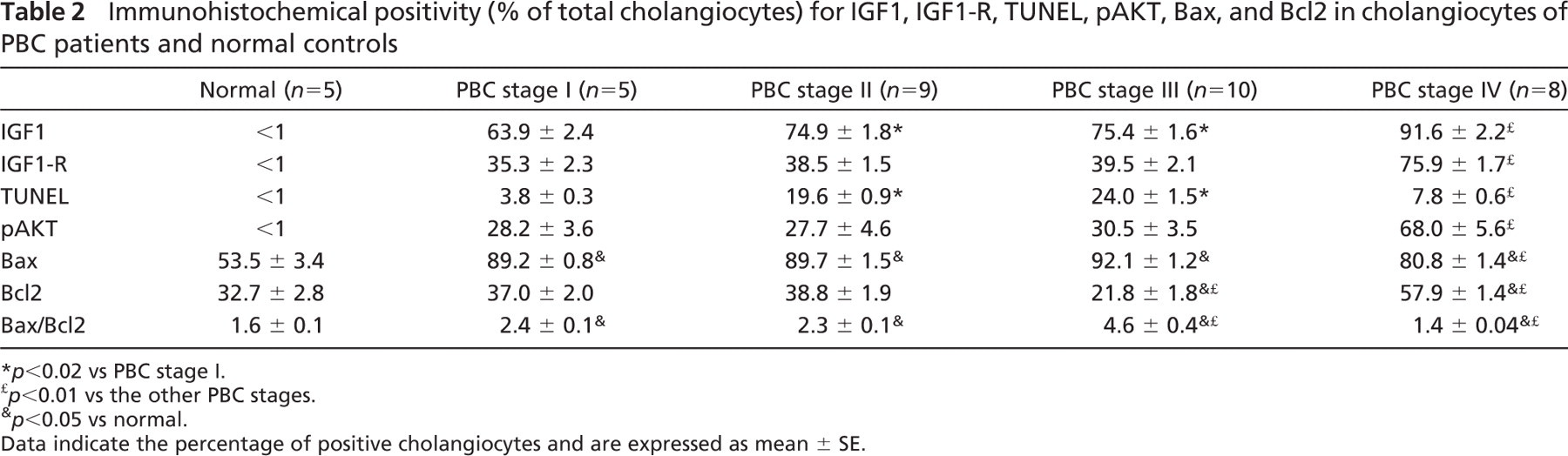
IHC analyses focus on interlobular bile ducts (20-100 μm in diameter) without significant differences between the different PBC stages and normal controls in the diameter of investigated ducts. IHC for IGF1 and IGF1-R in the liver of PBC patients and normal controls is shown in Figure 1 and quantitative data, expressed as percentage of positive cholangiocytes, are reported in Table 2. Less than 1% cholangiocytes of normal liver were positive for IGF1 and IGF1-R in the IHC study (Figures 1A and 1C). In contrast, most cholangiocytes of PBC patients were positive, with a staining located prevalently at cytoplasmic level for both IGF1 and IGF1-R (Figures 1B and 1D). When the different stages of PBC were compared, the positivity for IGF1 increased from 64% cholangiocytes in PBC stage I to ∼75% in stage II and III (p<0.02 vs stage I, Table 2) and to >90% in stage IV (p<0.01 vs stage I, II, or III, Table 2). IHC positivity for IGF1-R involved ∼35% to ∼40% cholangiocytes in PBC stages I, II, and III and markedly increased in PBC stage IV where it involved >75% of cholangiocytes (p<0.01 vs stages I, II, and III; Table 2). To investigate the signal transduction pathway activated by IGF1, we evaluated the IHC for pAKT, which was positive in <1% cholangiocytes of the normal liver (Figure 2A). In PBC, the pAKT-positive staining (Figure 2B) involved ∼30% of cholangiocytes in stages I, II, and III, whereas in PBC stage IV pAKT positivity markedly increased involving 70% cholangiocytes (p<0.01 vs PBC stages I, II, or III, Table 2). By considering all PBC stages, the percentage of cholangiocytes positive for pAKT was positively correlated with that of IGF1 (r = 0.72, p<0.03) or IGF1-R (r = 0.76, p<0.02).
