
Abstract
Sphingosine kinase (SPHK) catalyzes sphingosine phosphorylation to form a bioactive lipid mediator, sphingosine-1-phosphate (S1P). In the current study, we report the presence of SPHK-1 in mouse spermatozoa. SPHK-1 was localized to the acrosomes of spermatozoa, and its expression was proven by RT-PCR and Western blot analysis. SPHK activity of mouse spermatozoa was 18.1 pmol/min/mg protein. Furthermore, we identified the presence of the S1P receptors S1P1, S1P2, S1P3, and S1P5, in mouse spermatozoa by RT-PCR. These results suggest that S1P produced by SPHK-1 would play a role in the acrosomal reaction through S1P receptors.
S
Recent studies have revealed that S1P plays a cytoprotective role for both female and male germ cells. In the ovary, S1P was found to effectively block apoptosis induced by anti-cancer therapy (Morita et al. 2000) and to protect germ cells from radiation without propagating genomic damage (Paris et al. 2002). Likewise, in the testis, S1P inhibits germ cell apoptosis spontaneously induced by the culture of seminiferous tubules (Suomalainen et al. 2003) and irradiation (Otala et al. 2004). Therefore, the aim of the current study was to examine the expression and localization of the S1P-generating enzyme, SPHK-1, in mice germ cells.
Mice were maintained in the animal facility, and the experiments during this study using 8-week-old mice (C57BL/6 strain) were performed in accordance with the animal experimental guidelines of Gifu International Institute of Biotechnology. Rabbit anti-SPHK-1 antibody was prepared as described previously (Murate et al. 2001).
Immunofluorescent staining was performed as described previously (Matsumoto et al. 2002). Briefly, the frozen sections (7-μm-thick) of ovary and testis or spermatozoa on silanized slides (DAKO Cytomation, Copenhagen, Denmark) were fixed with 4% paraformaldehyde in 0.1 M sodium phosphate buffer (pH 7.4) for 30 min at room temperature (RT). After washing with 0.1% Tween 20 in phosphate-buffered saline (PBS-T), the specimens were blocked with blocking reagent containing 1.0% bovine serum albumin and 50% Block Ace (Dainihon-seiyaku, Osaka, Japan) for 1 hr at RT. The specimens were incubated with anti-SPHK-1 antibody overnight at 4C, followed by AlexaFluor-488-conjugated goat anti-rabbit IgG F(ab') (Invitrogen; Carlsbad, CA). After washing with PBS-T, the sections of ovary and testis were reacted with 4,6-diamidino-2-phenylindole for nuclear staining, and spermatozoa were reacted with AlexaFluor-594-conjugated peanut lectin for acrosome staining (Aviles et al. 1997). As negative controls, we used the specimens reacted with the secondary antibody alone. The stained specimens were observed with a fluorescence microscope (Olympus BX-51; Olympus, Tokyo, Japan).
To examine the SPHK-1 protein level by Western blot analysis, spermatozoa isolated from the tail region of epididymis were homogenized with chilled lysis buffer containing 10 mM Tris-HCl (pH 7.4), 1% NP-40, 0.1% deoxycholic acid, 0.1% SDS, 150 mM NaCl, 1 mM EDTA, and 1% Protease Inhibitor Cocktail (Sigma; St Louis, MO) and kept for 30 min on ice. After centrifugation at 14,000 rpm for 20 min at 4C, the resulting supernatant was collected for Western blot analysis. Protein content was measured with a DC Protein assay kit (Bio-Rad; Hercules, CA). Ten μg of protein was used for Western blot analysis (Matsumoto et al. 2003).
PCR primers
Real-time RT-PCR was performed on a LightCycler (Roche; Mannheim, Germany) using a LightCycler Fast Start DNA Master SYBR Green I kit (Roche) according to the manufacturer's instructions. Two μg of total RNA was reverse transcribed by Super Script II RNase H-reverse transcriptase (Invitrogen) using oligo(dT) primer (Invitrogen). The prepared cDNA sample was purified by a PCR purification kit (QIAGEN; Hilden, Germany) and used for PCR. Primers for each gene are shown in Table 1.
