
Abstract
The aim of the present study was to evaluate the expression of innate immunity receptors belonging to the Toll-like family in the neural plexuses of the different tracts of murine intestine, of the human ileum, and in lower dorsal root ganglia (DRGs) from where extrinsic afferents to these plexuses originate. Results obtained by immunohistochemistry and immunofluorescence on paraffin-embedded tissue and whole-mount preparations show that Toll-like receptors (TLRs) −3 and −7, recognizing viral RNA, and TLR4, recognizing lipopolysaccharide (membrane component of Gram-negative bacteria), are expressed in the myenteric and submucous plexuses of murine intestine and human ileum, and in DRGs primary sensory neurons. They also show that TLR4 immunostaining is stronger in murine distal large bowel. In murine tissue, expression of TLRs was present in both neurons and glial cells. These observations indicate that the enteric neural network might be directly activated by bacterial and viral components and is therefore more in the forefront than previously envisaged in defense responses of the intestinal wall and in the cross-talk with intestinal microbiota. They also highlight the presence of a peripheral neural network that by way of hardwired neurotransmission could potentially convey to the central nervous system specific information on our microbial counterpart and invading or potentially invading pathogens.
TLRs are pathogen recognition receptors that allow the innate immune system to rapidly recognize conserved microorganism-associated molecular patterns (MAMPs) of pathogenic, potentially pathogenic, or non-pathogenic and, moreover, useful commensal microorganisms. They can be subdivided into several subfamilies that recognize related MAMPs belonging to bacteria, fungi, parasites, viruses, and hosts (Akira et al. 2006). TLR activation by pathogens triggers rapid and localized responses mediated by phagocytes through different signaling pathways, leading to the production of proinflammatory cytokines, chemokines, and type 1 interferon (Akira et al. 2006). On the other hand, interaction of TLRs with intestinal microbiota appears to be rather multifaceted. Under normal conditions, it contributes to maintenance of the intestinal barrier and gut homeostasis, but when deregulated, it may lead to and/or precipitate inflammatory pathologies (Cario 2005; Harris et al. 2006).
The aim of the present study was to evaluate the expression of two other receptors of the Toll-like family, TLR3 and TLR7, recognizing viral double-stranded and single-stranded RNA respectively, in the ENS of the murine small and large intestine and of the human ileum. Moreover, because microbial density and predominance of Gram-negative bacteria increase from the proximal to the distal intestine (Hooper and Gordon 2001; Macpherson and Uhr 2004a,b), we also wanted to investigate whether a corresponding change in TLR4 immunostaining of enteric neurons could be observed. Additionally, TLR3, −4, and −7 expression has been evaluated in lower thoracic and lumbosacral murine DRGs.
Materials and Methods
Murine Tissue
C57BL/6N female mice, from 8 to 32 weeks old, were purchased from Charles River (Calco, Italy) and housed under specific pathogen-free conditions, maintained at constant temperature and humidity, with food and water given ad libitum. Animals were sacrificed by cervical dislocation. Handling and suppression were performed according to the regulations of the Università degli Studi di Milano. Following sacrifice, the intestinal tube was excised, laid out in a Petri dish, and dissected into its different portions under a steromicroscope. DRGs from lower thoracic and lumbosacral segments were collected from the appropriate intervertebral foramen under a surgical microscope.
Immediately after excision, samples were immersed in one of the following fixatives: 10% formalin in phosphate-buffered saline (PBS, 0.1 M, pH 7.4) for 4 hr or overnight at 4C, 4% paraformaldehyde in PBS for 4 hr or overnight at 4C, or Zamboni fixative (2% formalin, 0.2% picric acid in PBS) overnight at 4C.
Human Tissue
Small fragments encompassing the full thickness of the intestinal wall were obtained from the ileum of obese patients who underwent Scopinaro biliopancreatic diversion in the Department of Surgery, Multimedica Hospital, Sesto San Giovanni, Milan. All patients gave informed written consent. Ethical approval was obtained from the internal local board.
Human biopsies were fixed in 10% formalin in PBS overnight at 4C.
Antibodies
The following rabbit polyclonal antibodies against murine TLRs were used: TLR4 (ab13556 Novus Biologicals, Inc.; Littleton, CO); TLR3 (ab 210–367-R200, Alexis Biochemicals; Lausen, Switzerland); TLR7 (ab IMG-581A, Imgenex; San Diego, CA). Monoclonal antibody MS-280-PO directed against mouse glial fibrillary acidic protein (GFAP) was purchased from Lab Vision (Suffolk, UK).
Swine anti-rabbit immunoglobulins and rabbit peroxidase anti-peroxidase (PAP) were purchased from DAKO (Glostrup, Denmark). For the detection of mouse anti-mouse antibodies in murine tissue, the M.O.M. Kit (Vector Laboratories, Inc.; Burlingame, CA) was used.
Immunohistochemistry on Paraffin-embedded Tissue
Specimens were processed for paraffin embedding. Tissue sections (4-μm to 6-μm) were obtained from paraffin blocks and collected on silane-coated slides. For antigen retrieval, deparaffinized sections were either autoclaved for 6 min at 120C in sodium citrate buffer (0.01 M, pH 6) or treated in a microwave oven. Quenching of endogenous peroxidase activity was performed in 0.3% H2O2 in PBS. Nonspecific sites were blocked incubating slides for 30 min either with a solution consisting of 0.05 M Tris-HCl, 0.15 M NaCl, 0.1% gelatin, 0.5% ovalbumin, 0.05% Tween-20, and 0.2% fish gelatin, or with 10% goat serum. Afterwards, sections were incubated with one of the following primary antibodies: rabbit anti-mouse TLR4 diluted 1:50 in PBS overnight at 4C; rabbit anti-mouse TLR3 diluted 1:50 and rabbit anti-mouse TLR7 diluted 1:100 in PBS for 1 hr at 37C. After rinsing in PBS, sections were incubated in goat anti-rabbit diluted 1:100 in PBS for 1 hr at room temperature and then in rabbit PAP diluted 1:100 in PBS for 1 hr at room temperature. For the development of the reaction, a liquid DAB substrate chromogen system was used (DAB, DAKO). Slides were washed with PBS, dehydrated through an ascending series of ethanols, counterstained with hematoxylin, and mounted with entellan (Merck; Darmstadt, Germany). Controls were performed by replacing the primary antibody with non-immune rabbit serum or dilution buffer, by sequential omission of the secondary antibody and PAP complex, or by incubation with DAB reagent alone to exclude the possibility of non-suppressed endogenous peroxidase activity. Spleen was used as positive control.