Baseline characteristics of patients
p<0.05 vs normal.
p<0.05 vs stage I and II.
Data are mean ± SE; normal values: alkaline phosphatase, 98-270 U/liter.
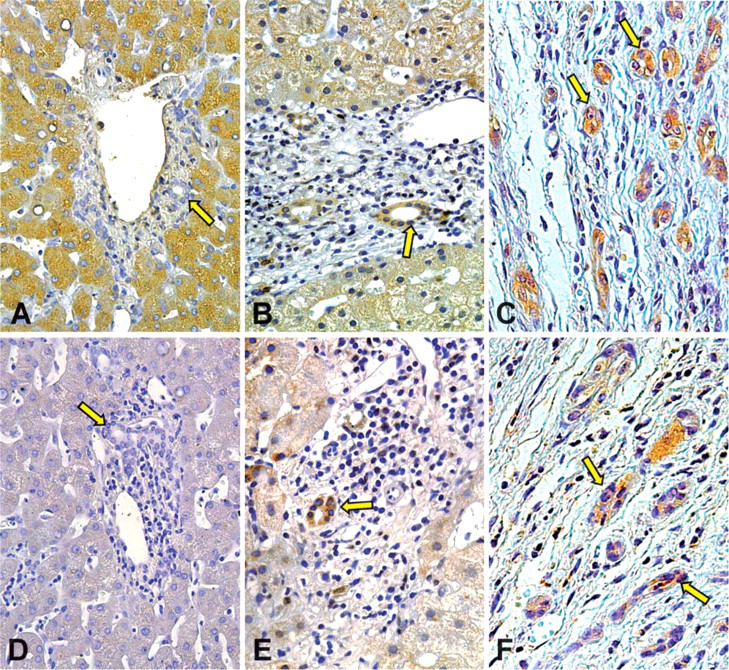
Immunohistochemistry for IGF-1 and IGF1-R in liver section of normal subject and PBC patient. Bile ducts (arrows) from normal liver
We next investigated IHC expression of Bcl2 and Bax (Figure 3), which are two main players in the regulation of apoptotic cascade (Willis et al. 2003). In the normal liver, ∼33% cholangiocytes were positive for the antiapoptotic protein Bcl2 (Figure 3A; Table 2), and this was slightly increased in PBC stage I (37%) and stage II (39%). In contrast, in PBC stage III (Figure 3B), positive staining for Bcl2 significantly declined (22%, p<0.05 vs normals, PBC stage I or stage II), whereas in PBC stage IV, Bcl2 positivity was markedly increased involving 58% cholangiocytes (p<0.01 vs normals or other PBC stages). A different trend was observed for the proapoptotic Bax, whose staining involved 53% cholangiocytes in the normal liver (Figure 3C), increased to ∼90% in PBC stages I, II, and III (p<0.01 vs normals; Figure 3D), and significantly decreased in the terminal PBC stage IV (81%, p<0.05 vs PBC stages I, II, and III). The Bax/Bcl2 ratio has been considered an expression of a balance between pro- and antiapoptotic proteins in the cell and, in fact, by considering all PBC stages, it was found to be directly correlated (r = 0.78, p<0.01, Figure 4) with TUNEL positivity. The Bax/Bcl2 ratio increased from 1.64 in normal cholangiocytes to 2.4/2.3 in PBC stages I and II (p<0.01 vs normal, Table 2) and reached the highest value in PBC stage III (4.6; p<0.01 vs normal, PBC stage I and stage II) when apoptosis is maximal (i.e., 24% TUNEL-positive cholangiocytes; Table 2). In PBC stage IV, however, the Bax/Bcl2 declined to 1.41 (p<0.01 vs PBC stages I, II, and III), and this coincided with a marked decline in the percentage of apoptotic cholangiocytes (i.e., 7.8% TUNEL-positive cholangiocytes; Table 2).
