
Abstract
Inactivation of the cell cycle inhibitor gene p16MTS1 seems to be involved in human papillomavirus (HPV)-related carcinogenesis because E6 and E7 oncoproteins may impair p16INK4a and, indirectly, bcl-2 functions. In this study, we analyzed the role of immunohistochemical expression of p16INK4a and bcl-2 in HPV-infected cervical biopsies as prognostic markers of the progression of squamous intraepithelial lesion (SIL). Sixty-five cervical biopsies were stratified into two subgroups according to the second biopsy: 27 of them maintained a low-grade (LG)-SIL diagnosis, and 38 progressed from LG-SIL to high-grade (HG)-SIL. p16INK4a and bcl-2 quantitative expression levels were measured by the immunoperoxidase method. PCR-DNA techniques were used to detect and type HPV. The Wilcoxon and Fisher exact tests were employed for the statistical analysis. In the group with an LG-SIL diagnosis at the second biopsy, no significant associations were found between p16INK4a and bcl-2 expression and presence of HPV16/18. In the group that progressed to HG-SIL, a significant association was observed between p16INK4a overexpression and HPV16/18 presence (p=0.021), but none with bcl-2 levels. It is concluded that immunohistochemical bcl-2 expression may not be useful for predicting the progression of HPV-related SIL. In contrast, p16INK4a overexpression seemed to be associated with HPV 16 and 18, suggesting that it may be a good marker for predicting SIL progression.
C
Although more than 85 types of HPV have been detected in the genital mucosa (Chan et al. 1995), in the majority of HPV-infected individuals, the virus is eliminated. However, occasionally, lesions do not regress and malignant progression may proceed under appropriate widespread metastasis (Canavan and Doshi 2000).
Following this reasoning, experimental research currently aimed at identifying biomarkers for the prognosis of squamous intraepithelial lesions (SILs) indicates the potential significance of p16INK4a and bcl-2 proteins (Ter Harmsel et al. 1996; Van de Putte et al. 2003).
p16INK4a is a tumor suppressor protein that inhibits the function of cdK4 and cdK6, which in turn regulate the G1 checkpoint. CDK/cyclin-D phosphorylate the retinoblastoma protein (pRb), resulting in a conformational change, with the release of E2F from pRb. Thus, inactivation of either p16INK4a or Rb function allows the cell to enter the S phase after only a brief pause at the G1 checkpoint. In addition, the E6 HPV oncoprotein has the ability to bind to p53, resulting in its degradation, and the E7 gene product inactivates the pRb pathway (Alani and Munger 1998).
Another possible mechanism that contributes to malignant cell transformation involves the deregulation of the apoptotic pathway. One among several components that control programmed cell death, bcl-2 is localized in the inner mitochondrial membrane and is responsible for the prevention of apoptotic cell death in several situations. Inappropriate expression of bcl-2 may prolong survival of defective and harmful cells, including those involved in HPV infection, thus increasing the probability of malignant conversion (Hockenbery et al. 1990; Green et al. 1994). Furthermore, overexpression of bcl-2 can block p53-mediated G1 arrest, and coexpression of c-myc and bcl-2 can inhibit p53-induced apoptosis (Ryan et al. 1994). However, the interference of HPV with bcl-2 expression leading to cancer progression is not clear, and further studies are needed to clarify the role of bcl-2 as an oncogenic marker.
In the present study, cervical biopsies presenting with low-grade (LG)-SIL, infected with high-risk HPV types (16 and 18), were assessed using immunohistochemical markers (p16INK4a and bcl-2) to verify their prognostic potential for progression toward high-grade (HG)-SIL.
Materials and Methods
Tissue Specimens
Sixty-five cervical biopsies were obtained from 31 patients colposcopically and were cytologically screened for HPV and SILs. All patients presented LG-SIL in a first biopsy (n= 31). They were followed for 6 months to 1 year (mean, 9 months), and then stratified according to the second biopsy diagnosis: group 1, which maintained LG-SIL (n= 13); and group 2, which progressed to HG-SIL (n= 18). Three slides presenting excessive digestion after antigen retrieval in the immunohistochemical procedure were not available for microscopic evaluation and thus, they were considered to be unsatisfactory and were excluded from the study. Formalin-fixed and paraffin-embedded cervical biopsies were selected retrospectively from the archives of the Pathology Department (from 1992 to 2002), School of Medicine of Ribeirão Preto, University of São Paulo, Brazil, and the study protocol was approved by the institutional ethics committee on human experimentation. Slides were stained with hematoxylin-eosin and histologically evaluated by two experienced histopathologists in a double-blind protocol. Thin (5-μm) sections were cut, placed on organosilane-pretreated slides, and submitted to immunohistochemical assays (p16INK4a and bcl-2). An additional 10-μm section was used for DNA extraction and HPV typing.