Double Immunofluorescence on Paraffin-embedded Tissue
For double immunofluorescence experiments, deparaffinized mouse intestine sections were rinsed in Tris-buffered saline with 1% bovine serum albumin (TBS-BSA) and either autoclaved for 6 min at 120C in sodium citrate buffer (0.01 M, pH 6) or subjected to microwave treatment in sodium citrate buffer (0.01 M, pH 6) for antigen retrieval. To remove tissue autofluorescence, sections were processed with sodium borohydride in TBS (0.1%, three times, 10 min each at 4C). After washing with TBS-BSA, nonspecific binding sites were blocked with 1:10 goat serum in TBS-BSA. Incubation with primary antibodies was performed as follows: rabbit anti-mouse TLR4 diluted 1:50 in TBS-BSA overnight at 4C; rabbit anti-mouse TLR3 and rabbit anti-mouse TLR7 diluted 1:50 in TBS-BSA for 1 hr at 37C. After rinsing in TBS-BSA, sections were incubated with FITC-conjugated goat anti-rabbit antibody diluted 1:200 in TBS-BSA for 1 hr at room temperature. For GFAP detection, sections were subsequently washed in TBS-BSA, and nonspecific sites were blocked with mouse IgG blocking reagent (M.O.M. Kit) for 1 hr, followed by incubation with mouse anti-mouse GFAP for 1 hr at 37C (diluted 1:100 in M.O.M. diluent), with biotinylated anti-mouse IgG for 10 min, and with tetra-rhodamine isothiocyanate-conjugated streptavidin (Jackson Laboratories; West Grove, PA) diluted 1:200 in TBS-BSA for 30 min at room temperature. After several washes in TBS-BSA, sections were incubated with DAPI (Sigma-Aldrich; St. Louis, MO) diluted 1:300,000 in TBS for 5 min. Sections were then mounted with Mowiol 4–88 (Calbiochem; La Jolla, CA). Controls were treated as described above, omitting the primary antibodies.
Immunohistochemistry on Whole-mount Preparations
Segments of murine small intestine, 1 cm long, were fixed by immersion in 4% paraformaldehyde in PBS for 2 hr at 4C, opened along the mesenteric border, and washed in PBS. The longitudinal muscle layer with attached myenteric ganglia was peeled off under the dissecting microscope using fine forceps. This preparation and the remaining inner layers of the intestine containing the submucous plexus were washed in PBS and processed free-floating for immunolabeling with TLR3, TLR4, and TLR7.
Whole-mount preparations were permeabilized in TBS containing 0.3% Triton X-100 (TBS-T) for 30 min, and incubated in 0.3% H2O2 in TBS for 30 min to inactivate endogenous peroxidase and then in 0.1% Na2B4O7 in TBS for 10 min to remove excess aldehydes. After rinsing in TBS-T, nonspecific sites were blocked, incubating samples for 30 min in a solution consisting of 0.05 M Tris-HCl, 0.15 M NaCl, 0.1% gelatin, 0.5% ovalbumin, 0.05% Tween-20, and 0.2% fish gelatin. Whole-mounts were then incubated for 36 hr at 4C with one of the following primary antibodies: rabbit anti-mouse TLR4 diluted 1:50 in TBS-T, rabbit anti-mouse TLR3 diluted 1:50 in TBS-T, or rabbit anti-mouse TLR7 diluted 1:100 in TBS-T. After several rinsings in TBS-T, samples were incubated with the secondary antibody, goat anti-rabbit diluted 1:100 in TBS-T for 2 hr at room temperature, and then with rabbit PAP diluted 1:100 in TBS-T for 2 hr at room temperature. The reaction was developed using the liquid DAB substrate chromogen system. Preparations were collected on silane-coated slides, air dried, and mounted with entellan.
Immunohistochemical reactions on whole-mount preparations and paraffin sections were observed with a Nikon Eclipse 80i (Nikon; Tokyo, Japan) equipped with bright-field high-quality objectives [20×, Nikon, Plan Fluor, numerical aperture (NA) 0.50; 40×, Nikon, Plan Fluor, NA 1, oil immersion; 60×, Nikon, Plan Apo, NA 1.40, oil immersion]. Images were acquired by digital camera (Nikon Digital Sight 5MC) and image acquisition software (ACT-2U).
Evaluation of immunofluorescence samples was performed with a Nikon C1 laser scanning confocal unit (Nikon D-Eclipse C1) attached to an inverted fluorescence microscope (Nikon Eclipse Ti-U) with a 60× objective (Nikon, Plan Apo, NA 1.40, oil immersion). Excitation was obtained with an air-cooled argon laser (488 nm) and a helium/neon laser (561 nm). Appropriate filter sets were used to collect fluorophore emissions. Conditions were set to avoid photo bleaching. Images were acquired digitally and processed using the operation software EZ-C1 for Nikon C1 confocal microscope. For each field of observation, 40 consecutive optical sections were obtained (steps of 0.20 μm). Three-dimensional images and galleries of the optical sections were analyzed to evaluate the expression of TLR3, −4, and −7 in GFAP-positive cells.