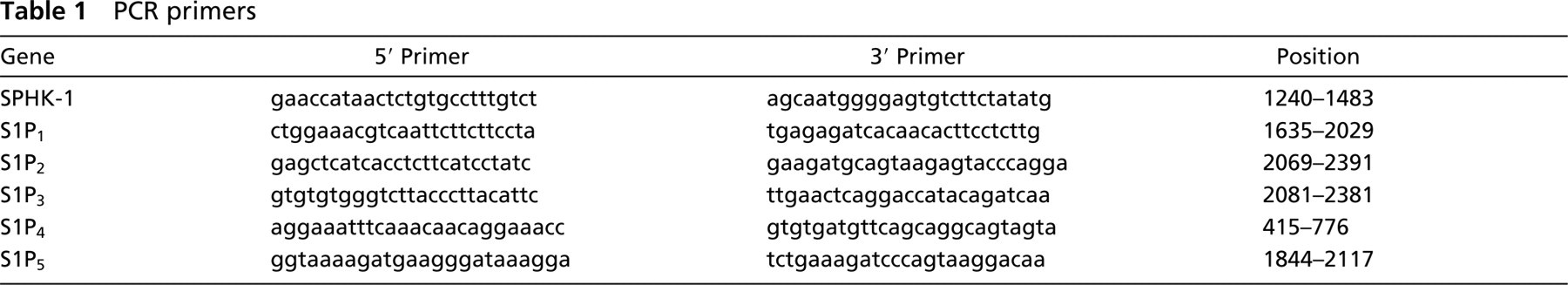
Immunofluorescent staining for SPHK-1 in mouse testis.
SPHK activity was measured as previously described (Murate et al. 2001). Briefly, 50 μg of protein was used for enzyme activity assay. The reaction was started by adding 0.05 μCi [3-3H]-sphingosine (American Radiolabeled Chemicals; St Louis, MO) and 1 mM ATP in a final volume of 50 μl. After incubation at 37C for 30 min, the reaction was terminated with 5 μl of 1 N HCl followed by 200 μl of chloroform: methanol: HCl (1000:200:1, v/v). After vortex, 62.5 μl of 2 M KCl and 62.5 μl of chloroform were added for phase separation. The separated chloroform phase was condensed by vacuum evaporation, applied on Silica Gel 60 thin-layer plates, and then developed with a solvent of butanol:ethanol:acetic acid:water (80:20: 10:20, v/v). Cold authentic sphingosine and S1P were added to each sample just before chromatography, and their position was identified with Ninhydrin reagent. The spot corresponding to S1P was visualized by autoradiography, scraped off the plates, and its radioactivity was measured in a liquid scintillation counter.
