
Abstract
The macrophage-derived neutrophil chemotactic factor (MNCF) is a
N
Few recent studies show that lectins can attract neutrophils. Lectins can act as cell recognition mediators in a wide range of biological systems because of their great versatility and diversity in binding to carbohydrates (Sharon and Lis 1989). Plant lectins, such as KM+ (Santos-de-Oliveira et al. 1994), and mammalian lectins, such as macrophage-derived neutrophil chemotactic factor (MNCF) (Dias-Baruffi et al. 1995a,b) and galectin-3 (Sano et al. 2000; Sano and Liu 2001), can directly attract leukocytes and induce their movement. The neutrophil migration-inducing activity of MNCF has two unique characteristics. First, the neutrophil recruitment triggered in vivo by MNCF occurs even in animals pretreated with dexamethasone (Cunha and Ferreira 1986; Dias-Baruffi et al. 1995a,b). Second, incubation of MNCF with
In the present study, we demonstrate that MNCF binds to glycans of a GPCR on the surface of human neutrophils and induces neutrophil polarization. Following incubation of neutrophils with MNCF, the lectin bound to the cell surface and was rapidly detected in the cytoplasm, indicating internalization of lectin-glycosylated ligand complexes. The response of the neutrophils to MNCF is comparable to that seen with the binding of known neutrophil attractants to their specific receptors. This report provides evidence for the significant role animal lectins play in the inflammatory response.
Materials and Methods
Neutrophil Purification
Heparinized human blood from healthy volunteers was layered on a neutrophil isolation medium (Cardinal Associates; Santa Fe, NM) density gradient and centrifuged at 400 × g for 30 min. The neutrophils were washed by centrifugation and suspended in RPMI medium (Gibco-BRL, Life Technologies; Gaithersburg, MD), at a concentration of 106 cells/ml. Samples were subjected to hypotonic lysis to eliminate remaining erythrocytes. Resulting preparations were 98% pure, and more than 95% of the neutrophils were viable as measured by trypan blue.
Preparation of the Lectin MNCF
MNCF was obtained and purified as previously described (Cunha and Ferreira 1986; Dias-Baruffi et al. 1995a). Briefly, the supernatant from LPS-stimulated rat macrophage monolayers was submitted to affinity chromatography on an agarose-
Polarization Assays
Human neutrophils were plated at 106 cells/ml in RPMI medium. The neutrophils were incubated with MNCF (10 μg/ml RPMI), fMLP (10−6 M/well), casein (15 μg/ml RPMI), or RPMI medium alone for 45 min at 37C. In some experiments, the attractants were incubated with 0.2 M
Transmission Electron Microscopy
Samples were rinsed twice in PBS and once in 0.1 M cacodylate (EM Sciences; Fort Washington, PA) buffer (pH 7.4) and then fixed at room temperature for 40 min in 2% glutaraldehyde, 2% paraformaldehyde in the cacodylate buffer containing 0.05% CaCl2. After rinsing twice in PBS and once in cacodylate buffer, the cell preparations were embedded in 2% agar and postfixed at room temperature for 1 hr in 2% osmium tetroxide (EM Sciences). The samples were rinsed in distilled water, dehydrated through a graded series of ethanol and acetone, and embedded in Embed 812 (EM Sciences). Thin sections were cut with a diamond knife, mounted on copper grids, stained for 10 min in Reynolds’ lead citrate (Reynolds 1963), and stained for another 10 min in uranyl acetate. Specimens were examined with a Philips EM 208 (Philips Electron Optics; Eindhoven, The Netherlands).
Localization of the MNCF-Ligand Complexes on the Surface of Human Neutrophils
Human neutrophils were placed on coverslips coated with Biobond (EM Sciences), incubated with biotinylated MNCF (10 μg/ml RPMI) (Moreno et al. 2003) or RPMI medium alone at 37C for 30, 60, 120, and 180 min, and fixed with 2% formaldehyde at RT for 20 min. The cells were rinsed and then incubated with streptavidin-FITC (Pierce Chemical Co.) for 30 min. The slides were mounted in Fluormount-G (EM Sciences) and examined by confocal microscopy using a Leica TCS NT confocal microscope (Leica Microsystems; Heidelberg, Germany).