Quantitative Evaluation of TLR3, TLR4, and TLR7 Immunohistochemistry in Murine Myenteric Plexus
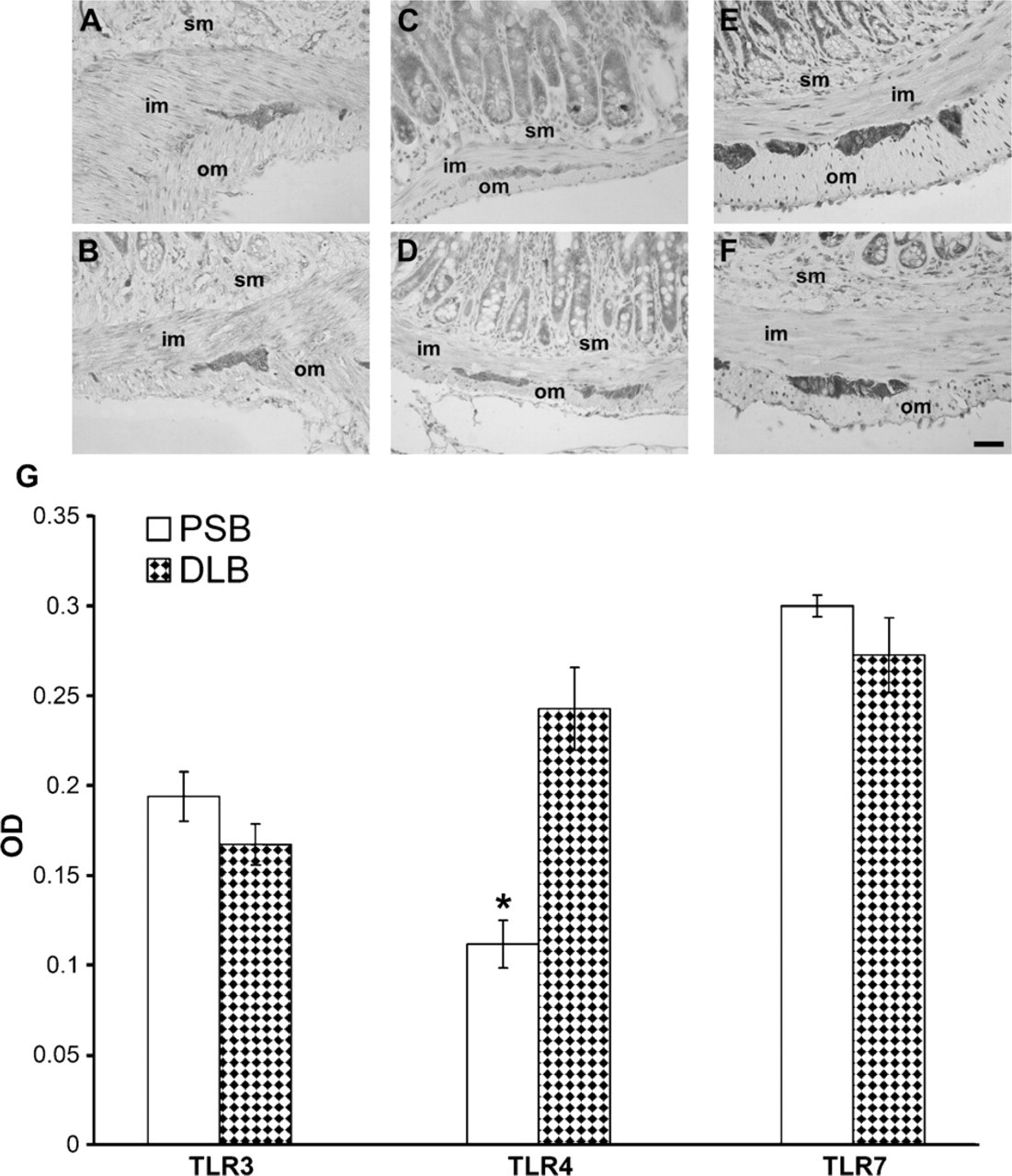
To evaluate differences in the level of immunostaining between the proximal small bowel and distal large bowel, staining intensity was computed as optical density (OD) and measured in 21 samples for the large intestine and 18 samples for the small intestine.
Digitally fixed images of slices at 40× magnification were analyzed using an image analyzer (Image Pro Plus 4.5.019, Media Cybernetics Inc.; Silver Spring, MD). For each ganglion, the unbiased OD was calculated by evaluating its OD and subtracting from this value the OD of an equivalent area surrounding the ganglion.
Data are presented as mean ± SEM. Statistical significance was evaluated by Student's t-test for unpaired samples and set at 1% (p=0.01).
Semiquantitative Evaluation of TLR4 Immunohistochemistry in Murine Myenteric Plexus
For semiquantitative evaluation of TLR4 immunostain-ing of murine myenteric plexus, sections from proximal small bowel and distal large bowel were scored independently by two observers for labeling intensity as follows: -, undetectable immunostaining; +, moderate immunostaining; ++, strong immunostaining.
Western Blotting
Proteins were extracted from DRGs by lysis in 0.001 M Tris-HCl, pH 7.6, 0.1 M NaCl, 0.001 M EDTA, pH 8, 1% Triton X-100, 10 mg/ml aprotinin, 10 mg/ml leu-peptin, 10 mg/ml pepstatin, and 100 mg/ml phenyl-methylsulfonylfluoride for 1 hr at 4C, followed by centrifugation for 15 min at 5000 rpm at 4C. The supranatant was then harvested at −80C. Proteins were estimated quantitatively using the BCA Protein Assay Kit (Pierce; Rockford, IL), measuring at 562 nm with a spectrophotometer (Ultrospec 2100 pro, Amersham Biosciences; Little Chalfont, UK). Protein samples (10 μg) were fractionated on an 8% acrylamide slab gel containing 0.1% SDS (Sigma-Aldrich) and transferred onto a nitrocellulose filter (Amersham Biosciences). As positive control, spleen extract was treated as described above. After incubation for 1 hr in TBS with 1% Tween-20 (Sigma-Aldrich) and 5% milk powder to block nonspecific binding sites, the filter was incubated with primary antibodies directed against rabbit anti-mouse TLR4 (1:200 in TBS, 0.1% Tween-20, and 1% milk powder) for 2 hr, and against rabbit anti-mouse TLR3 and rabbit anti-mouse TLR7 (both 1:500 in 1% TBS) for 1 hr at room temperature. After three washes of 20 min each in TBS, 1% Tween-20, and 1% milk powder, the filter was incubated with a goat anti-rabbit peroxidase-conjugated secondary antibody (1:1000 in TBS, 0.1% Tween-20, and 5% milk powder) for 1 hr at room temperature and then washed three times in 1% TBS for 10 min. Bands were visualized using enhanced chemiluminescence Western blotting detection reagents and autoradiography film (Amersham Biosciences).
Results
TLR3, TLR4, and TLR7 Immunolocalization in the Neural Plexuses of Murine Intestine and Human Ileum
Immunolocalizations performed on paraffin-embedded tissue (Figures 1 and 2) and whole-mount preparations (Figure 3), revealed that TLR3, −4, and −7 are expressed in the myenteric and submucous plexuses of murine small and large intestine. In whole-mount preparations, it was clearly visible that ganglia and nerve fibers connecting them were immunolabeled, as well as nerve fibers traveling in the muscle layers and in the submucosa (Figure 3). In the latter, association of immunopositive neurons with Peyer's patches was also observed (Figure 3E). Many TLR3-immunopositive fibers were endowed with varicosities (Figure 3A). TLR3 and TLR7 expression could also be detected in the myenteric plexus of the human ileum (Figure 4).