Immunohistochemical positivity (% of total cholangiocytes) for IGF1, IGF1-R, TUNEL, pAKT, Bax, and Bcl2 in cholangiocytes of PBC patients and normal controls
p<0.02 vs PBC stage I.
p<0.01 vs the other PBC stages.
p<0.05 vs normal.
Data indicate the percentage of positive cholangiocytes and are expressed as mean ± SE.
By considering the ductopenic stages of PBC (i.e., stage III and stage IV), expression of IGF1, IGF1-R, or pAKT in cholangiocytes of PBC patients was directly correlated with the antiapoptotic protein Bcl2 (r = 0.87, 0.86, 0.88; p<0.01) and inversely correlated with the proapoptotic protein Bax (r = −0.73, −0.76, −0.75; p<0.02) and with the Bax/Bcl2 ratio (r = −0.81, −0.80, −0.82; p<0.01).
Discussion
The main findings of this study indicate that (1) cholangiocytes lining interlobular bile ducts of PBC patients, but not normal subjects, showed a significant positivity for IGF1, IGF1-R, and pAKT, which increases from stage I to stage IV; (2) >70% of cholangiocytes surviving in the terminal ductopenic stage of PBC were positive for IGF1, IGF1-R, and pAKT; (3) in the ductopenic stages III and IV of PBC, expression of IGF1, IGF1-R, and pAKT in cholangiocytes was directly correlated with the antiapoptotic protein Bcl2 but inversely correlated with the proapoptotic protein Bax and with Bax/Bcl2 ratio and with TUNEL positivity.
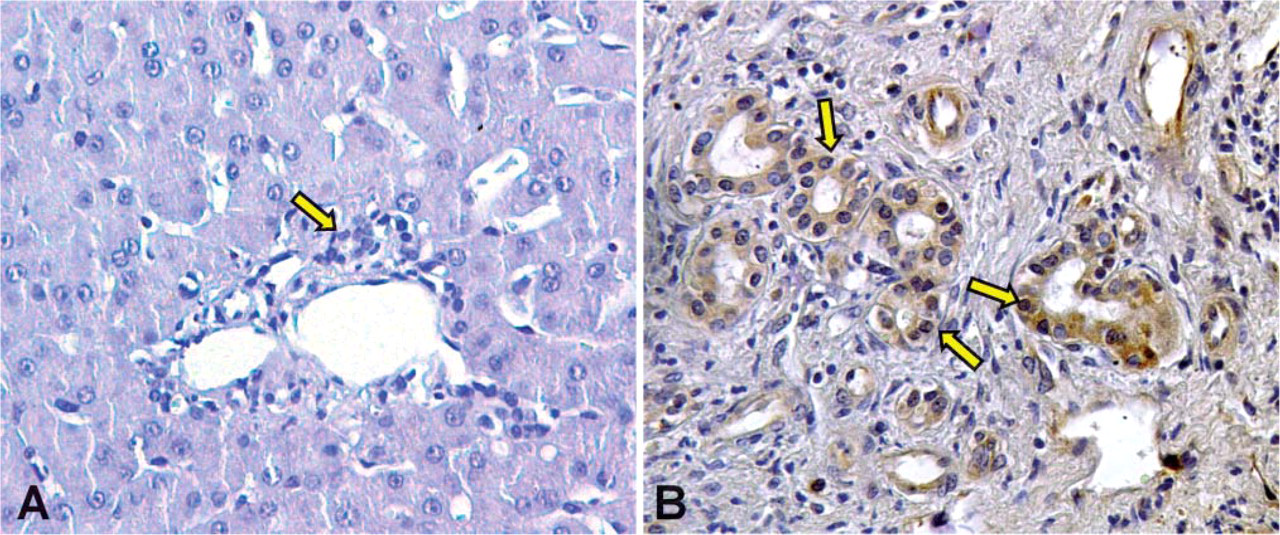
Immunohistochemistry for pAKT in liver section of normal subject and PBC patient. Bile ducts (arrows) from normal liver
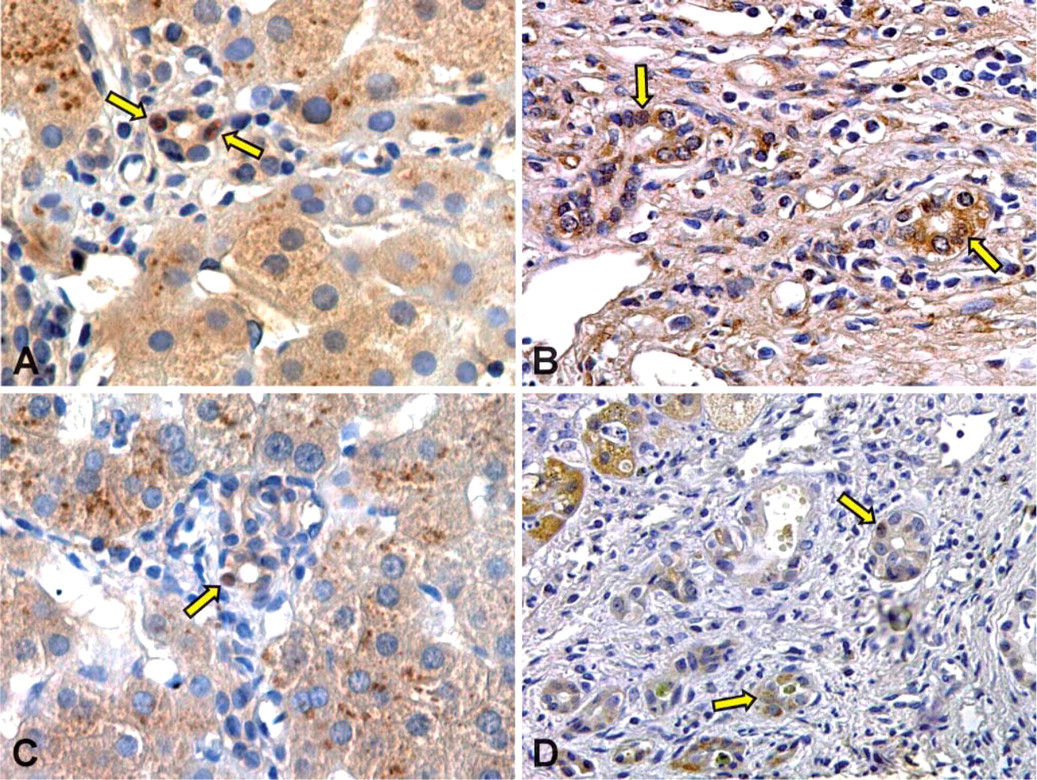
Immunohistochemistry for Bax and Bcl2 in liver section from normal subject and PBC patient. Bile ducts from normal (arrows in
IGF1 is a circulating peptide hormone and locally acting growth factor with endocrine, paracrine, and autocrine functions (Ge and Rudikoff 2000; Tumber et al. 2000; van Golen and Feldman 2000; Hong et al. 2001; Agnusdei and Gentilella 2005). A great deal of evidence shows that signaling activated by IGF1 and IGF1-R plays a powerful role in cell survival and prevention of apoptotic cell death, and this was documented in different cell types including osteoblasts (Tumber et al. 2000), melanoma cells (Ge and Rudikoff 2000), cardiac myoblasts (Hong et al. 2001), neuronal cells (van Golen and Feldman 2000), and different epithelia (Ahmad et al. 1999). Cellular