
Abstract
The epithelial barrier of the upper respiratory tract plays a crucial role in host defense. In this study, to elucidate whether there is antigen monitoring by dendritic cells (DCs) beyond the epithelial tight-junction barrier in allergic rhinitis, we investigated the expression and function of tight junctions and characterized DCs in the epithelium of nasal mucosa from patients with allergic rhinitis. In reverse transcription-polymerase chain reaction, mRNAs of tight-junction proteins occludin, JAM-1, ZO-1, and claudin-1, −4, −7, −8, −12, −13, and −14 were detected in the nasal mucosa. Occludin, JAM-1, and ZO-1 were colocalized in the uppermost layer in the pseudostratified epithelium of the nasal mucosa, whereas claudin-1, −4, and −7 were found throughout the epithelium. In freeze-fracture replicas of the nasal mucosa, continuous tight-junction strands formed well-developed networks. Epithelial barrier function measured by a dye tracer was well maintained in occludin-positive tight junctions in the epithelium of the nasal mucosa. HLA-DR- and CD11c-positive DCs expressed claudin-1 and penetrated beyond occludin in the epithelium of the nasal mucosa with, but not without, allergic rhinitis. These results indicate that DCs may easily access antigens beyond epithelial tight junctions in the human nasal mucosa of allergic rhinitis.
T
The upper respiratory tract, which is the first site of invading antigen exposure, plays a crucial role in host defense via the mucosal immune response. The epithelium of upper respiratory tissues such as nasal mucosa forms a continuous barrier against a wide variety of exogenous antigens (Herard et al. 1996; Van Kempen et al. 2000). The epithelial barrier function is regulated in large part by the apical-most intercellular junction, referred to as the tight junction (Schneeberger and Lynch 1992). Previous studies have shown that following allergic inflammation, disruption of tight junctions occurs in cultured human nasal epithelial cells (Kobayashi et al. 2000; Wan et al. 2000; Takeuchi et al. 2001). However, the distribution, expression, and function of tight-junction proteins in the epithelium of human nasal mucosa, though not those of the lower respiratory tissue such as human bronchial mucosa, are still unclear (Coyne et al. 2002,2003a,b).
Allergic rhinitis is thought to be the result of the accumulation and activation of infiltrating inflammatory cells releasing mediators and cytokines (Mygind and Dahl 1996). Antigen-presenting cells (APCs), particularly dendritic cells (DCs), seem to play an important role in allergic rhinitis (Fokkens 1999). In human nasal mucosa, different members of the DC population are observed (Fokkens 1999; Upham 2003; Jahnsen et al. 2004). The numbers of HLA-DR + DCs are increased in the epithelium of allergic rhinitis (Godthelp et al. 1996). Recently, Rescigno et al. (2001) discovered a new mechanism for pathogen uptake in the mucosa, by which DCs open the tight junctions between epithelial cells and send dendrites outside the epithelium to directly sample the pathogen. DCs express tight-junction proteins such as occludin, claudin-1, and ZO-1 to preserve the integrity of the epithelial barrier.
Thus, in the present study, we investigate the distribution, expression, and function of tight junctions in the epithelium of nasal mucosa from patients with allergic rhinitis. Furthermore, we characterize DCs, including their localization in the human nasal mucosa. We show that in the human nasal mucosa of allergic rhinitis, HLA-DR- and CD11c-positive DCs express tight-junction protein claudin-1 and penetrate beyond well-developed epithelial tight junctions.
Materials and Methods
Human Tissue Samples
Seven nasal mucosa tissues were obtained from five patients with allergic rhinitis (donors with perennial rhinitis ranging in age from 21 to 64 years) and two patients without allergic rhinitis who were undergoing inferior turbinectomy at Sapporo Medical University. The diagnosis of allergic rhinitis was established on the basis of the medical history, symptoms, nasal cytology, and specific IgE to aeroallergens (1.1–19.5 AU/ml IgE to house-dust allergens and 8.5–21.1 AU/ml IgE to mite allergens). Informed consent was obtained from all patients, and the study was approved by the Ethics Committee of Sapporo Medical University. Tissues were immediately frozen in liquid nitrogen and stored at −70C for immunohistochemistry and RNA extraction. Some tissue samples were fixed in 10% formaldehyde in PBS for hematoxylin-eosin (HE) staining.