Internalization of MNCF–Ligand Complexes by Human Neutrophils
Non-fixed human neutrophils were placed on coverslips coated with Biobond (EM Sciences) and incubated with biotinylated MNCF (10 μg/ml RPMI) or with RPMI medium alone at 37C for 30, 60, 120, and 180 min. After being rinsed with PBS, the cells were treated for 30 min at 4C with 0.2 M
Chemotaxis Assays
Neutrophil migration was assayed in a modified Boyden chamber (a microchamber with 48 wells; NeuroProbe, Cabin John, MD) (Bignold 1988). One of the attractants, MNCF (10 μg/well), IL-8 (40 ng/well), or fMLP (10−6 M/well) was added to the lower wells of the chamber, separated from the upper wells by a 5-μm polycarbonate membrane filter (25 × 80 mm) (Poretics Corporation; Livermore, CA). After placing human neutrophils (5 × 104 cells/well) in the upper wells, the chambers were incubated for 45 min at 37C in a humidified incubator containing 5% CO2 in air. Cells that migrated through the entire width of the membrane were counted. Random migration was assessed by using RPMI medium in the lower chamber. To examine the role of GPCRs on the MNCF chemotactic activity, neutrophils were preincubated with pertussis toxin (PTx, 500 ng/ml; List Biological Laboratories, Campbell, CA) for 90 min at 37C, washed twice with RPMI medium, and added to the upper wells (5 × 104 cells/well). The cells treated with PTx had a similar viability to that of the untreated cells (>95%). Five fields were counted for each assay, and each sample was assayed in triplicate. All results were expressed as mean ± SD.
Scanning Electron Microscopy
Human neutrophils were incubated for 45 min at 37C with MNCF (10 μg/ml RPMI), which was preincubated for 45 min at 37C with or without 0.2 M