effects of IGF-I are mediated through binding to the IGF1-R, although IGF-binding proteins also may modulate the action of IGF1, controlling the amount of free IGF1 that can bind to its receptor (Baxter 2000). Following ligand-induced stimulation of tyrosine kinase activity, autophosphorylation of tyrosine residues of the IGF1-R occurs, leading to phosphorylation of other sites in the receptor and associated substrate proteins which, in turn, activate downstream signaling (O'Connor et al. 1997; Chen et al. 1998). The best defined pathway by which IGF1-R signaling prevents apoptosis and favors cell survival is mediated by PI3K/AKT pathway. After tyrosine phosphorylation by the IGF1-R of insulin receptor substrate-1 (IRS-1), PI3K is activated by binding of the SH2 domain of its regulatory subunit to IRS-1, leading to an increase in phosphatidylinositol 3,4,5-trisphosphate (PIP3) (Dudek et al. 1997). Elevated PIP3 binds to the pleckstrin homology domain of Akt, which plays a key role in the prevention of apoptotic cell death because it phosphorylates and inactivates several proteins that are involved in the apoptotic cascade (Dudek et al. 1997). A primary target is the Bcl2 family member, Bad (Leverrier et al. 1999; Chrysis et al. 2001), which in its non-phosphorylated state is located at the mitochondrial membrane where it interacts with Bcl2, thus blocking the antiapoptotic functions of this protein. Once phosphorylated by Akt, Bad associates with cytosolic proteins and becomes unable to interfere with Bcl2 (Heck et al. 1999; Peruzzi et al. 1999). The activated pAkt also increases the levels of antiapoptotic proteins including Bcl2 and Bcl-x and several extracellular matrix adhesion molecules (Leverrier et al. 1999; Chrysis et al. 2001) and of the antiapoptotic transcription factor NF-kB (Leverrier et al. 1999) and may stimulate as yet unidentified coactivators that reduce the activity of the proapoptotic molecule, p53 (Yamaguchi et al. 2001). Downregulation of p53 is particularly relevant to preventing apoptosis because p53 can cause a decrease in expression of the IGF1-R and upregulate the proapoptotic Bax protein (Grimberg 2000). Given the relevance in favoring cell survival, exogenous IGF1 administration has been attempted and is currently under evaluation in different human pathological conditions including liver cirrhosis (Agnusdei and Gentilella 2005; Conchillo et al. 2005; Fernandez et al. 2005).