Immunohistochemistry
For immunohistochemistry of human nasal mucosa and adenoid slices, 10-μm-thick frozen sections were made with a cryostat. The sections were fixed with cold acetone and ethanol (1:1) for 10 min. After rinsing in PBS, the sections of human nasal mucosa and adenoid were incubated with monoclonal anti-occludin, polyclonal anti-ZO-1, polyclonal anti-JAM-1, polyclonal anti-claudin-1, monoclonal anti-claudin-4, and polyclonal anti-claudin-7 antibodies at room temperature for 1 hr and then were incubated with Alexa 488 (green)-conjugated anti-mouse IgG or anti-rabbit IgG (Molecular Probes Inc.; Eugene, OR) at room temperature for 1 hr. These primary antibodies were obtained from Zymed Laboratories (San Francisco, CA). Some sections were used for double staining of occludin and claudin-1, occludin and JAM-1, occludin and ZO-1, occludin and claudin-4, occludin and claudin-7, claudin-1 and claudin-4, or claudin-1 and LCA (DAKO; Carpinteria, CA), which were visualized by Alexa 488 (green)-conjugated anti-rabbit IgG and Alexa 594 (red)-conjugated anti-mouse IgG (Molecular Probes). Some sections from human nasal mucosa were incubated with monoclonal anti-CD8, monoclonal anti-CD11b, monoclonal anti-CD11c, monoclonal anti-CD80, monoclonal anti-CD86, and monoclonal anti-human HLA-DR antibodies. These first antibodies were obtained from DAKO. Some sections were used for double staining of occludin and HLA-DR, occludin and CD11c, claudin-1 and HLA-DR, claudin-1 and CD11c, which were visualized by Alexa 488 (green)–conjugated anti-rabbit IgG and Alexa 594 (red)–conjugated anti-mouse IgG (Molecular Probes). The specimens were examined using an epifluorescence microscope (Olympus; Tokyo, Japan) and a laser-scanning confocal microscope (MRC 1024; Bio-Rad, Hercules, CA).
RNA Isolation and Reverse Transcription-Polymerase Chain Reaction (RT-PCR) Analysis
Total RNA was extracted from the nasal mucosa after homogenization in TRIzol reagent (Gibco BRL; Gaithersburg, MD). For RT-PCR, 1 μg of total RNA was reverse transcribed into cDNA using the manufacturer's recommended conditions (Invitrogen; Carlsbad, CA). Each cDNA synthesis was performed in a total volume of 20 μl for 50 min at 42C and terminated by incubation for 15 min at 70C. Subsequently, RT-PCR was performed using 2 μl of the 20-μl total RT product, PCR buffer, dNTPs, and Premix Taq DNA polymerase under the manufacturer's recommended conditions (Takara Inc.; Siga, Japan). Conditions applied for PCR were 96C for 30 sec, 30 cycles of 96C for 15 sec, 55C for 30 sec, 72C for 1 min, and 72C for 7 min using a PerkinElmer/Cetus Thermocycler Model 2400 (PerkinElmer Inc.; Branchburg, NJ). Ten μl of the 20-μl total PCR reaction was analyzed by electrophoresis in ethidium bromide–impregnated 1% agarose gels. Primers used to detect occludin, ZO-1, JAM-1, and claudin-1, −2, −3, −4, −5, −6, −7, −8, −9, −12, −13, and −14 had the following sequences: occludin (sense 5'-TCAGGGAATATCCACCTATCACTTCAG-3’ and anti-sense 5'-CATCAGCAGCAGCCATGTACTCTTCAC-3', amplicon length: 136 bp), ZO-1 (sense 5'-CGGTCCTCTGAGCCTGTAAG-3’ and antisense 5'-GGATCTACATGCGACGACAA-3', amplicon length: 435 bp), JAM-1 (sense 5'-GGTCAAGGTCAAGCTCAT-3’ and antisense 5'-CTGAGTAAGGCAAATGCAG-3', amplicon length: 765 bp), claudin-1 (sense 5'-GCTGCTGGGTTTCATCCTG-3’ and antisense 5'-CACATAGTCTTTCCCACTAGAAG-3', amplicon length: 619 bp), claudin-2 (sense 5'-GCAAACAGGCTCCGAAGATACT-3’ and antisense 5'-CTCTGTACTTGGGCATCATCTC-3'), claudin-3 (sense 5'-TGCTGTTCCTTCTCGCCGCC-3’ and antisense 5'-CTTAGACGAAGTCCATGCGG-3', amplicon length: 247 bp), claudin-4 (sense 5'-AGCCTTCCAGGTCCTCAACT-3’ and antisense 5'-AGCAGCGAGTAGAAG-3', amplicon