Immunohistochemical Staining Procedures
p16INK4a . For the detection of p16INK4a protein, immunohistochemistry was performed using the streptavidin-biotin system (LSAB; DAKO, Glostrup, Denmark). The sections were dewaxed in xylene, rehydrated in graded alcohol, rinsed in water, and treated by five immersions in a 0.3% hydrogen peroxide bath in absolute methanol (five minutes each change) and rinsing in water to inhibit endogenous peroxidase activity. For antigen retrieval, the sections were immersed in 10 mM sodium citrate buffer, pH 6.0, and boiled twice for 12 min in a high-intensity microwave oven. The slides were incubated with a primary monoclonal antibody for p16INK4a (p16/Abs4, diluted 1:50; Labvision, Freemont, CA) in a humidified chamber at 4C overnight and then incubated with biotinylated anti-rabbit antibody and the streptavidin-peroxidase complex at 37C for 30 min. Careful rinses were performed, with several changes of PBS between each stage of the procedure. The sections were incubated with DAB (Gibco; Gaithersburg, MD), lightly counterstained with Carrazzi's hematoxylin without acid for 60 sec, exhaustively rewashed with tap water, air dried, and mounted with Permount mounting medium (MERCK; Darmstadt, Germany). Positive controls consisting of HeLa cell block sections, and negative controls consisting of HeLa cell block slides, omitting the primary antibody, were included in each assay The HeLa cell line was kindly provided by Dr Sandro Roberto Valentini, School of Pharmaceutical Sciences of Araraquara, University of São Paulo State, Brazil.
Quantitative Evaluation of p16INK4a Staining. The immunohistochemical expression of p16INK4a was quantified in a double-blind protocol and classified according to nuclear and cytoplasmic positivity. The biopsies were scored according to the following stratification, adapted from Klaes et al. (2001): negative (<1% of nucleus and cytoplasm positivity); sporadic (1% to 10% of nuclei and cytoplasm with weak and scattered positivity); focal (10% to 30% labeled nuclei and cytoplasm with strong positivity, spreading in one tissue area); and diffuse (>30% to 87% of labeled cells with strong positivity spreading in several tissue areas). Biopsies that exhibited a diffuse pattern were considered to have immunohistochemical overexpression of p16INK4a. Focal distribution was interpreted as moderate expression and sporadic positivity as low expression, as previously described by Fregonesi et al. (2003).
bcl-2 Protein. The sections were dewaxed in toluene, rehydrated in graded alcohol, and rinsed in water. For antigen retrieval, the sections were immersed in 10 mM sodium citrate buffer, pH 6.0, and boiled in a water bath (95-99C) for 30 min. To prevent cross-reaction with endogenous peroxidase, the slides were treated with 3% H2O2 for 20 min and then incubated overnight at room temperature with bcl-2 antibody (mouse monoclonal antibody, clone 124; DAKO) diluted 1:200. Sections of human tonsils were used as positive controls, and a negative control to detect background staining was performed by omitting the primary antibody.
After incubation with the primary antibody, immunoperoxidase staining was performed using a universal biotinylated secondary antibody mixed with a preformed avidin and biotinylated horseradish peroxidase macromolecular complex (Novostain Super ABC Kit; NovoCastra, Newcastle upon Tyne, UK), diluted 1:200, for 1 hr. Slides were alternately washed three times with PBS and TBST-EDTA (Tris-buffered saline; 0.05 M Tris, pH 9.5). Diaminobenzidine (0.5 mg/8 ml; Sigma, St. Louis, MO) was used as chromogen. Slides were then sequentially counterstained with Harris' haematoxylin, without acid, for 40 sec, stained blue with 0.05% ammoniacal water (20 sec) and exhaustively rewashed with tap water. Finally, slides were dehydrated and mounted.
A minimum of 10 fields (total area equivalent to 0.75 mm2) for light microscopy evaluation at 400X magnification were assessed per case. The expression of bcl-2 antigens was evaluated using a previously described semiquantitative method (Coleman and Stanley 1994). A total score representing the sum of the intensity and distribution of staining in fields presenting dysplastic epithelium and in the respective subepithelial stroma was attributed to each case. Staining intensity (epithelia and stroma) was scored as follows: 0, no staining; 1, weak staining; 2, moderate staining; and 3, intense staining.
HPV Detection and Typing. HPV DNA obtained from paraffin blocks (Frank et al. 1996) was amplified by PCR using 12.5 pM of dNTP, 25 pM of each primer, 1.5 U Taq DNA polymerase (Gibco), 5 μl of 10X enzyme buffer, 20 μg of genomic DNA (50 ng), and distilled deionized H2O to complete a total volume of 25 μl. The mixture was processed in a thermocycler apparatus (MJ Research, Waltham, MA) under the following cycling conditions: 1 cycle at 95C for 5 min; 30 cycles at 95C for 30 sec, at 55C for 30 sec, and at 72C for 1 min; and finally, 1 cycle at 72C for 10 min, and then at 4C indefinitely.