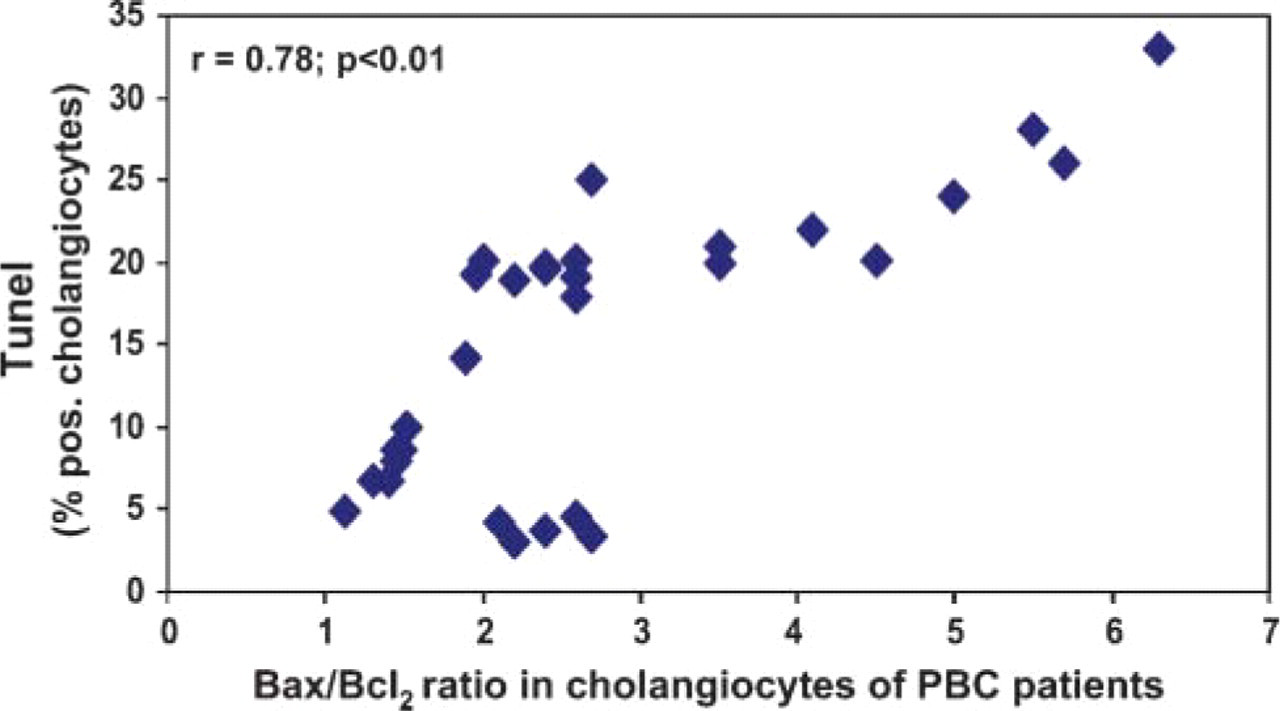
Relationship between Bax/Bcl2 ratio and TUNEL positivity in cholangiocytes of patients with different stages of PBC. By considering cholangiocytes lining interlobular bile ducts of all PBC stages, the Bax/Bcl2 ratio is directly correlated (r = 0.78, p<0.01) with TUNEL positivity.
We have recently shown in experimental studies that the IGF1 system and PI3kinase/Akt pathway are activated and drive cholangiocyte proliferation and resistance against apoptosis in obstructive cholestasis induced by bile duct ligation (Alvaro et al. 2005) and in determining the in vitro resistance of hepatocytes and cholangiocytes against bile salt (BS)-induced apoptosis (Drudi Metalli et al. 2005). In the present study, we focus on PBC, a disease that progressively evolves toward ductopenia with disappearance of interlobular bile ducts (Desmet et al. 1995,1998a,b; Kuroki et al. 1996; Boyer 1997; Nakanuma et al. 2000; Harada et al. 2001, Tinmouth et al. 2002; Alvaro et al. 2004; Kaplan and Gershwin 2005). Most studies demonstrated that apoptosis is a major mechanism of cholangiocyte death during PBC (Kuroki et al. 1996; Nakanuma et al. 2000; Harada et al. 2001, Tinmouth et al. 2002; Alvaro et al. 2004), and this was also confirmed by the present findings. In a previous study we showed that the progression toward ductopenia is caused by a relative predominance of cholangiocyte apoptosis toward proliferation that specifically occurs in the ductopenic PBC stage III and IV (Alvaro et al. 2004). However, while in stage III, both proliferation and apoptosis are markedly enhanced. In terminal stage IV they decline, indicating a sort of turning off of damaging processes in residual ducts (Alvaro et al. 2004). We focus our IHC study on interlobular bile ducts, which are the exclusive target of damage in PBC (Desmet et al. 1995,1998a,b), and the mean diameter of bile ducts investigated was similar among the different PBC stages and between PBC and controls to allow a reliable comparison of the measured parameters. Furthermore, the study was performed in postmenopausal PBC patients and controls, thus excluding interference of cyclical changes of estrogen serum levels on the IGF1 system. This is of relevance