Statistical Analysis
Data are presented as the mean ± SD. Statistical determinations of the difference between means of experimental groups were performed using one-way ANOVA with Bonferroni posttest for multiple comparisons. Differences that provided p<0.001 were considered as being statistically significant. All experiments were performed at least three times, each one using neutrophils from a different donor.
Results
MNCF Induces Human Neutrophil Polarization
The ability of MNCF to induce neutrophil polarization, a well-known characteristic of neutrophil activation by chemoattractants (Sánchez-Madrid and Del Pozo 1999), was examined. In a previous study (Dias-Baruffi et al. 1995b), we showed that the dose-response curve for MNCF-induced neutrophil migration was bell shaped with a maximum response at 10 μg/ml, an observation that was confirmed in the present study. MNCF (10 μg/ml) was able to induce the polarization of 85% of the neutrophils. The chemoattractants fMLP and casein induced polarization in 70% and 78% of neutrophils, respectively, whereas only 18% of cells incubated with medium alone were polarized (Figure 1). To verify if neutrophil polarization induced by MNCF was dependent on sugar recognition, assays were performed using MNCF preincubated with either of the monosaccharides
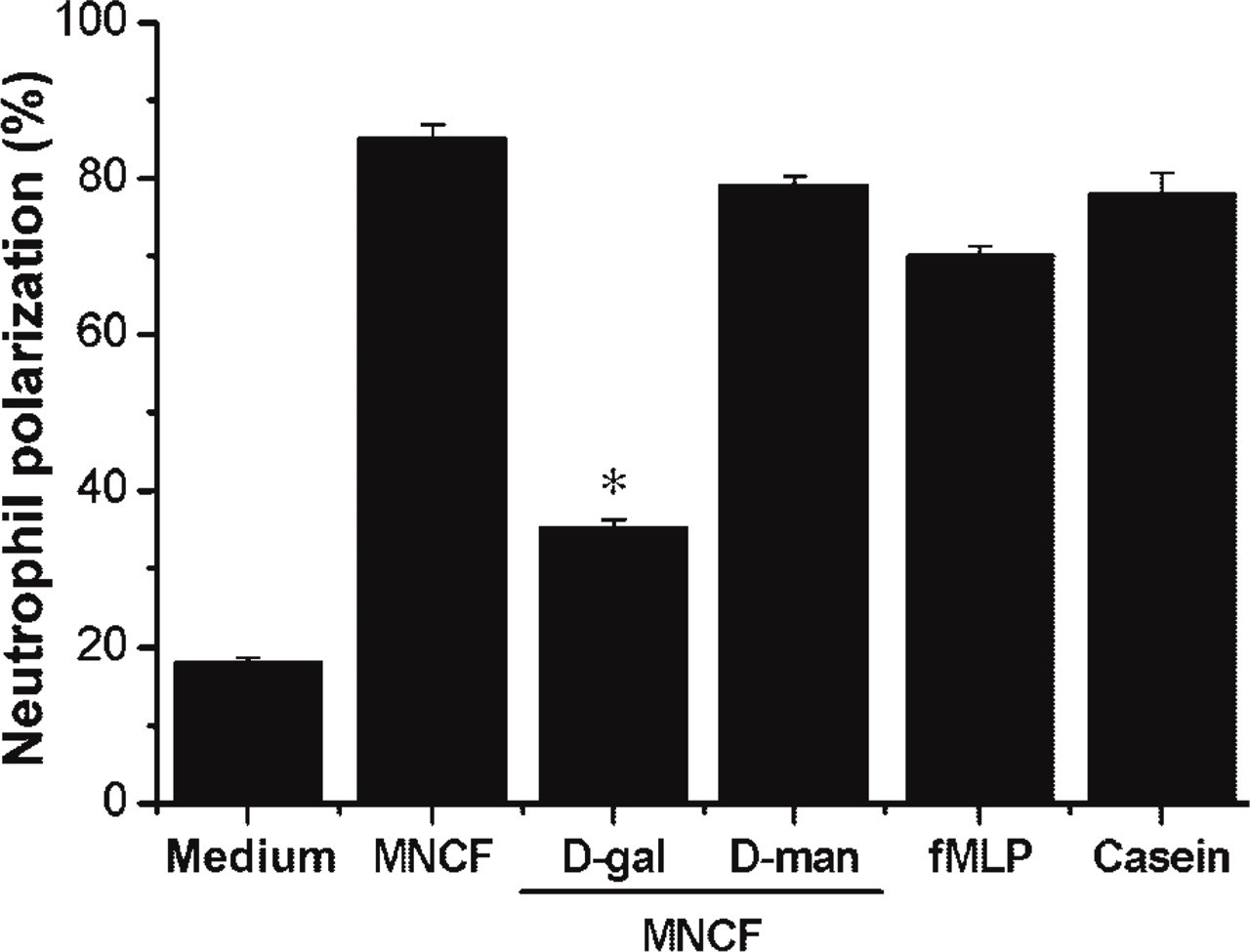
MNCF induces neutrophil polarization, an effect inhibited by
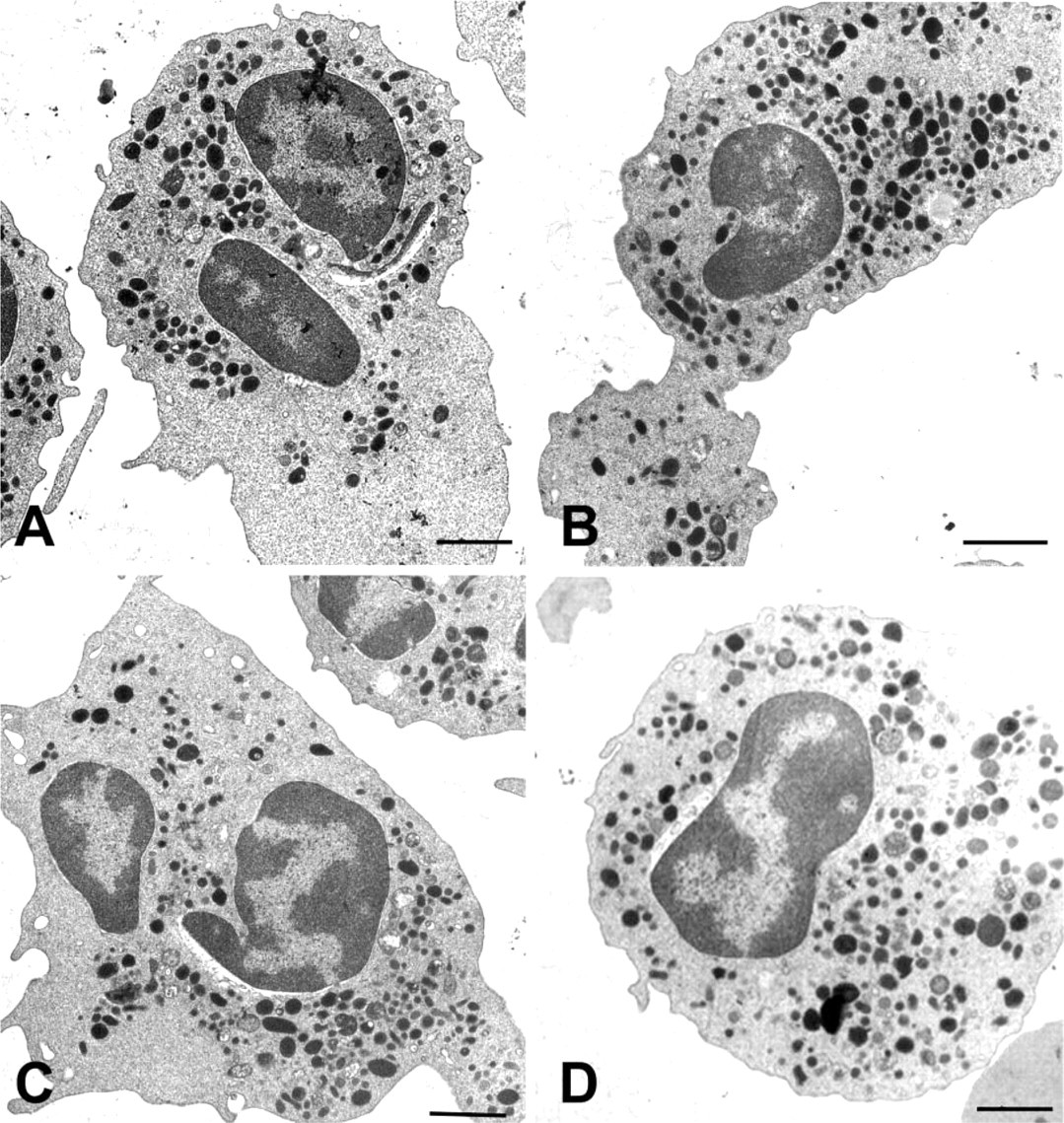
MNCF induces ultrastructural changes in neutrophils. (