length: 249 bp), claudin-5 (sense 5'-GACTCGGTGCTGGCTCTGAG-3’ and antisense 5'-CGTAGTTCTTCTTGTCGTAG-3'), claudin-6 (sense 5'-TGAGGCCCAAAAGCGGGAGC-3’ and antisense 5'-CGTAATTCTTGGTAGGGTAC-3'), claudin-7 (sense 5'-AGGCATAATTTTCATCGTGG-3’ and antisense 5'-GAGTTGGACTTAGGGTAAGAGCG-3', amplicon length: 210 bp), claudin-8 (sense 5'-TCATCCCTGTGAGCTGGGTT-3’ and antisense 5'-TGGAGTAGACGCTCGGTGAC-3', amplicon length: 215 bp), claudin-9 (sense 5'-AGGCCCGTATCGTGCTCACC-3’ and antisense 5'-ACGTAGTCCCTCTTGTCCAG-3'), claudin-12 (sense 5'-CTCCCCATCTATCTGGGTCA-3’ and antisense 5'-GGTGGATGGGAGTACAATGG-3', amplicon length: 201 bp), claudin-13 (sense 5'-TGGTCGTCAGCAAACAAGAG-3’ and antisense 5'-GCCAAGCAATGGGTTAAAGA-3', amplicon length: 455 bp), claudin-14 (sense 5'-GCACCTAAGGACCAGATCCA-3’ and antisense 5'-CATCCACAGTCCCTTCAGGT-3', amplicon length: 627 bp). To provide a qualitative control for reaction efficiency, PCR reactions were performed with primers coding for the housekeeping gene G3PDH (sense 5'-ACCACAGTCCATGCCATCAC-3’ and antisense 5'-TCCACCACCCTGTTGCTGTA-3', amplicon length: 452 bp).
Freeze-fracture Analysis
For a freeze-fracture experiment, the nasal mucosa tissues were immersed in 40% glycerin solution after fixation in 2.5% glutaraldehyde in 0.1 M PBS (pH 7.3). The specimens were mounted on a copper stage, frozen in liquid nitrogen, fractured at −150C to −160C, replicated by platinum/carbon from an electron beam gun positioned at a 45° angle followed by carbon applied from overhead in a JFD-7000 freeze-fracture device (JEOL; Tokyo, Japan). After the replicas were thawed, they were floated on filtered 10% sodium hypochlorite solution for 10 min in a Teflon dish. Replicas were washed in distilled water for 30 min, mounted on copper grids, and examined at 100 KV on a JEOL 1200EX transmission electron microscope.
Barrier Function Assay Using Dye Tracer
To evaluate the barrier function of tight junctions, the nasal mucosa tissues were incubated with rhodamine–dextran (molecular mass 1000 Da; Molecular Probes) dissolved in PBS for 15 min on ice. The specimens were immediately frozen in liquid nitrogen, sectioned 10 μm thick with a cryostat, and mounted on glass slides. Some sections were double stained with occludin or claudin-1 visualized by Alexa 488 (green)–conjugated anti-mouse IgG. The specimens were examined using an epifluorescence microscope (Olympus).
Results
Distribution of Tight-junction Proteins in the Epithelium of Nasal Mucosa from Patients with Allergic Rhinitis
In HE staining of nasal mucosa from patients with allergic rhinitis, basement membrane thickening was observed, and the surface epithelium of the mucosa and the epithelium of mucosal glands were pseudostratified (Figure 1A). However, the epithelial damage of the nasal mucosa was minimal (Figure 1A). To investigate the distribution of tight-junction proteins in the epithelium of the nasal mucosa from patients with allergic rhinitis, double immunohistochemistry was carried out. In the epithelium of the nasal mucosa and nasal gland, occludin was found in the uppermost layers (Figure 1A). Claudin-1 was observed throughout the epithelium and was colocalized with occludin in the uppermost layers (Figure 1A). Claudin-1 was highly expressed in the basal layers (Figure 1A). LCA-positive lymphocytes were observed in the claudin-1-positive pseudostratified epithelium of the nasal mucosa and nasal gland (Figure 1A). Furthermore, JAM-1 and ZO-1 were colocalized with occludin in the uppermost layers of the epithelium (Figure 1B). Claudin-4 and −7 were observed throughout the epithelium and colocalized with occludin in the uppermost layers (Figure 1B).