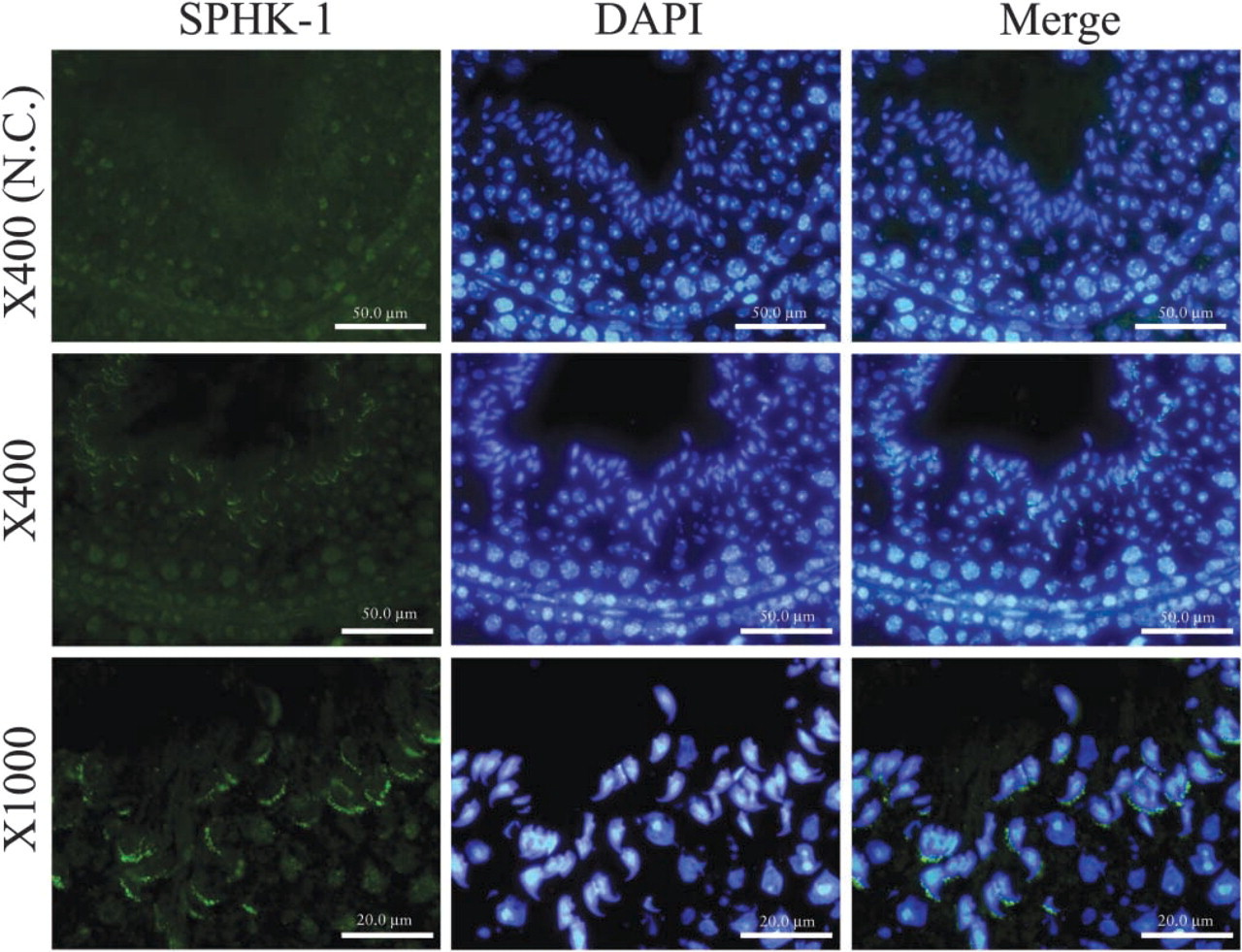
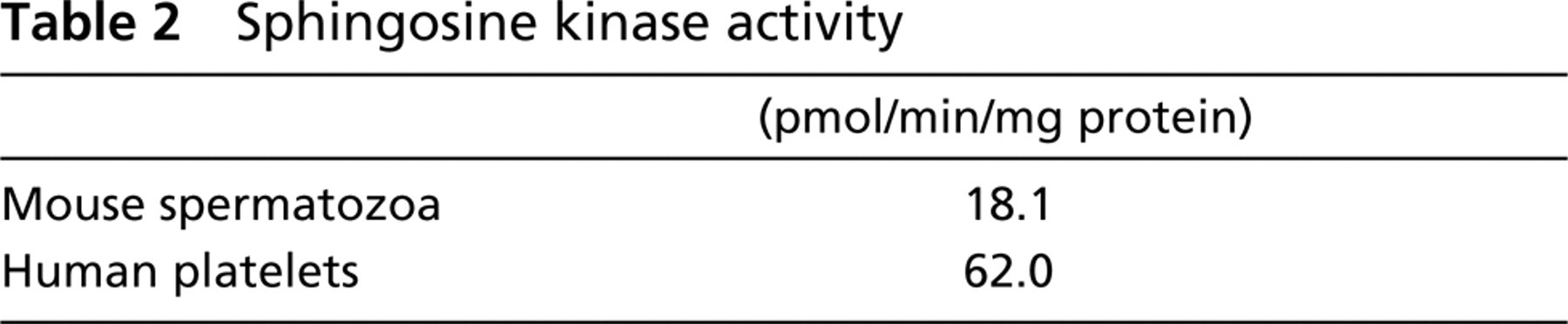
To examine the expression of SPHK-1 in mouse germ cells, we carried out immunofluorescent staining of mice ovary and testis. Although SPHK-1 was undetectable in female germ cells (data not shown), it was detected in male germ cells. SPHK-1 was observed to be localized to the head of spermatozoa but not in other cells of seminiferous tubules, such as spermatogonia, spermatocytes, and Sertoli cells (Figure 1). Next, we investigated expression and localization of SPHK-1 in matured spermatozoa. Matured spermatozoa were collected from the tail region of epididymis for RT-PCR, Western blot analysis, and immunofluorescent staining. The expressions of mRNA and protein of SPHK-1 were confirmed in matured spermatozoa (Figures 2A and and 2B). Double staining for SPHK-1 and acrosome revealed that SPHK-1 was localized to acrosomes (Figure 3C). Furthermore, we measured SPHK activity in matured spermatozoa. As a positive control, we used human platelets because of the high SPHK activity (Banno et al. 1998). The SPHK activity of spermatozoa was lower compared with human platelets (Table 2).
Examination of SPHK-1 expression in mouse spermatozoa. Spermatozoa were collected from the tail region of epididymis.