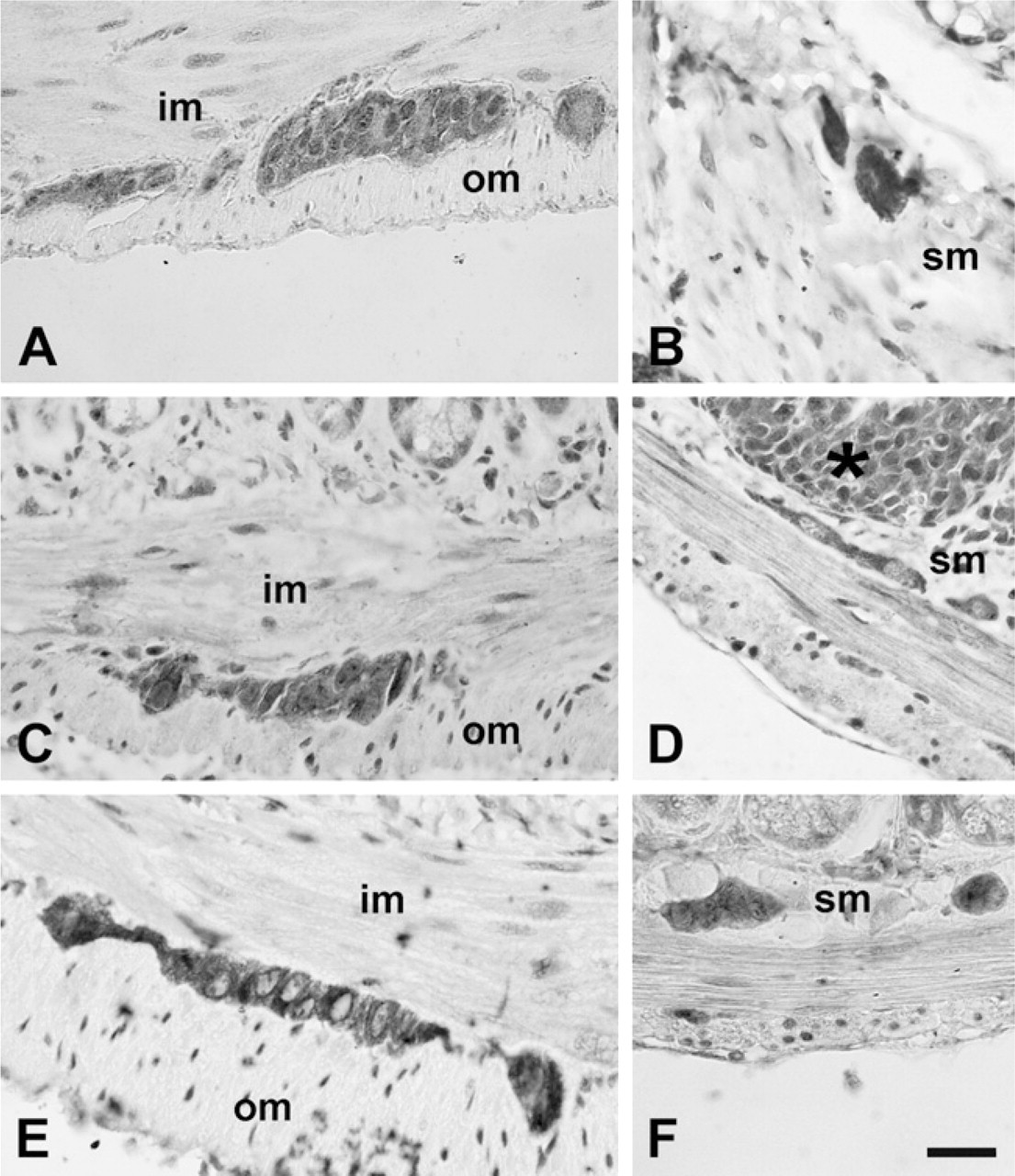
Immunohistochemical labeling for Toll-like receptor 3 (TLR3) (
In murine samples, the intensity of TLR4 immunostaining was stronger in the distal colon (Figure 5D) than in the proximal small bowel (Figure 5C), where weaker labeling intensity and lack of staining in some ganglia were visible. Semiquantitative (Table 1) and quantitative (Figure 5G) evaluations performed on sections from the proximal small bowel and from the distal large bowel confirmed this finding. Conversely, intensity of TLR3 (Figures 5A, 5B, and 5G) and TLR7 (Figures 5E-5G) immunostaining did not vary along the intestinal axis.
Double immunofluorescence of TLRs with the glial cell marker GFAP in murine tissue showed that GFAP-positive profiles were also positive for TLR3 and TLR4, whereas colocalization with TLR7 was not always present (Figure 2).
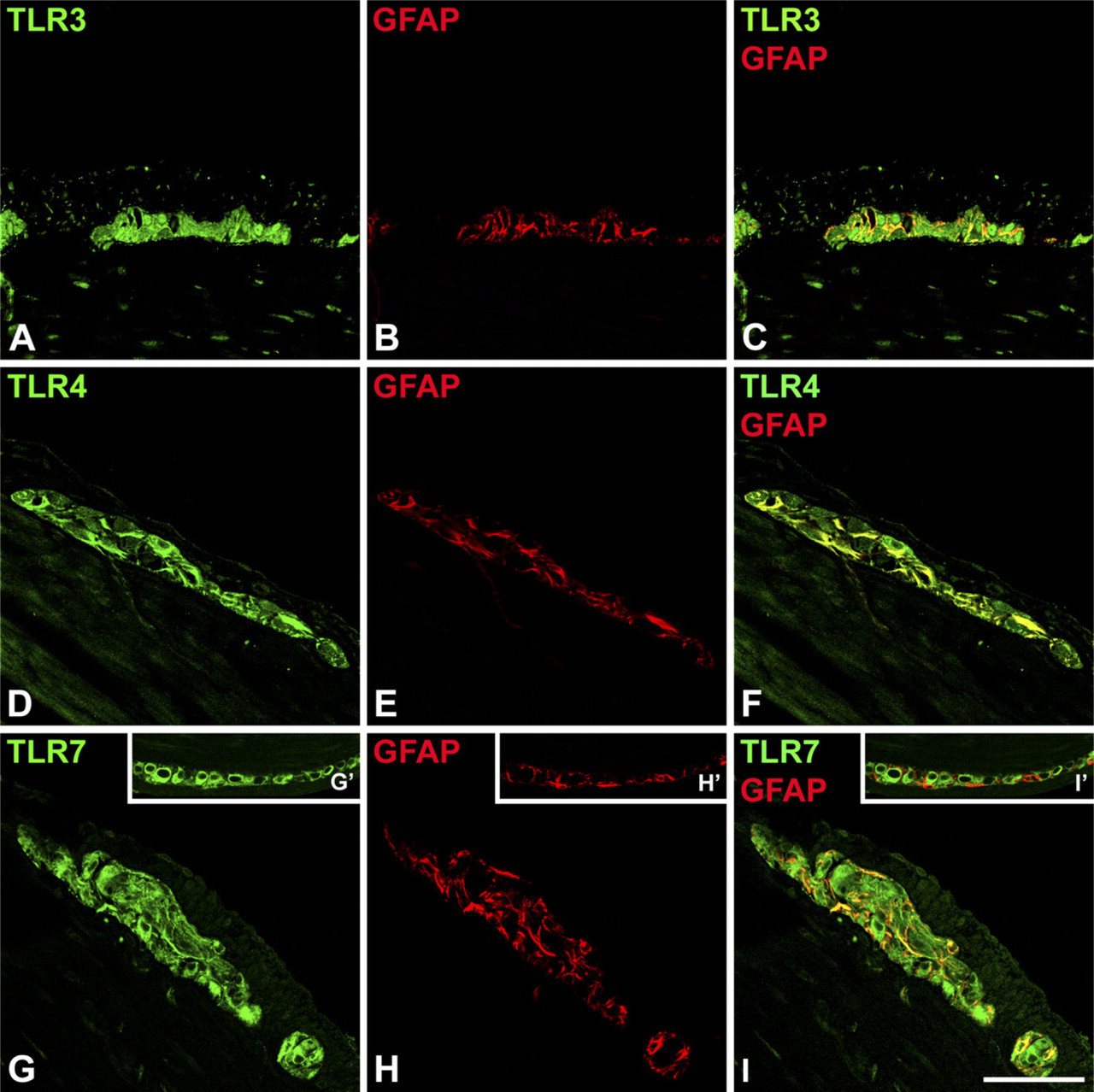