Because formalin may degrade DNA, producing DNA fragments of different lengths, several pairs of primers were used. Primers GP5+ and GP6+ (Ting and Manos 1990), which amplify small DNA fragments, and primers MY09 and MY11 (Snijders et al. 1990), which amplify longer DNA fragments, were used for generic HPV amplification. Because HPV 16 and HPV 18 are the types most frequently associated with SIL and invasive cervical neoplasia, attention was focused on them. Primers HPV16E7.667/HPV16E7.774, specific for HP16 and HPV18E7.696/HPV18E7.799 (Walboomers et al. 1999), specific for HPV 18 were also used. All DNAs were amplified with these three sets of primers, together with a set of primers for a housekeeping gene (globin) (Saiki et al. 1988) as an internal control of amplification. Amplified DNA was applied to a 10% polyacrylamide gel, electrophoresed at 200 V for 1.45 hr, and stained with AgNO3 by the method of Sanguinetti et al. (1994).
It is important to note that the absence of HPV 16 or HPV 18 did not exclude the presence of HPV types other than 16 or 18, which could be identified if specific primers for them were used.
Statistical Analysis
The Mann-Whitney and Wilcoxon nonparametric tests were applied to compare the LG- and HG-SIL groups and quantitative immuhistochemical expression (p16INK4a and bcl-2). Fisher's exact test was used for the comparisons between the LG-SIL and HG-SIL groups for p16INK4a, bcl-2 overexpression, and HPV types. P values were two-sided, and the level of significance was set at ≤0.05. All data were analyzed using the Instat Mac 2.01 software (GraphPad, San Diego, CA).
Results
HPV Detection and Typing
All samples analyzed were positive for the presence of HPV. In the group in which the first biopsies were diagnosed as LG-SIL, HPV 16 was identified in 12/31 cases (38.7%), HPV 18 in 21/31 cases (67.7%), and both infections (HPV 16 and 18) were identified in 8/31 cases (25.8%). HPV typing according to the histological diagnosis at the second biopsy is presented in Table 1.
In the group that maintained the diagnosis of LG-SIL at the second biopsy, no significant associations were found between the presence of HPV 16 infection alone and the presence of both HPV 16 and 18 infections (p= 0.5804 and p= 1.00, respectively) when the second group of biopsies presenting LG-SIL was compared with the first group of biopsies.
In the group that progressed from LG-SIL to HG-SIL, no significant associations were observed between the presence of HPV 16 and the presence of both HPV 16 and 18 infections (both p= 1.00) when the second group presenting biopsies with HG-SIL was compared with the first group of biopsies.
Although HPV 18 was the most frequent HPV type observed in both groups, HPV 16 alone and associated with HPV 18 was found at higher frequency in the group that progressed from LG-SIL to HG-SIL than in the group that maintained LG-SIL in the second biopsy.
p16INK4a Immunohistochemistry
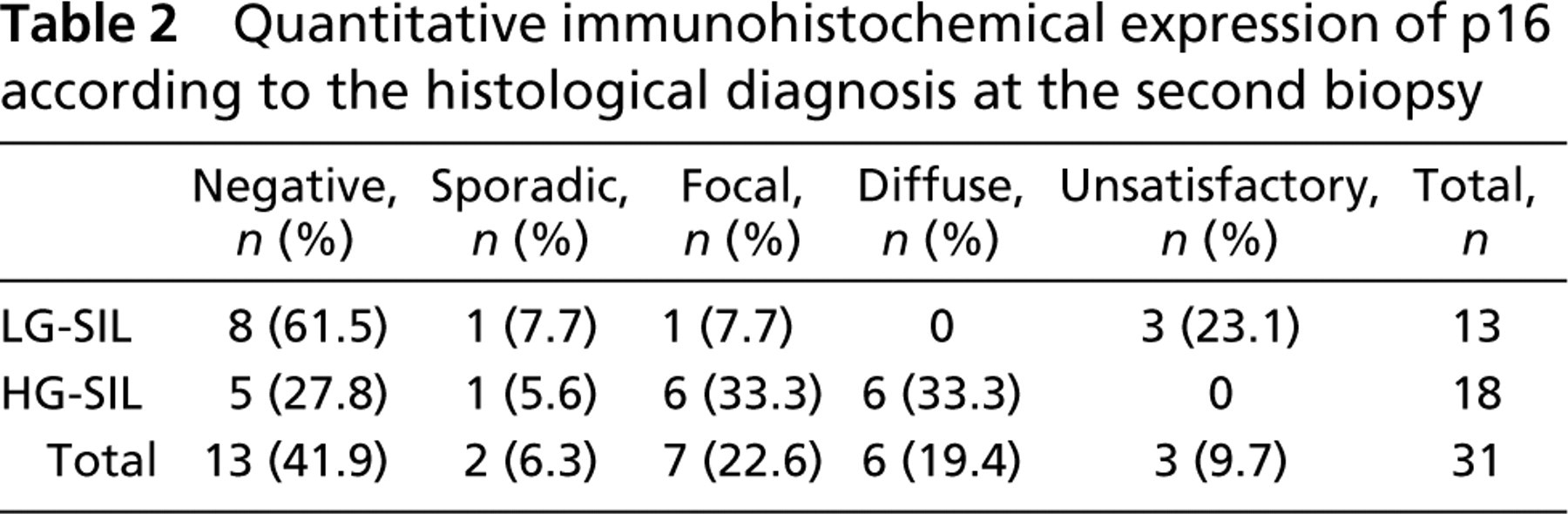
In the first biopsies with LG-SIL, p16INK4a staining distribution was negative in 11/31 cases (35.5%), sporadic in 1/31 (3.2%), focal in 7/31 (22.6%), and diffuse in 7/31 (22.6%). Five of 31 (16.1%) samples were considered unsatisfactory for this study.