Transmission electron microscopy was also used to examine the polarization of neutrophils induced by MNCF. The cells incubated with MNCF (Figure 2A) were elongate, contained abundant electron-dense granules in the cytoplasm, and had lamellipodium free of organelles. Similar effects on neutrophil morphology were seen in cells treated with either fMLP (Figure 2B) or casein (Figure 2C). Neutrophils incubated with RPMI medium only (Figure 2D) maintained their spherical shape with characteristic granules.
MNCF Binds to the Neutrophil Cell Surface and Is Internalized
To visualize the MNCF bound to glycoligands on the surface of human neutrophils, cells were incubated with biotinylated MNCF followed by streptavidin–FITC. After a 30-min incubation, confocal microscopy showed that biotinylated MNCF was evenly bound to the cell surface (Figure 3A). Fluorescent labeling on the cell surface clearly decreased with longer incubation periods (Figures 3C, 3E, and 3G).
Lectin internalization was then investigated. Neutrophils were incubated with biotinylated MNCF for varying times, rinsed with 0.2 M
Pertussis Toxin (PTx) Inhibits Neutrophil Migration Induced by MNCF
To determine if MNCF induces neutrophil migration by interacting with GPCRs, we investigated the effect of PTx on cell polarization and migration induced by this lectin. Pretreatment of neutrophils with PTx abolished the MNCF-induced neutrophil migration (Figure 4). The original response to MNCF (97.87 ± 1.337) decreased drastically (9.94 ± 0.665) and was similar to that found in the absence of the stimuli (13.06 ± 0.72). As expected, IL-8 and fMLP were also unable to induce migration of neutrophils that had been pretreated with PTx (Figure 4). This finding is in agreement with other studies showing that chemokines and other known neutrophil attractants trigger directional cell movement by binding to GPCRs (Baggiolini et al. 1997; Locati and Murphy 1999; Mantovani 1999).