Confocal scanning microscopy analysis of TLR3 (
Immunolabeling could also be observed in Peyer's patches, in cells of probable macrophagic and lynfocitic nature in the submucosa, and in the epithelial lining. Labeling for TLRs was mostly cytoplasmic. However, in paraffin-embedded tissue, but apparently not in whole mounts, nuclear staining was present in some neurons (most notably for TLR3) and in smooth muscle cells. Specificity of the immunolabeling was verified using positive and negative controls as described in Materials and Methods. The different types of fixation did not change the quality of the immunoreactivity.
Expression of TLR4, TLR3, and TLR7 in Murine DRGs
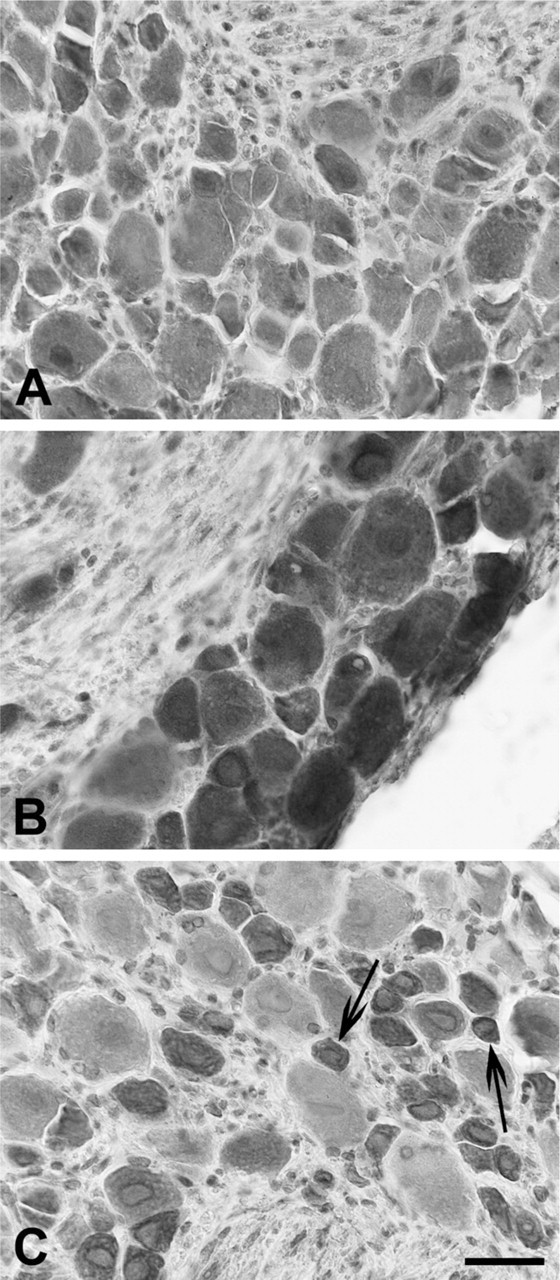
Immunohistochemistry (Figure 6) and Western blot analysis (Figure 7) showed that murine lower thoracic and lumbosacral DRGs express TLR3, TLR4, and TLR7.