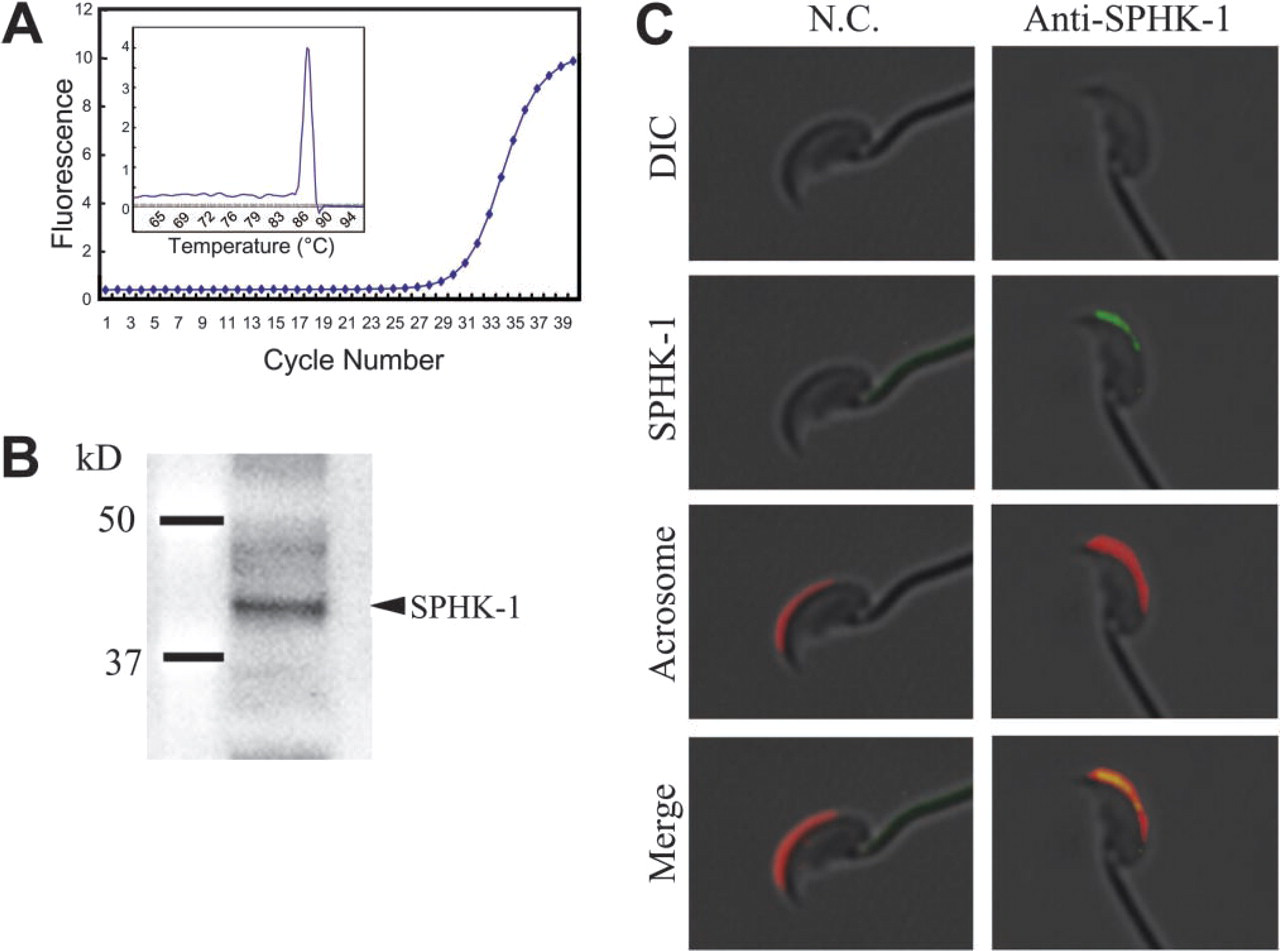
Real-time RT-PCR analysis of S1P receptors, S1P1-S1P5, in mouse spermatozoa. Spermatozoa were collected from the tail region of epididymis. The PCR products were evaluated by melting curve (inserts).
Sphingosine kinase activity
Acrosomes lie in front of the nuclei of the sperm head and contain enzymes to lyse the outer coverings of the oocyte during early fertilization (acrosomal reaction). Our finding indicating the localization of SPHK-1 in acrosomes suggests involvement of SPHK-1 in the acrosomal reaction.
S1P produced by SPHK is a ligand for G-protein-coupled receptors (S1P1-S1P5), and it has become clear that many important biological functions of S1P are mediated through these receptors (Spiegel and Milstien 2003). To gain insight into the functions of S1P in mouse spermatozoa, we have carried out RT-PCR for detection of the S1P receptors. Figure 3 shows that matured spermatozoa expressed mRNAs of other S1P receptors, except for S1P4. Thus, it is likely that S1P functions through its receptors.