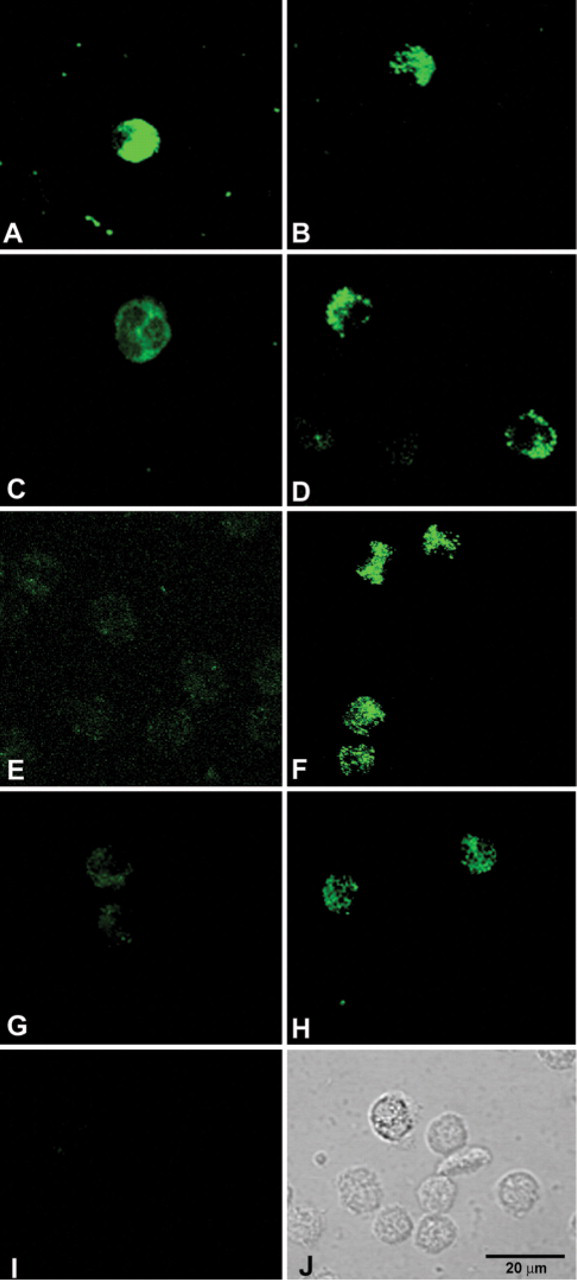
Kinetics of MNCF binding to the neutrophil surface and MNCF internalization. Isolated human neutrophils were incubated with biotinylated MNCF and labeled with streptavidin–FITC. Confocal microscopy showed that the MNCF was evenly distributed on the cell surface at 30 min of incubation (
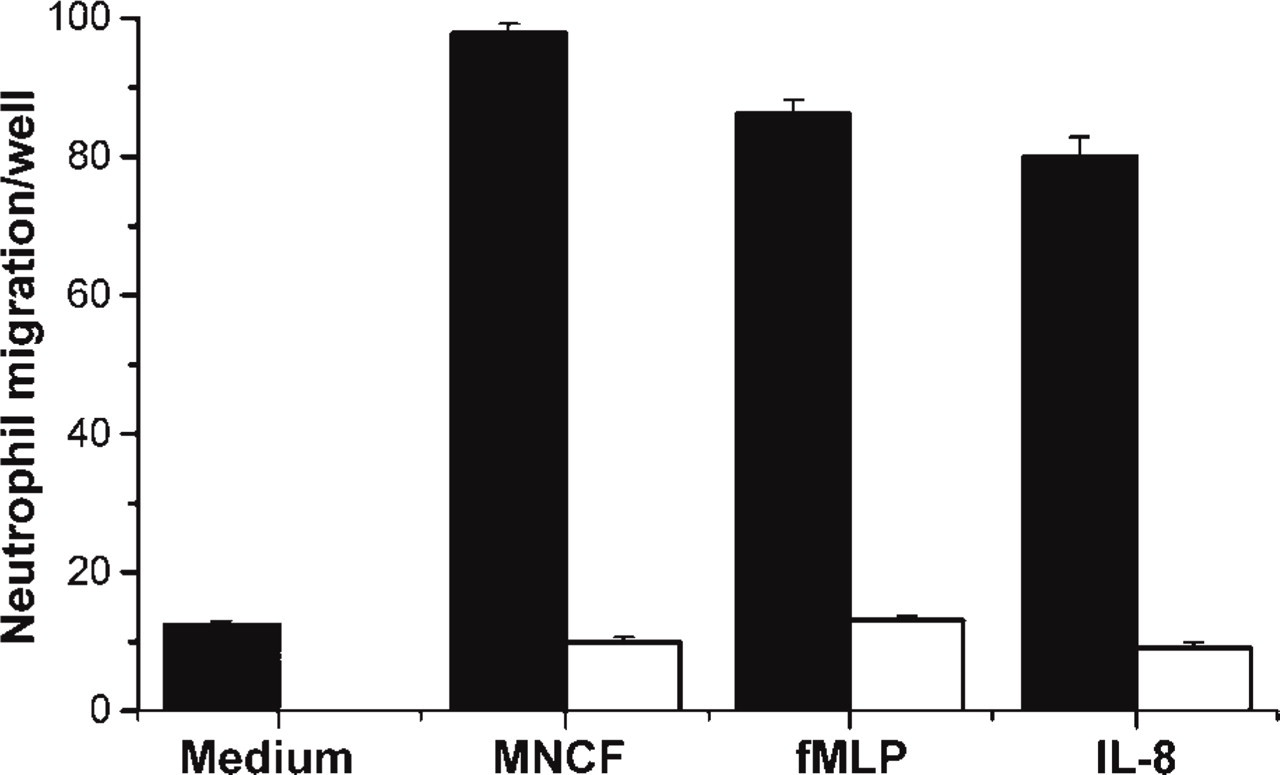
Pertussis toxin inhibits MNCF-induced neutrophil migration. A neutrophil chemotaxis assay was carried out in a 48-well Boyden microchamber. MNCF (10 μg/well), IL-8 (40 ng/well), or fMLP (10−6 M/well) was placed in the wells of the lower chamber; medium (RPMI) was used as negative control. The wells of the upper chamber received isolated human neutrophils that were preincubated in medium with (filled bars) or without (open bars) pertussis toxin (PTx 500 ng/ml). The number of cells that migrated through the filter separating the two chambers was scored in each field. The figure is representative of the results of three different experiments, each one using neutrophils from a different donor. All results were expressed as mean ± SD.
Scanning electron microscopy showed that after a 30-min incubation with MNCF the neutrophils were spread over the substratum, exhibited deep membrane ruffling, and lost their fine microvilli (Figure 5A). When neutrophils were pretreated with PTx and then incubated with MNCF, no morphological changes were observed (Figure 5B). The neutrophils maintained their spherical shape, and their surface was covered with microvilli. Neutrophils and neutrophils pretreated with PTx incubated with RPMI medium only also showed no morphological changes (Figures 5C and 5D).
Discussion
In the present study, we have expanded our characterization of the inflammatory properties of the mammalian lectin MNCF, which is secreted by activated macrophages (Dias-Baruffi et al. 1995a,b; Moreno et al. 2003). When MNCF binds to the cell surface, it induces neutrophil polarization and migration. This neutrophil activation, as well as migration, can be inhibited by exposure to pertussis toxin, suggesting that MNCF acts through a GPCR. MNCF induces neutrophil migration by binding to