Epithelial cells exhibited p16INK4a immunopositivity, in contrast to glandular cells, which did not stain. In the second biopsies with SIL, p16INK4a expression was significantly distributed according to lesion grade, i.e., LG-SIL predominantly presented sporadic staining. In HG-SIL, diffuse p16INK4a staining was mainly distributed from the basal layer up to the epithelial surface (Figures 1D, 1E, and 1F).
HPV typing according to the histological diagnosis at the second biopsy
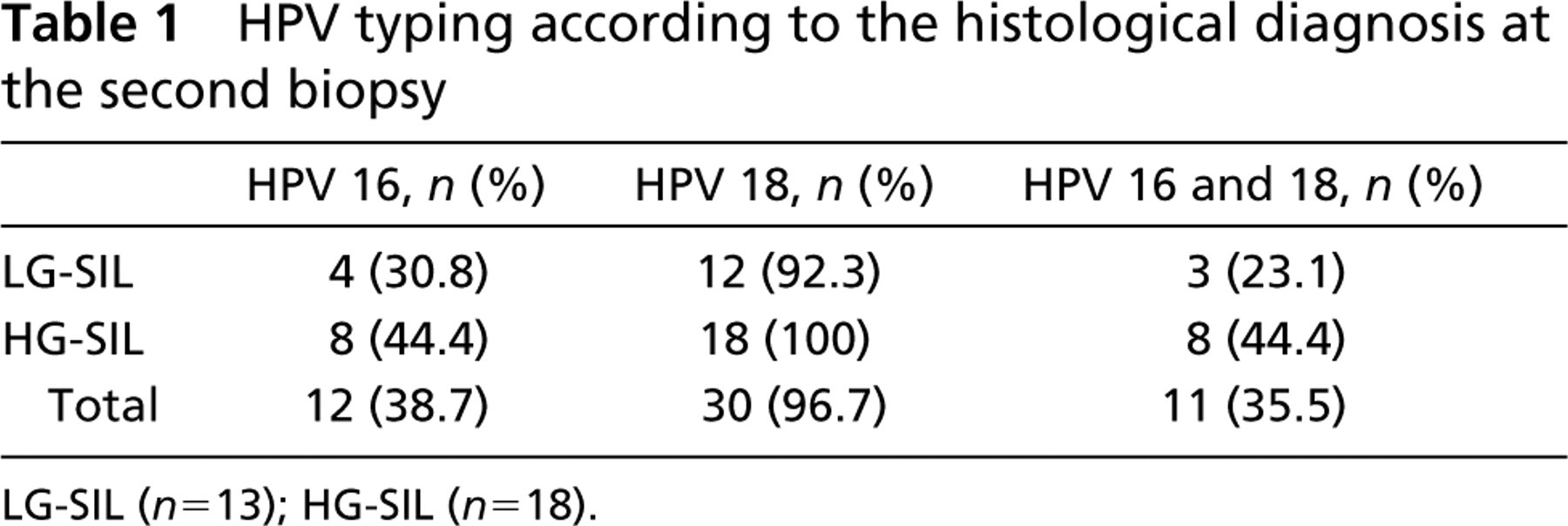
LG-SIL (n= 13), HG-SIL (n= 18).