Research on S1P signaling has attracted increasing attention over the last few years, and has revealed the role of S1P in a variety of physiological processes, such as cell growth and apoptosis. The existence of both SPHK-1 and S1P receptors in mouse spermatozoa indicates that S1P produced by SPHK-1 plays important roles in spermatozoa. Recent studies have shown a role of S1P in protection against cell death of male germ cells (Suomalainen et al. 2003; Otala et al. 2004), implying that S1P exerts a cytoprotective role in spermatozoa.
Recently, it has been revealed that spermatozoa deliver their own RNAs to the oocyte during fertilization, suggesting that sperm RNAs are important in early zygotic and embryonic development (Ostermeier et al. 2004). Morita et al. (2000) have reported the ability of S1P to suppress oocyte apoptosis. Consequently, mRNAs of SPHK-1 and S1P in spermatozoa might be delivered to the oocyte and function in the zygote.
In conclusion, our present study has demonstrated the localization of SPHK-1 in spermatozoan acrosomes and the expression of mRNAs of S1P receptors in spermatozoa. These results provide important information for understanding the roles of S1P in germ cells.
Footnotes
Acknowledgements
This work was supported in part by a Grant-in-Aid for Scientific Research on Priority Areas (B) (16390098) from the Ministry of Education, Culture, Sports, Science and Technology of Japan.