Following short exposures to MNCF, human neutrophils became polarized, contrasting with non-stimulated cells and similarly to cells incubated with well-characterized neutrophil attractants such as fMLP and casein (Sánchez-Madrid and Del Pozo 1999; Wagner and Roth 2000; Lindbom and Werr 2002). As observed in response to other known neutrophil attractants (Lauffenburger and Horwitz 1996), the transition from a spherical to a typically polarized shape induced by MNCF is associated with the formation of a broad, ruffled lamellipodium that is free of organelles. Large electron-dense granules were observed in the cell body, indicating the movement of secretory granules, which might reflect the reorganization of cell compartments that follows neutrophil activation after a wide variety of inflammatory stimuli (Sengelov et al. 1993a,b; Faurschou and Borregaard 2003).
The fact that
The MNCF binding pattern on the cell surface corresponds to the uniformly dispersed localization of receptors for well-known chemoattractants (Servant et al. 1999; Jin et al. 2000). The kinetics of MNCF binding and internalization in neutrophils, as reported here, is consistent with previous studies on the internalization of attractant–receptor complexes by neutrophils (Grob et al. 1990; Samanta et al. 1990; Chuntharapai and Kim 1995; Sabroe et al. 1997). Feniger-Barish et al. (1999) demonstrated that IL-8 internalization by neutrophils is initiated within 5 min of contact with the cells, and the intracellular signal is more intense at 30 and 120 min. A rapid and reversible disappearance of attractant receptors from the neutrophil surface, upon ligand binding, involves receptor clustering on the plasma membrane, receptor internalization, and recycling and accounts for the chemoattractant desensitization of neutrophils (Sabroe et al. 1997), a process that regulates neutrophil accumulation during an inflammatory response in vivo.