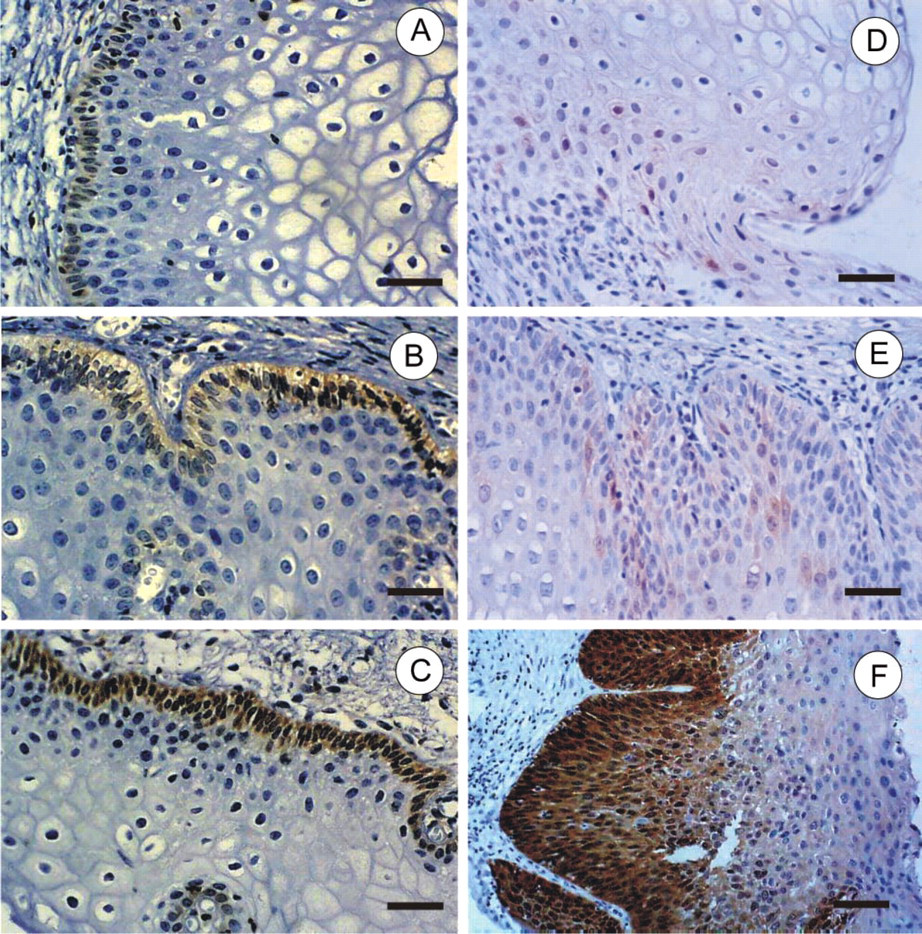
Immunohistochemical reaction for bcl-2 and p16.
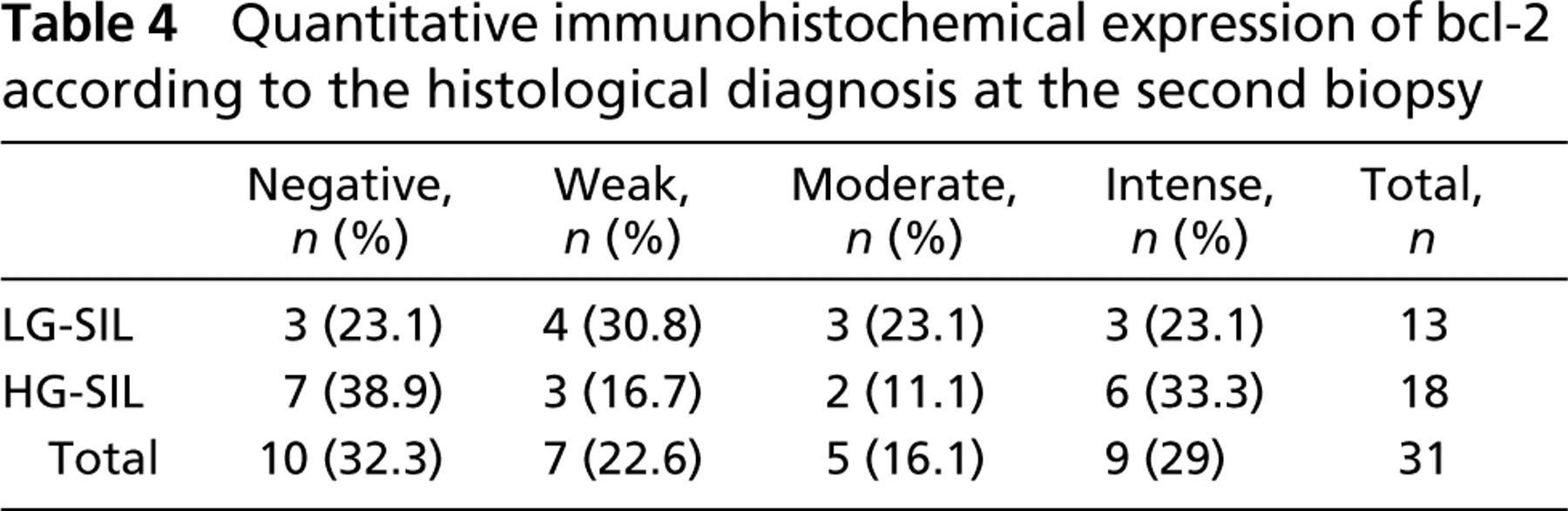
Quantitative immunohistochemical p16INK4a expression according to histological diagnosis at the second biopsy is presented in Table 2. In slides presenting with LG-SIL, p16INK4a negative expression was observed in most cases (61.5%). In contrast, in slides with HG-SIL, p16INK4a presented predominantly focal and diffuse expression, observed in 66.7% of the cases. A significant p16INK4a overexpression was observed when the group that progressed from LG- to HG-SIL was compared with the group that did not progress to HG-SIL (RR: 3.667; p = 0.021; CI: 1.004-13.397). When other parameters were compared, no associations were observed.