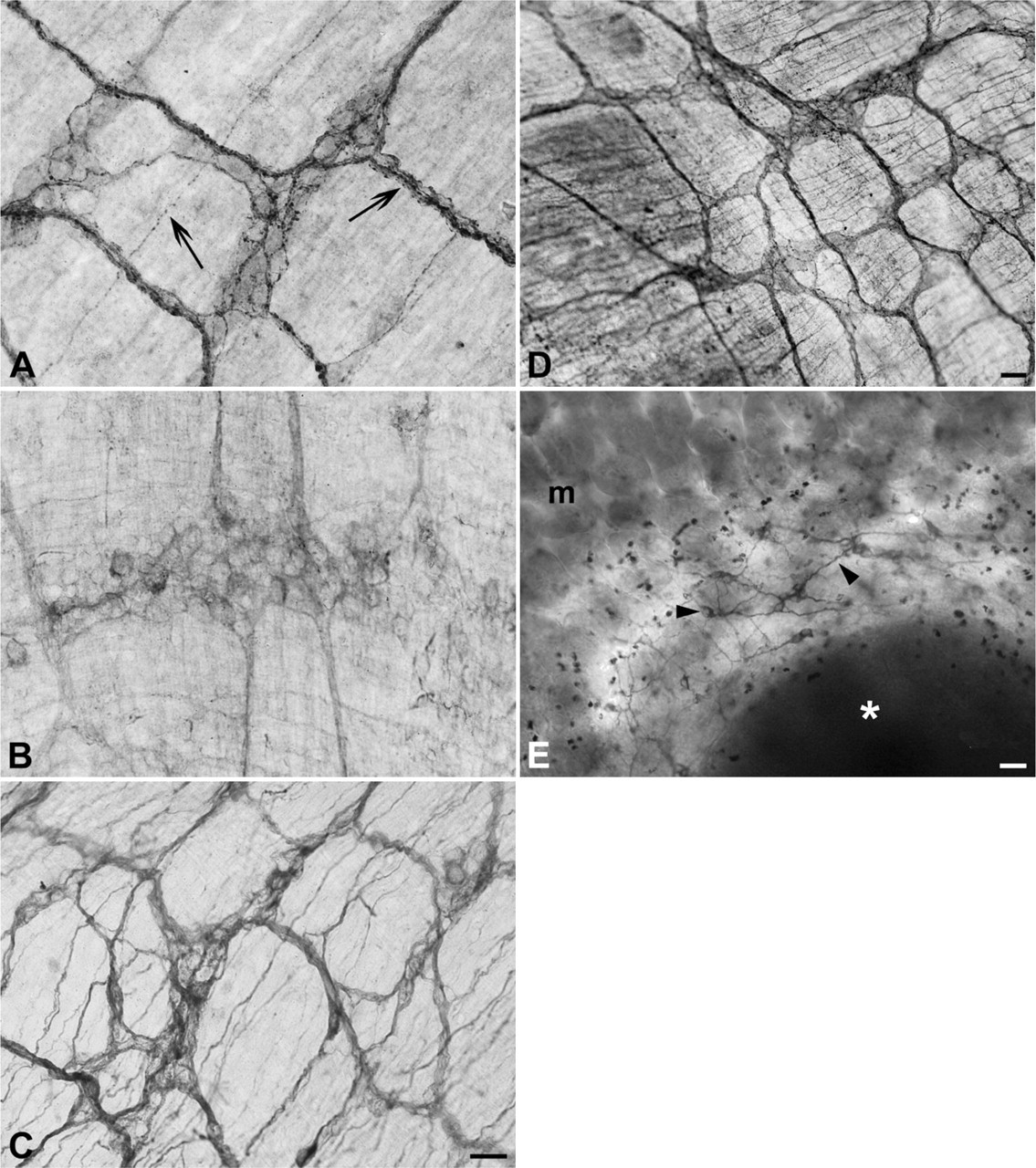
Whole-mount immunohistochemistry of murine small bowel. (
Cytoplasmic staining of neuronal cell bodies was often associated with nuclear or perinuclear labeling. Nerve fibers were also immunoreactive. TLR4 im-munolabeling was very strong in neurons of all sizes (Figure 6B), whereas for TLR3, the intensity of the immunostaining was more heterogeneously distributed (Figure 6A). In the case of TLR7, small-sized neurons were strongly reactive, compared with those of larger size, whose immunolabeling was much weaker or absent (Figure 6C).
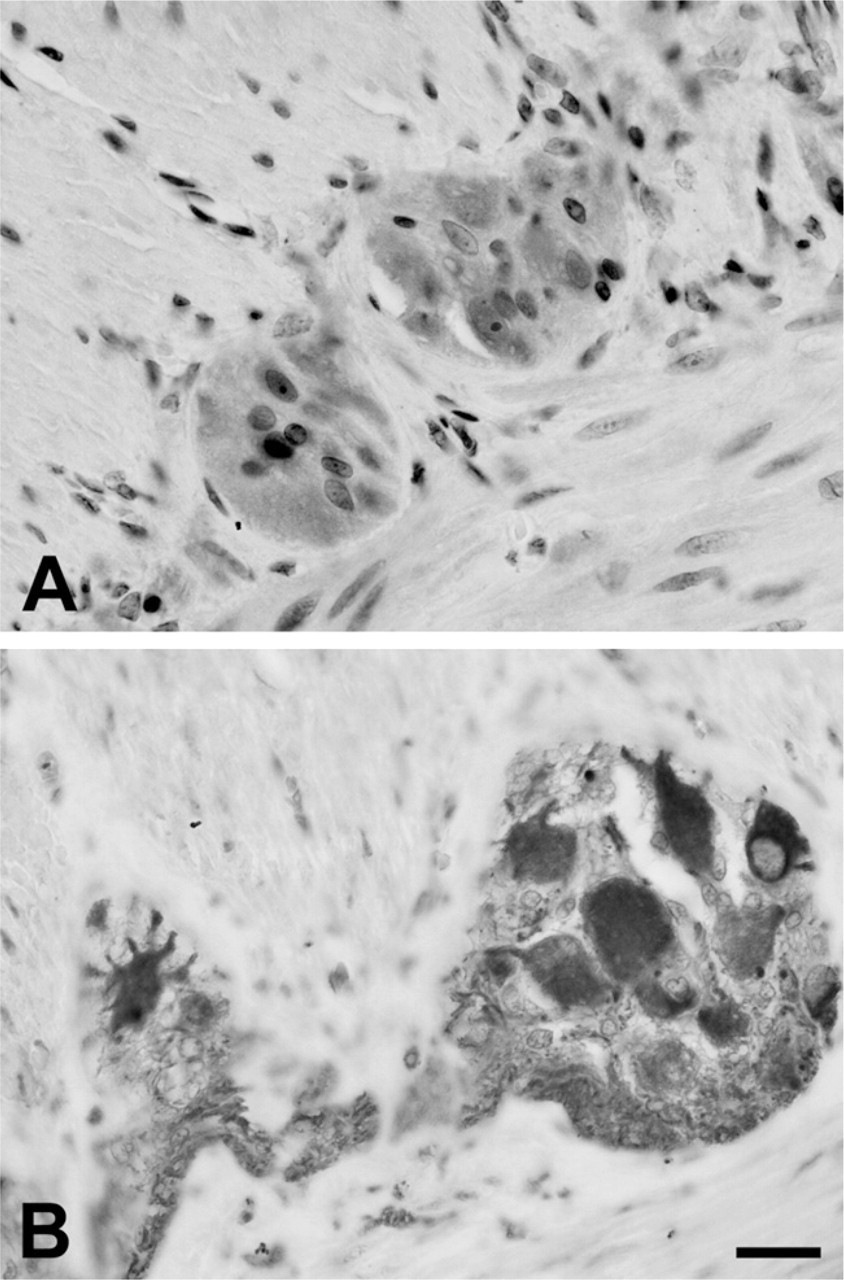
Immunolabeling for TLR3 (
Discussion
In the present study, we show for the first time that TLR3 and TLR7, recognizing viral RNA, are expressed in neurons and glial cells of the myenteric and submucous plexuses of murine small and large bowel and in plexuses of the human ileum. We also show that TLR4, besides being expressed in the neurons of the myenteric plexus of the murine jejunum (Rumio et al. 2006), is also expressed in the remaining intestine and in the sub-mucous plexus, in both neurons and glial cells, and that the intensity of its immunolabeling is stronger in the distal large bowel. Furthermore, we report the expression of the three receptors in lower thoracic and lumbosacral murine DRGs.