Expression in mRNAs of Tight Junctions in Human Nasal Mucosa
To investigate the expression in mRNA of tight junctions, occludin, JAM-1, ZO-1, claudins 1–9, and claudins 12–14 in nasal mucosa from patients with allergic rhinitis, RT-PCR was carried out. As shown in Figure 2A, expression of mRNAs of occludin, JAM-1, ZO-1, and claudin-1, −4, −7, −8, −12, −13, and −14 was detected in human nasal mucosa.
Freeze-fracture Analysis of Human Nasal Mucosa
To investigate whether tight-junction structures in the nasal mucosa expressed several claudins that can form tight-junction strands, we performed freeze-fracture analysis. In the freeze-fracture replicas, continuous tight-junction strands that formed well-developed networks were observed on the subapical membranes in the nasal mucosa (Figure 2B).
Barrier Function Assay of Tight Junctions in Human Nasal Mucosa
To evaluate the barrier function of tight junctions in the nasal mucosa, dye tracing using rhodamine-dextran was carried out. The dye remained on the uppermost layer of the epithelium of the nasal mucosa and leaked from the surgical surface to inner tissue (Figure 2C). In the double staining for occludin and rhodamine-dextran or claudin-1 and rhodamine-dextran of the epithelium of the nasal mucosa, the dye was sharply stopped at occludin-positive, but not claudin-1-positive, tight junctions (Figure 2C: c,d).
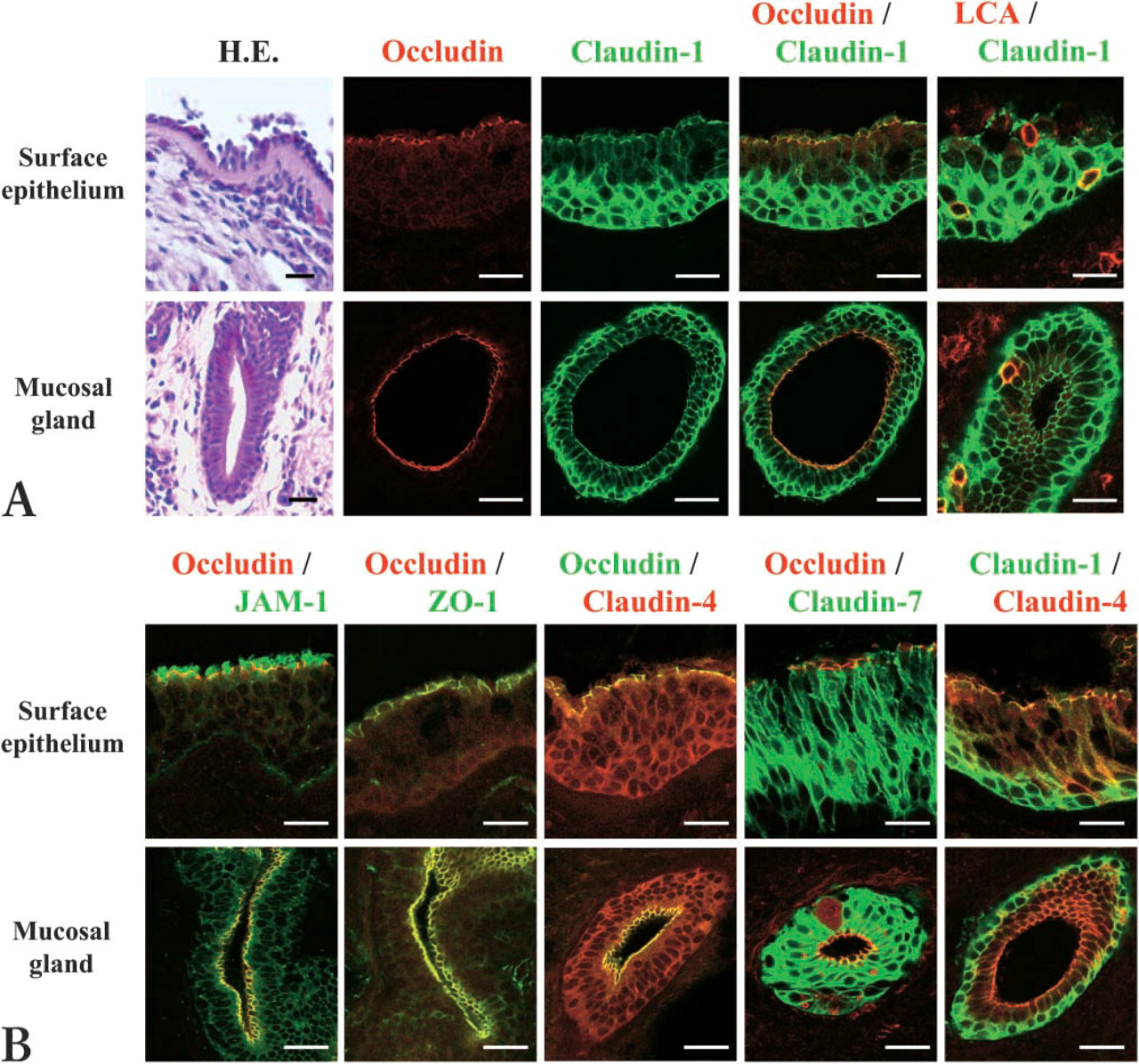
Hematoxylin-eosin staining (
Localization and Tight-junction Protein Expression of DCs in the Epithelium of Human Nasal Mucosa
It is thought that DCs open the tight junctions between epithelial cells and send dendrites outside the epithelium, expressing tight-junction proteins in the gastrointestinal mucosa (Rescigno et al. 2001). However, in the nasal mucosa, the interaction between DCs and tight junctions is still unclear. We examined phenotypes of DCs in the epithelium of nasal mucosa from patients with allergic rhinitis and localization of DCs in the epithelium. We used several antibodies (see Materials and Methods) for DC antigens and performed double immunohistochemistry for occludin and