Receptors for well-established chemoattractants belong to the serpentine family characterized by seven-transmembrane domains. They transmit intracellular signals by receptor phosphorylation dependent G proteins (Chuang et al. 1996; Ferguson et al. 1996; Böhm et al. 1997; Koenig and Edwardson 1997), a process blocked by pertussis toxin (Becker et al. 1987; Berger et al. 2002). In our experiments, pertussis toxin drastically inhibited MNCF, as well as fMLP and IL-8 without affecting cell viability. Among lectins, this effect is not unique to MNCF, once it has been demonstrated that galectin-3 attracts mononuclear leukocytes through the activation of a specific GPCR (Sano et al. 2000).
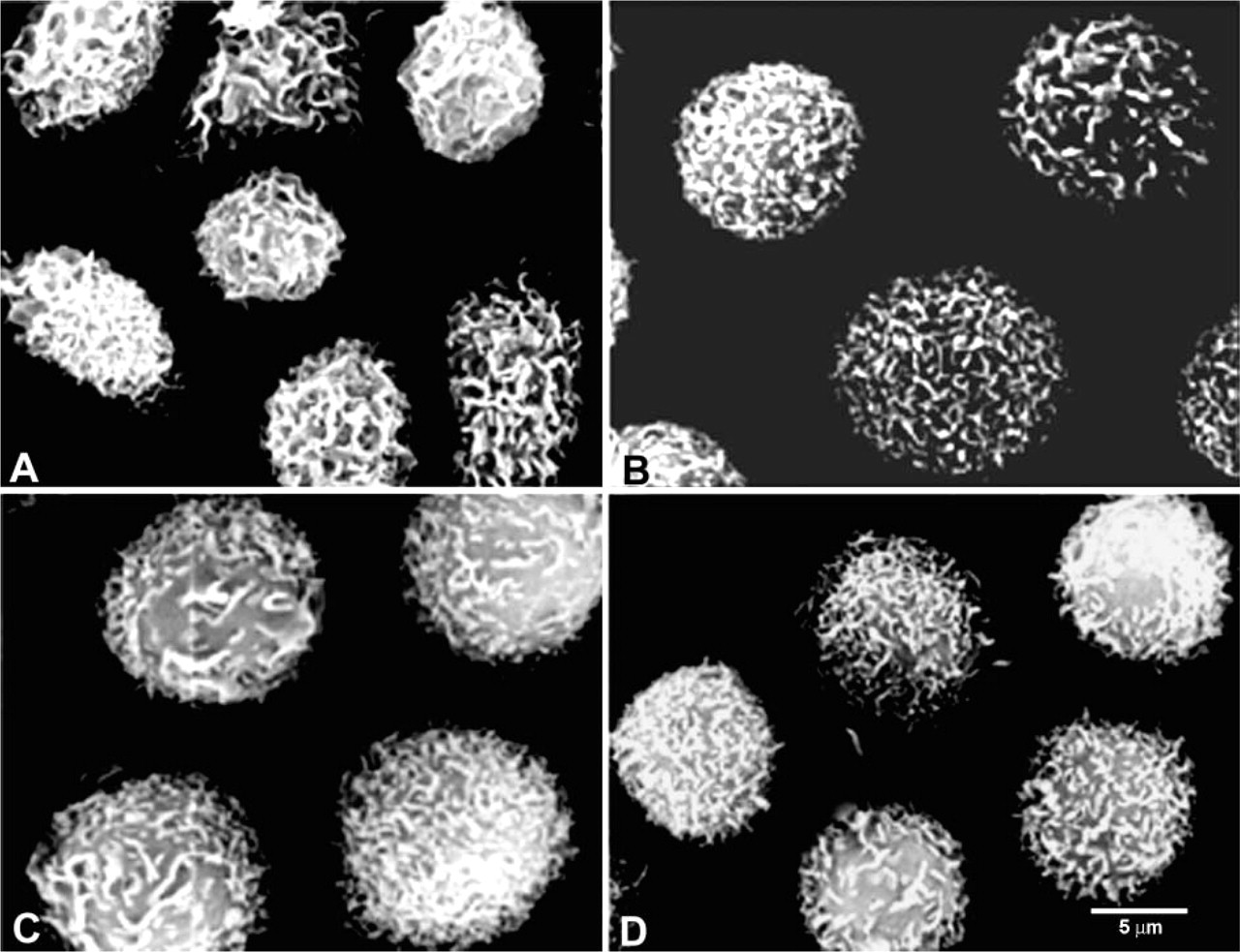
Neutrophil pretreatment with pertussis toxin inhibits morphological changes induced by MNCF. Isolated human neutrophils incubated for 30 min at 37C with MNCF (
IL-8 and other CXC chemokines activate human neutrophils through their interaction with the CXCR1 and CXCR2 receptors, which are coexpressed on the cell surface (Holmes et al. 1991; Murphy and Tiffany 1991). These receptors are seven-transmembrane domain-type proteins functionally coupled to G proteins. Because they are glycoproteins (Besemer et al. 1989; Grob et al. 1990), these receptors are potential ligands for MNCF. The CXCR2 amino acid sequence contains consensus sites for N-glycosylation at asparagine residues, located within the N-terminal part of the receptor (Asn17) and within the second extracellular loop (Asn186 and Asn197) (Murphy and Tiffany 1991). The second extracellular loop is a critical domain for agonist binding and for initiation of the signal transduction pathway (Vaidehi et al. 2002). At least two of these sites are used, each carrying an N-linked sugar moiety of 9 kDa. Indeed, two distinct CXCR-2 glycoforms were identified in neutrophils, each with a distinct subcellular distribution. The highly glycosylated variants are expressed on the cell surface, whereas a less-glycosylated form is found as an intracellular pool of CXCR-2 molecules. Full glycosylation of the receptor is required for its resistance to neutrophil proteases (Ludwig et al. 2000) but not for its interaction with IL-8. This observation goes against the general idea that N-glycans on cell-surface receptors influence their ligand binding (Paulson 1989). On the other hand, our observations strongly suggest that when agonists are lectins, a direct involvement of receptor glycans in binding and cell signaling may be required. This is true for KM+, a plant lectin that induces neutrophil migration (Santos-de-Oliveira et al. 1994) by interacting with CXCR-2 glycans (Pereira-da-Silva G et al., unpublished data). Highly glycosylated CXCR-2 forms are likely targets for MNCF recognition on the neutrophil surface and are probably able to initiate cell activation.