Quantitative immunohistochemical expression of p16 according to the histological diagnosis at the second biopsy
The results of immunohistochemical distribution of p16INK4a related to HPV type at the second biopsy are presented in Table 3. With respect to HPV type, in the group that maintained an LG-SIL diagnosis at the second biopsy, no significant associations were found between quantitative p16INK4a overexpression and HPV 16 (p= 0.6155) or HPV 18 infection (p= 0.4316).
In the group that progressed from LG-SIL to HG-SIL in the second biopsy, p16INK4a overexpression was associated only with HPV 16 infection (RR, infinity; p= 0.0294). No association was found between p16INK4a overexpression and HPV 18 in the second biopsy (p= 0.4316).
bcl-2 Immunohistochemistry
In the first group of biopsies, 12/31 (38.7%) cases were negative for bcl-2 immunostaining, 4/31 (12.9%) cases showed weak staining, 9/31 (29.0%) moderate staining, and 4/31 (12.9%) intense staining. Two of 31 (6.45%) were classified as unsatisfactory slides (Figures 1A, 1B, and 1C).
In the second group of biopsies, irrespective of lesion grade, the basal epithelial layer was positive for bcl-2 immunostaining in 65.7% of the slides, and the glandular epithelium stained positively in 54.3% of the slides. Moreover, inflammatory stromal cells stained positively in 22.9% of the slides.
The results of immunohistochemical distribution of bcl-2 related to HPV type at the second biopsy are presented in Table 4 and Table 5. In the group that maintained LG-SIL at the second biopsy, no significant association was found between bcl-2 overexpression and HPV 16 infection (p= 1.00). In the group that progressed from LG-SIL to HG-SIL, bcl-2 overexpression observed by intensity and quantitative staining was not associated with HPV 16 infection (p= 0.664 and p= 0.795, respectively). No significant associations with HPV 18 infection were detected in either group.
p16 Immunostaining distribution according to HPV type at the second biopsy
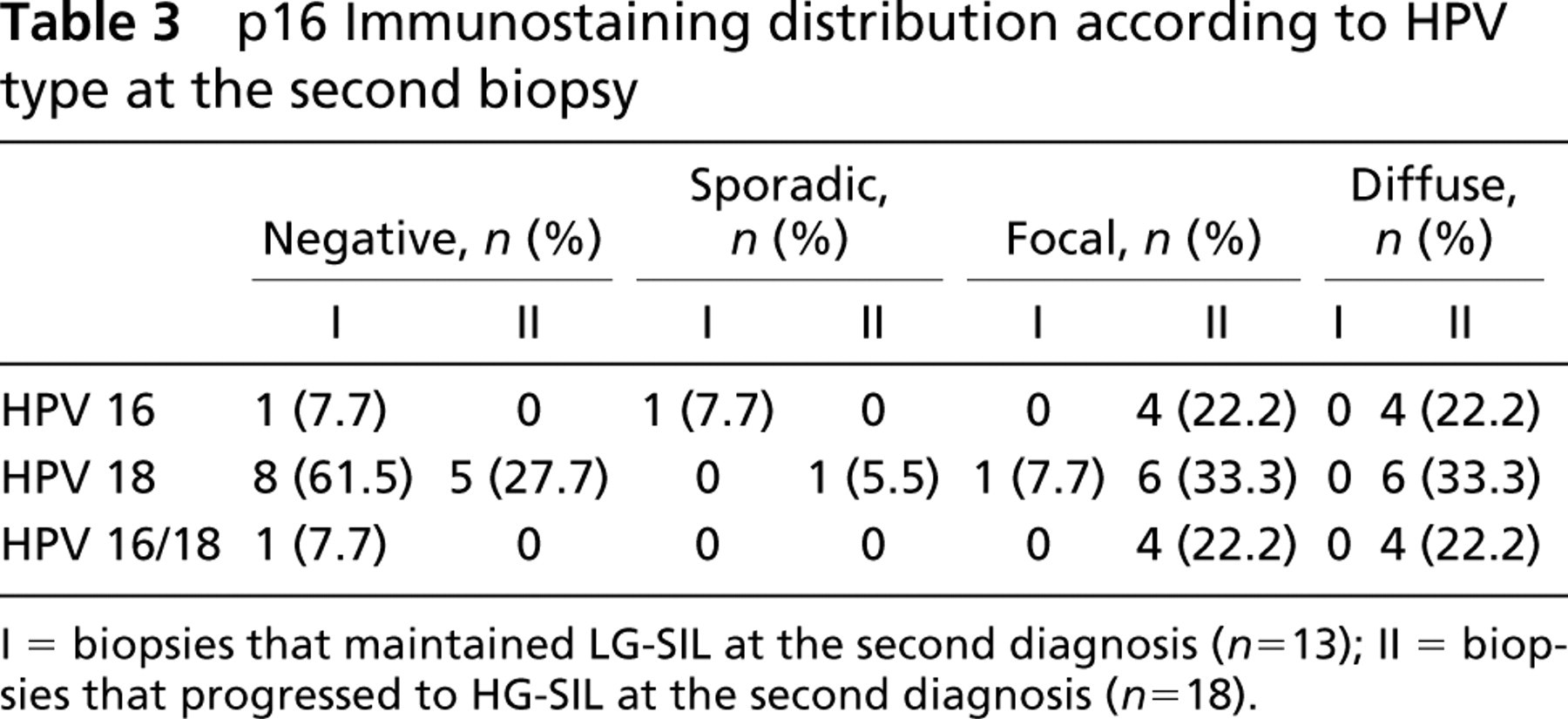
I = biopsies that maintained LG-SIL at the second diagnosis (n= 13); II = biopsies that progressed to HG-SIL at the second diagnosis (n= 18).
Quantitative immunohistochemical expression of bcl-2 according to the histological diagnosis at the second biopsy
Discussion