HLA-DR, occludin and CD11c, claudin-1 and HLA-DR, or claudin-1 and CD11c. CD8-, CD11c-, and HLA-DR-positive DCs were detected in the epithelium and submucosal layers of nasal mucosa from patients with allergic rhinitis (Figures 3A, 3D, 3G, and 3J) and were found in the claudin-1-positive epithelium (Figures 3B, 3E, 3H, and 3K). Dendritic cytoplasm of HLA-DR- and CD11c-positive DCs was observed near or beyond occludin, which indicated the uppermost layers of the epithelium (Figures 3C and 3F, arrows). HLA-DR- and CD11c-positive DCs expressed claudin-1 in the epithelium (Figures 3I and 3L). On the other hand, in human nasal mucosa without allergic rhinitis, there were few CD11c- or HLA-DR-positive DCs in the epithelium, and they never penetrated beyond occludin (data not shown).
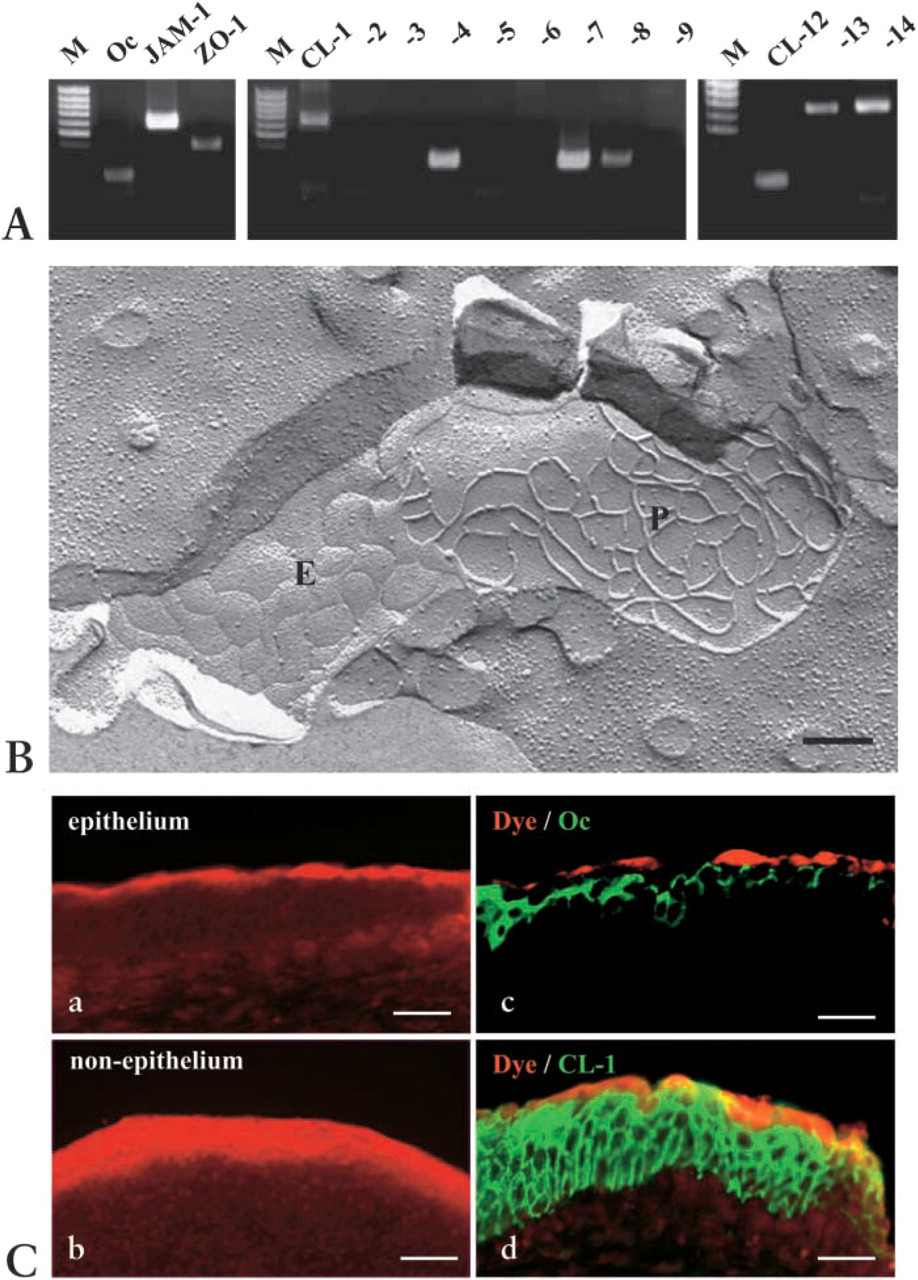
RT-PCR in mRNAs of tight junctions in human nasal mucosa from patients with allergic rhinitis. M, 100-bp ladder DNA marker. Expression of mRNAs of occludin, JAM-1, ZO-1, and claudin-1, −4, −7, −8, −12, −13, and −14 was detected in the human nasal mucosa. (
Discussion
In this study, the epithelial damage of the nasal mucosa of patients with allergic rhinitis was minimal, and the protein expression, structures, and function of the epithelial tight junctions were well maintained. In the human nasal mucosa of allergic rhinitis, HLA-DR- and CD11c-positive DCs expressed tight-junction protein claudin-1 and penetrated beyond the apical-most tight-junction protein occludin to minimize the increase in permeability of the epithelial barrier.
The tight junction is a complex structure composed of several protein components. The claudins comprise a multigene family, with 24 members identified to date, and the expression pattern of claudins is tissue and segment specific (Turksen and Troy 2004). In pseudostratified columnar, ciliated epithelium of the human bronchial mucosa, such as the lower respiratory tissue, claudin-3 and −5 were localized to the apical-most region, whereas claudin-1, −4, and −7 were detected throughout the lateral intercellular junctions (Coyne et al. 2002). In