given the well-known interaction between estrogens and IGF1 (Kahlert et al. 2000). The major finding of our study is that cholangiocytes lining interlobular bile ducts of PBC patients, but not normal subjects, express IGF1, IGF1-R, and pAKT. IHC negativity of IGF1 and IGF1-R in normal subjects probably depends upon the low amount of protein expressed under normal conditions where also hepatocytes, the main cell type of the organism producing IGF1 (Baxter 2000), are almost negative. We analyzed IHC expression of the phosphorylated form of AKT to evaluate the state of activation of the main intracellular transduction pathway driving IGF1 effect on cell survival and resistance against apoptosis (Ge and Rudikoff 2000; Tumber et al. 2000; van Golen and Feldman 2000; Hong et al. 2001; Agnusdei and Gentilella 2005). Consistently, pAKT expression in cholangiocytes was linearly correlated with the expression of IGF1 and IGF1-R. Most importantly, in the terminal ductopenic stages of the disease, >70% of cholangiocytes express IGF1, IGF1-R, and pAKT, indicating that almost only cells displaying activation of IGF1 system are selected and survive against the immunomediated damage because in these cells apoptosis is prevented by upregulation of the IGF1 system. This was further confirmed by the inverse correlation between IGF1, IGF1-R, pAKT, and the Bax/Bcl2 ratio observed in the ductopenic stages of PBC. Bax and Bcl2 and their ratio were investigated as major players in the complex loop of agents modulating the apoptotic machinery (Ortiz et al. 2000; Martinez-Arribas et al. 2003; Alvaro et al. 2005). The Bax/Bcl2 ratio reached the highest value in PBC stage III (i.e., 4.6) when apoptosis is maximal (i.e., 24% TUNEL-positive cholangiocytes), whereas in PBC stage IV, where TUNEL positivity involves only 7.8% of cholangiocytes, the Bax/Bcl2 ratio reached the lowest level among the different PBC stages. In many different cell types, the IGF1 system directly modulates the expression and activity of the antiapoptotic protein Bcl2 (Leverrier et al. 1999; Chrysis et al. 2001). In cholangiocytes of PBC patients we also confirmed such type of correlation. In fact, the percentage of cholangiocytes expressing Bcl2 was markedly increased in the terminal stages of the disease, and this was associated and directly correlated with