Abnormalities of several proteins of the cell cycle regulatory machinery and disruption of cell cycle controls, including p16INK4a, seem to be a common feature of several cancers. Contradictory results have been reported about the predictive value of p16INK4a in tumor progression. If, on the one hand, p16INK4a overexpression has been related to colorectal (Esteller et al. 2001), esophageal (Sturm et al. 2001), lung (Hayashi et al. 1994), oral (Fregonesi et al. 2003), pancreatic (Caldas et al. 1994) and cervical (Van de Putte et al. 2003) cancer development, on the other hand, other studies have associated absent or low p16INK4a expression with tumor progression, as observed in head/neck tumors (Namazie et al. 2002), neuroblastomas (Takita et al. 1998), melanomas (Straume et al. 2000), and endometrial carcinomas (Salvesen et al. 2000). In this study, we evaluated the potential of p16INK4a and bcl-2 immunohistochemical expression to predict the course of premalignant cervical lesions associated with HPV types 16 and 18.
Previous studies have demonstrated that negative or low p16INK4a expression is associated with LG-SIL, whereas p16INK4a overexpression is related to the presence of HG-SIL and invasive cervical cancer (Keating et al. 2001; Murphy et al. 2003). In the present series, significant p16INK4a overexpression was observed in the group that progressed from LG- to HG-SIL when compared with the group that did not progress. To our knowledge, this is the first study with a retrospective follow-up design carried out to evaluate SIL progression and the association of p16INK4a overexpression and HPV 16 infection. In fact, Sakaguchi et al. (1996) and Sano et al. (1998) were the first authors to relate p16INK4a overexpression to malignant cellular behavior. Later, other studies also reported a significant association between p16INK4a overexpression and HG-SIL related to high-risk HPV types (Klaes et al. 2001; Sano et al. 2002). It is possible that not all HPV types classified as high-risk possess the same potential for the cell cycle disruption or altered gene expression that leads to p16INK4a upregulation. Thus, the present results highlight the possible potential of p16INK4a as a marker for type-specific HPV-related HG-SIL and cervical cancer progression.
bcl-2 Immunostaining distribution according to HPV type at the second biopsy
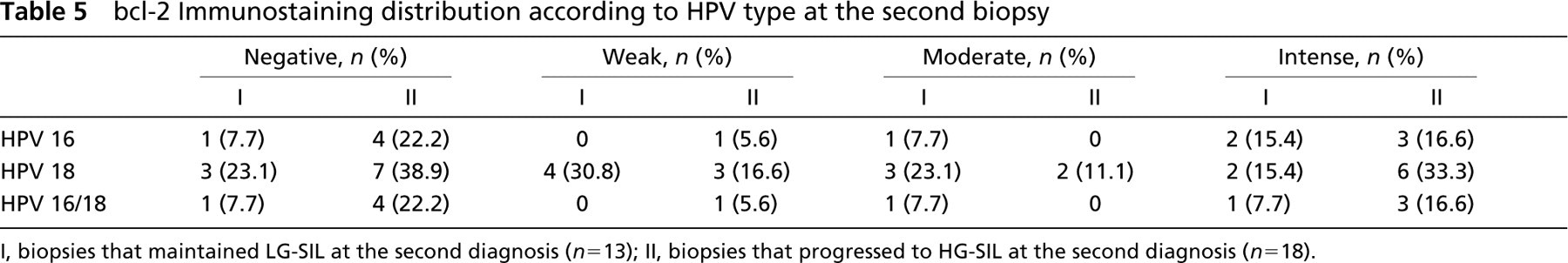
I, biopsies that maintained LG-SIL at the second diagnosis (n= 13), II, biopsies that progressed to HG-SIL at the second diagnosis (n= 18).