primary cultured bronchial cells, occludin and ZO-1 were localized to the apical-most region and JAM-1 was detected throughout the lateral intercellular junction (Coyne et al. 2003a). In the freeze-fracture replicas of the primary cultured bronchial cells, continuous tight-junction strands formed well-developed networks and the epithelial barrier function measured by a dye tracer was maintained (Coyne et al. 2003a,b). In this study, in the human nasal mucosa, expression of mRNAs of occludin, JAM-1, ZO-1, and claudin-1, −4, −7, −8, −12, −13, and −14 was detected. In the pseudostratified columnar epithelium of the nasal mucosa, occludin, JAM-1, and ZO-1 were found in the uppermost layers and claudin-1, −4, and −7 were detected throughout the epithelium. In the freeze-fracture replicas of the nasal mucosa, continuous tight-junction strands formed well-developed networks. Epithelial barrier function measured by a dye tracer was maintained in the nasal mucosa of allergic rhinitis. Because the nose and bronchi have a different embryologic origin (Bousquet et al. 2004), the epithelium of nasal mucosa and bronchial mucosa presented, in part, differences in expression and localization of tight-junction proteins but both had well-developed tight junctions. Furthermore, expression and function of the epithelial tight junctions in the nasal mucosa were also different from those of the oral mucosa when we previously examined the epithelial tight junctions of human palatine tonsils such as the oral mucosa (Go et al. 2004). In the stratified epithelium of human palatine tonsils, mRNAs of occludin, ZO-1, JAM-1, and claudin-1, −3, −4, −7, −8, and −14 were detected. Occludin and claudin-14 were expressed in the uppermost layer of the palatine tonsillar surface epithelium, whereas ZO-1, JAM-1, and claudin-1, −4, and −7 were found throughout the epithelium. In the crypt epithelium, claudin-4 was preferentially expressed in the upper layers. However, in the freeze-fracture replicas of the palatine tonsils, short fragments of continuous tight-junction strands were observed, but they did not form networks.