Expression of TLRs by the enteric neural network highlights the presence of a TLR-based neural surveillance system hardwired in the intestinal wall, and strongly suggests that viral and bacterial agents could directly activate intestinal neural responses without the interposition of immune or epithelial cells. The close proximity to a physiological and potentially pathological microbial environment might partly explain the difference in TLR expression between the ENS and the central nervous system (CNS). In the latter, relevant TLR expression can be observed in neurons only during pathological conditions (Jackson et al. 2006; Lafon et al. 2006), whereas it is constitutively present in glial cells and microglia (Bsibsi et al. 2002; Kielian 2006).
Closeness to a heavily loaded microbial environment (Hooper and Gordon 2001; Wood 2004) and adaptation to microorganisms normally present in the intestinal lumen might also explain the finding of stronger TLR4 immunolabeling in the distal colon, as also suggested by Ortega-Cava et al. (2003), who observed the same gradient in TLR4 mRNA and protein expression in the gut mucosa.
To date, cellular localization of TLR3 and TLR7 in the intestinal wall has not been much described and, to our knowledge, only Furrie et al. (2005) reported TLR3 expression in mature epithelial cells and lamina propria cells. Also, although TLR involvement in bacterial infections of the gastrointestinal tract has been addressed in several studies (Cario 2005; Harris et al. 2006), much less is known about the contribution of TLRs to the intestinal responses to viruses (Harris et al. 2006). The gastrointestinal tract provides the main route of access to the host for several viruses (Oberste et al. 2000), including the human immunodeficiency virus (Smith et al. 2003) and poliovirus (Mueller et al. 2005), and for unconventional viruses or agents such as those involved in prion diseases (McBride et al. 2001). These viruses can cause gastroenteritis, which is characterized by electrolyte and fluid secretion that seems to be triggered by local neural reflexes whose mechanism of activation is at present unknown (Morris and Estes 2001; Kordasti et al. 2004). Some enteroviruses also show different degrees of neurotropism and neuro-virulence that can lead to overt cytotoxic effects, latent infection, or viral spreading to the CNS (Irie et al. 1989; Morrison et al. 1991; Chen et al. 2003; Holland-Cunz et al. 2006; Selgrad et al. 2009). Although we did not perform any functional evaluation of TLR3 and TLR7 expression in enteric neurons and glial cells in this study, it is likely that they may be part of the signaling pathways that neurons have developed to respond to viral invasions (Patterson et al. 2002; Orvedahl and Levine 2008). In the CNS, a defensive role for TLR3 in neurons and glial cells has been demonstrated (Carpentier et al. 2005; Lafon et al. 2006; Daffis et al. 2008), whereas for TLR7, very few data are available (Lewis et al. 2008).
In our study, we observed widespread cytoplasmic labeling of neurons and fiber tracts connecting ganglia. Very little is known about the intracellular trafficking of TLRs in neurons (Lafon et al. 2006; Ménager et al. 2009). However, data are available from other cell types. Viral TLRs mostly localize to the endoplasmic reticulum and to endosomes where contact with viral nucleic acids takes place (Kawai and Akira 2007). In neurons, endocytic activity is also present in the somato-dendritic domain, and following endocytosis, endosome-containing viral particles have been shown in the different compartments of the cell, including dendrites and axon (Lewis and Lentz 1998). TLR4 is classified as a membrane receptor. Nonetheless, there are several reports that indicate a cytoplasmic localization as well, including the Golgi apparatus and microtubules (Uronen-Hansson et al. 2004; Hassan et al. 2006).
Immunolabeling for TLR3 (
Semiquantitative analysis of TLR4 immunolabeling of murine myenteric plexus in proximal small bowel and distal large bowel a
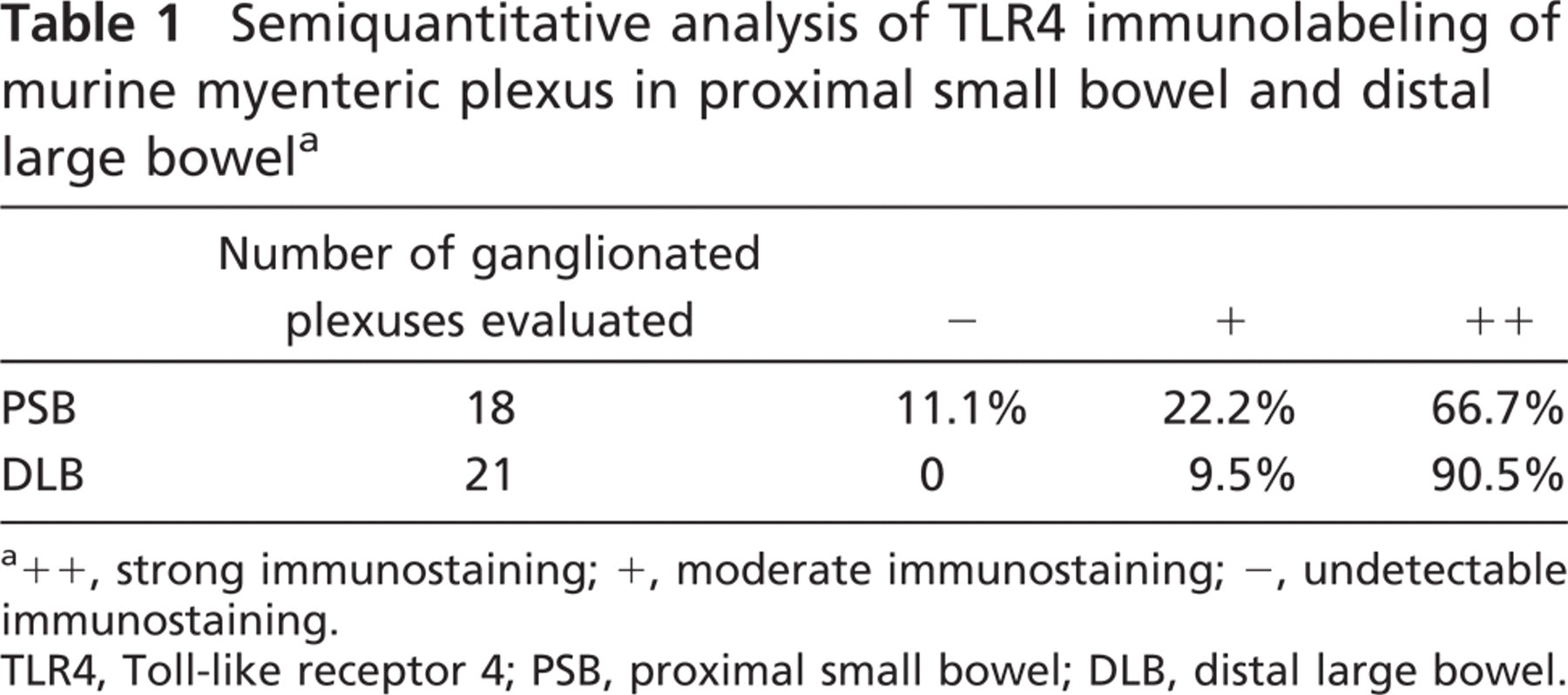
a++, strong immunostaining; +, moderate immunostaining; -, undetectable immunostaining.