The analysis of genes involved in the regulation of apoptosis in cervical cancer is increasingly important for providing a better insight into the process of tumorigenesis and for determining which intraepithelial lesions are likely to progress to invasive carcinoma (Ciavattini et al. 1999). One of the genes is bcl-2, a cellular protein associated with cell membranes (plasma, endoplasmic reticulum, and mitochondria) that, in a complex form, prevents cell death under certain circumstances (Gillet and Brun 1996; Plas and Thompson 2002). Some studies have demonstrated that high bcl-2 expression may be related to a good prognosis in several cancers, such as breast, ovarian, and cervical cancer, as well as lymphoma (Crawford et al. 1998; Dimitrakakis et al. 2000). Conversely, the expression of bcl-2 protein was nearly absent in progressing oral lesions (Chang et al. 2002), and present at low levels in a microinvasive squamous cervical carcinoma (Kohlberger et al. 2003).
In the present series, bcl-2 expression was confined to the basal cell layer in all biopsies, regardless of histological grade or SIL progression. These results were in contrast to those of Ter Harmsel et al. (1996), which showed immuhistochemical bcl-2 overexpression in all cases of LG-SIL (CIN I) in basal layers, and in HG-SIL (CIN II and III), throughout the epithelium. With respect to bcl-2 intensity, we verified higher basal layer staining in lesions that progressed to HG-SIL, as previously reported by other authors (Ter Harmsel et al. 1996). Furthermore, we found bcl-2 overexpression in 6/13 (42.9%) of the biopsies that maintained the histological diagnosis of LG-SIL and negative or weak bcl-2 staining in 10/18 (55.4%) of the biopsies that progressed to HG-SIL. It is possible that low levels of bcl-2 in LG-SIL indicate a limited apoptotic protection, and thus only a small number of LG-SIL lesions will progress (Nasiell et al. 1986). The role of apoptosis-related proteins in cervical neoplasia might differ from that observed in other human cancers, and this relationship remains ill defined. Some authors conclude that bcl-2 expression may not be a good marker for predicting the progression of cervical lesions (Uehara et al. 1995; Jain et al. 2003). Longer follow-up studies are needed to better determine whether bcl-2 overexpression in LG-SIL will result in further cancer evolution or low bcl-2 expression in the group that progresses to HG-SIL will result in less malignancy.
There is evidence that E6 HPV oncoprotein targets p53 for proteolytic degradation, contributing to deregulation of the cell cycle. Inasmuch as p53 also controls bcl-2 transcription, p53 degradation may induce high bcl-2 levels, promoting cellular escape from the apoptotic pathway and finally contributing to malignant transformation (Werness et al. 1990; Kouzarides 1995). Interactions among bcl-2, p53, and Rb genes have been reported in a variety of systems, suggesting a relationship between unregulated apoptosis and a lack of tumor suppressor functions in malignant disease (Gillet and Brun 1996). This may indicate a complex linkage between HPV infection and apoptosis that we could not show in our study. No association between HPV infection and bcl-2 overexpression was found, in contrast to the results reported by Kurvinen et al. (1996) and in agreement with other recent studies (Ciavattini et al. 1999; Brychtova et al. 2000; Rihakova et al. 2001), suggesting that viral oncoproteins may not deregulate bcl-2 gene expression.
We conclude that HPV types 16 and 18 are involved in p16INK4a overexpression associated with SIL progression. Moreover, our study demonstrated that p16INK4a immunohistochemical evaluation could be useful as a biomarker for progressive malignancy in HPV-related cervical cancer. No apparent correlation was found between the presence of HPV-DNA and bcl-2 protein expression, suggesting that HPV itself is not directly involved in the apoptosis regulatory pathways that regulate apoptosis in cervical neoplasias. Thus, further studies are needed to determine whether viral oncoproteins are involved in other intrinsic or extrinsic components of deregulation in cervical cancer.
Footnotes
Acknowledgments
This work was supported by grants from Conselho Nacional de Desenvolvimento Científico e Tecnológico [National Council of Scientific and Technological Development] 133990/03-6 (MCMG), Fundação para o Desenvolvimento da UNESP [Foundation for the development of UNESP] 084/2002 (CPS), and Fundação de Amparo a Pesquisa do Estado de São Paulo [Foundation to Support São Paulo State's Research] 01/02908-2 (MAGG).
We wish to thank Ana Maria Rocha for excellent technical assistance.