Occludin is known to be highly concentrated at tight-junction strands in the apical-most epithelial cells and is the best general marker for such strands, but is not indispensable for their formation (Tsukita and Furuse 2002). In mouse epidermis, the diffusion of a subcutaneously injected tracer was sharply stopped at occludin-positive tight junctions (Furuse et al. 2002). In this study, the dye tracer was also sharply stopped at occludin-positive, but not claudin-1-positive, tight junctions in the epithelium of the nasal mucosa. This result suggests that occludin-positive tight junctions are also closely related to epithelial barrier function in human nasal mucosa.
In human nasal mucosa, different members of the DC population are observed and the DCs seem to play an important role in allergic rhinitis (Fokkens 1999; Upham 2003; Jahnsen et al. 2004). In this disease, the number of respiratory mucosal DCs is increased (Godthelp et al. 1996), and these DCs express a high-affinity IgE receptor (Tunon-De-Lara et al. 1996). Recently, Rescigno et al. (2001) discovered a route by which some pathogenic microorganisms could invade through epithelial cells via intraepithelial DCs, because the DCs between the epithelial cells of the intestine are reported to express tight-junction proteins such as occludin, claudin-1, and ZO-1. Furthermore, Der p 1, which is a cysteine proteinase allergen, causes disruption of tight-junction barrier and may move via the paracellular pathway to interact with APCs, including DCs, in lung epithelium (Wan et al. 1999). In this study, to elucidate whether there is a similar mechanism between DCs and tight junctions in the nasal mucosa of allergic rhinitis, we characterized DCs in the human nasal mucosa of allergic rhinitis and examined the localization of DCs and tight-junction proteins in the epithelium using immunohistochemistry. In the epithelium and submucosal layers of nasal mucosa of allergic rhinitis, CD8-positive, CD11c-positive, and HLA-DR-positive DCs were detected. In double immunostaining, HLA-DR- and CD11c-positive DCs expressed tight-junction protein claudin-1 and penetrated beyond tight-junction protein occludin in the epithelium of the nasal mucosa of allergic rhinitis. In the nasal mucosa without allergic rhinitis, there were few CD11c- and HLA-DR-positive DCs in the epithelium, and they did not penetrate beyond occludin-positive tight junctions (data not shown). These results suggest that DCs may uptake antigens beyond the well-developed epithelial barrier in human nasal mucosa of allergic rhinitis. In this study, however, the numbers and localization of DCs were not associated with the levels of allergic rhinitis.