IGF1, IGF1-R, and pAKT expression. This indicates that cholangiocytes, where the IGF1 system is activated, display an attitude toward survival rather than apoptosis and that the upregulation of the IGF-1 system prevents cholangiocyte apoptosis. Unfortunately, double staining in the same cells for markers of IGF1 system activation and markers of survival/apoptosis was not possible in the present study due to material insufficiency. However, when evaluated morphologically, >90% of cholangiocytes positive for IGF1, IGF1, or pAKT lack features of apoptosis (cell shrinkage, chromatin condensation), whereas >90% of cholangiocytes negative for IGF1, IGF1-R, and pAKT display morphological signs of apoptosis. In PBC, hepatocytes also showed positivity for IGF1, but this is not surprising because these cells are the main source of circulating IGF1 and are damaged by BS accumulation as the consequence of cholestasis (Baxter 2000).
Several experimental and human studies indicate that, in cells expressing estrogen receptors, a synergism exists between estrogen and IGF1 in modulating survival and resistance against apoptosis, with a preferential involvement of the estrogen receptor α subtype (Kahlert et al. 2000; Cossette et al. 2002; Alvaro et al. 2005). This was also demonstrated in cholangiocytes (Alvaro et al. 2005). However, in the terminal stage IV of PBC, cholangiocytes lose the expression of the estrogen receptor α subtype and, therefore, the IGF1 system remains as a major pathway in promoting cell survival.
In conclusion, we have shown that the IGF1 system activation is a feature of biliary epithelium lining inter-lobular bile ducts of PBC patients and specifically that this represents a characteristic of almost all cholangiocytes surviving in the terminal ductopenic stages of the disease. In light of our findings, pharmacological or genetic strategies aimed to sustain the IGF1 system may help in delaying disease progression.
Footnotes
Acknowledgements
This work was supported by MIUR grants (to DA and AFA): PRIN 2003, 2005: #2003060137_002, 2003060498_002, 2005069739_003, 2005067975_002. E.G. and P.O. are supported by PRIN 2003, 2005, Biomedicina-Cluster C04-progetto n.ro 5 and ex 60% grants.
We thank F. Lucarelli and Cinzia Tesse for technical assistance in immunoblotting and Tracie Dornbusch for English editing.