Relatively high concentrations of MNCF are required for the demonstration of optimal chemoattractant and cell-polarizing activities, when compared with chemokine concentrations necessary to trigger similar responses in neutrophils (Feniger-Barish et al. 1999). High local concentrations of MNCF could be attained if there is a burst of proteins released from stimulated macrophages as postulated galectin-3 (Sano et al. 2000). We observed that macrophages present strong labeling for MNCF in their cytoplasm and that the lectin is rapidly mobilized to the cell surface and to the extracellular medium following LPS stimulus (unpublished data). One hour after stimulation, the cells no longer expressed MNCF, and the lectin could be captured from the supernatant by
The resistance of the MNCF activity to glucocorticoids makes the lectin a very special inflammatory mediator and justifies the interest in its full biological and structural characterization. Until now, this task has been seriously limited by the fact that only one MNCF source has been identified, i.e., rat peritoneal macrophages requiring a large number of animals to yield a low amount of purified lectin. The obtained preparations were demonstrated to be free of chemokines. Other macrophage products able to induce neutrophil migration, such as IL-8, were recovered in a different chromatography fraction obtained from MNCF, i.e., they were not adsorbed on immobilized
In conclusion, the present study shows that MNCF exerts its neutrophil migration-inducing property by recognizing
Footnotes
Acknowledgements
Supported by FAPESP (00/09333–2), CNPq (350418/00–4), and FAEPA grants. A.N.M. received a PhD research fellowship from FAPESP (98/06948–4).
We thank Mrs. Sandra M.O. Thomaz, Mrs. Imaculada C. Bragheto, and Mrs. Patricia Vendruscolo for technical assistance, and we thank Mrs. Erica Vendruscolo for assistance in preparing the manuscript. We are indebted to Mrs. Marcia S.Z. Graeff, Mrs. Maria Tereza P. Maglia, and Mr. Jose Augusto Maulin for expert assistance with confocal and electron microscopy. We are grateful to Dr. Marcelo Dias-Baruffi for helpful discussions.