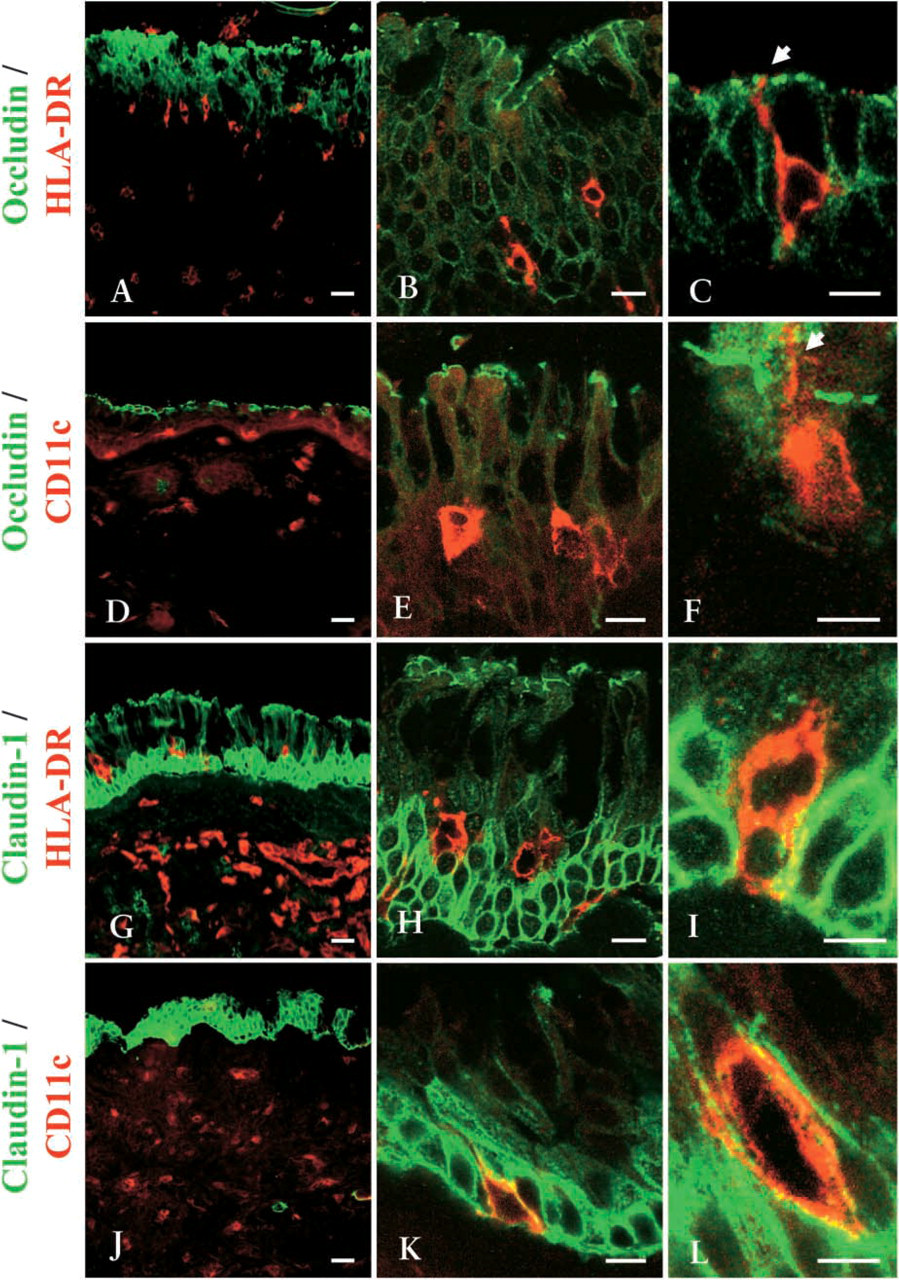
Double-immunohistochemical staining for occludin and HLA-DR (
In conclusion, we have demonstrated for the first time that in human nasal mucosa of allergic rhinitis in situ, HLA-DR- and CD11c-positive DCs express tight-junction proteins and penetrate beyond well-developed epithelial tight junctions. In the claudin family, claudin-3 and −4 are known to be receptors for enterotoxin of Clostridium perfringens and are overexpressed in some solid tumors (Katahira et al. 1997; Fujita et al. 2000). Furthermore, the tight junction–associated proteins JAM and CAR have been suggested to be receptors for reovirus or coxsackievirus and adenovirus (Barton et al. 2001; Cohen et al. 2001). Further studies are necessary to elucidate the specific roles of the tight-junction molecules in innate immunity in human nasal mucosa as well as in allergic reaction.
Footnotes
Acknowledgements
This work was supported by grants-in-aid from the Ministry of Education, Culture, Sports, and Science and Technology; by the Ministry of Health, Labor, and Welfare of Japan; and by the Long-range Research Initiative Project of the Japan Chemical Industry Association.
We are grateful to Dr. K. Asano, Dr. M. Hirao, and Dr. M. Kurose for the performance of inferior turbinectomy, and to Dr. T. Kita (Kyoto University) for the JAM-1 antibody. We thank Ms. E. Suzuki (Sapporo Medical University) for technical support.