TLR4, Toll-like receptor 4; PSB, proximal small bowel; DLB, distal large bowel.
Under normal conditions, the ENS does not appear to be directly exposed to the luminal content, inasmuch as neural projections into the intestinal lumen have not been demonstrated. The first possible site of interaction between enteric neurons and microbial components seems therefore to be limited to the subepithelial compartment. However, several pathological events of infective and non-infective nature cause disruption of the epithelial layer integrity (Svensson et al. 2004; Laukoetter et al. 2008), and bacteria and viruses can also translocate across the integer epithelial barrier by way of specialized cells able to convey them to underlying immune cells located in the mucosa and Peyer's patches (Bomsel and Alfsen 2003; Niedergang and Kweon 2005). At these sites, close apposition between immune cells and nerve fibers has been demonstrated (Ma et al. 2007; Vulchanova et al. 2007). In addition to pathological conditions, bacterial translocation is also normally present and contributes to gut-associated lymphoid tissue development, induction of tolerance to commensal microbiota, and constant stimulation of immune cells (Macpherson and Uhr 2004a,b). Physiological “close encounters” between enteric neurons and commensal microbiota are therefore not unlikely and might be at the base of a steady stimulation of neuro-immunomodulatory mechanisms participating in gut homeostasis. Modulatory effects of prebiotics and probiotics on nerve cells have already been demonstrated for enteric neurons (Kamm et al. 2004), DRGs (Kamiya et al. 2006), and rat brain neurons or pheochromocytoma-differentiated cells (Sobol et al. 2005).
Immunolabeling for TLR3 (
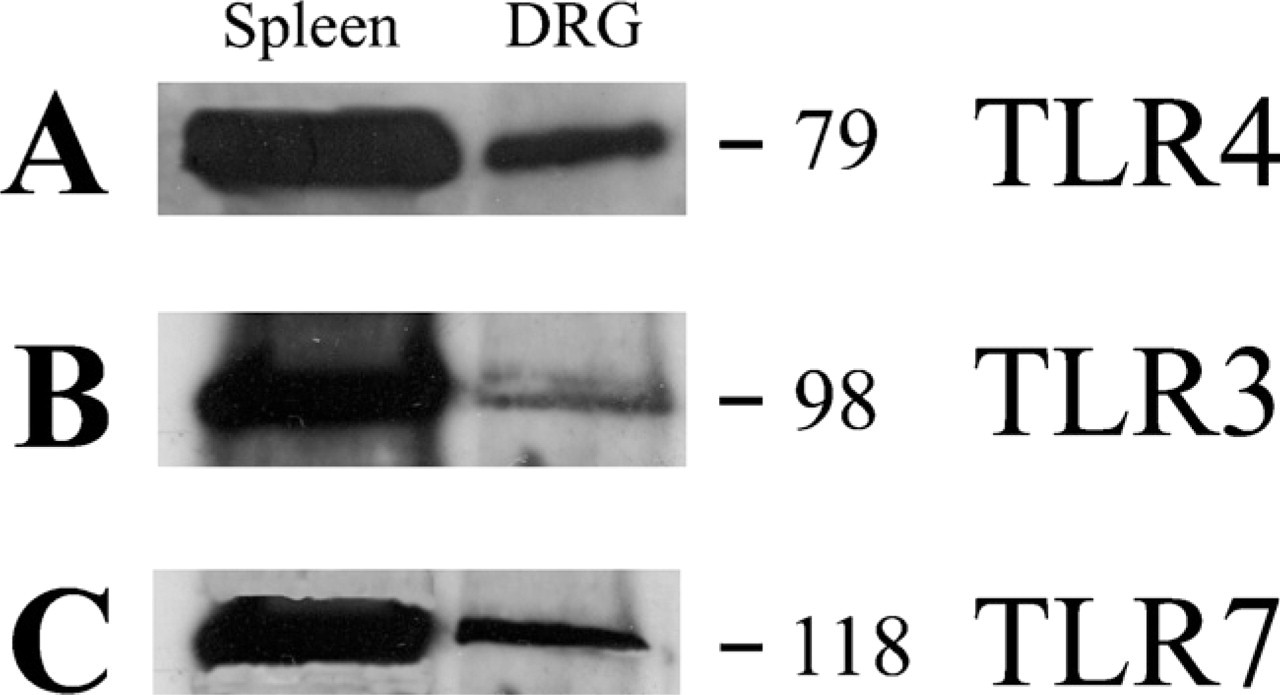
Western blot analysis of protein extract from mouse lower thoracic and lumbosacral DRGs, probed with the antibodies anti-TLR4 (
We also investigated TLR3, TLR4, and TLR7 expression in lower thoracic and lumbosacral DRGs. This part of the study was undertaken to determine whether spinal ganglia harboring neurons projecting to the intestine expressed TLRs and for the more general purpose of ascertaining whether TLR expression could be considered a somehow diffuse feature of the adult peripheral nervous system. Although we did not specifically identify primary sensory neurons projecting to the intestine, it is reasonable to assume that they express the three receptors on the basis of the following observations: different degrees of TLR3 and TLR4 immunoreactivity could be observed in all DRGs neurons examined; and strong TLR7 immunostaining was present in all small- to medium-sized neurons—the size range of visceral sensory neurons.
The presence of a direct mechanism of recognition of MAMPs in primary sensory neurons could be very meaningful when considering both the afferent function and efferent function of these cells (Holzer and Maggi 1998). By directly recognizing a characteristic microbial component and not only a generic inflammatory event, DRGs sensory neurons could be the first element in a labeled neural pathway, conveying to the CNS early and specific information on the nature and localization of microorganisms detected in the intestinal wall. Involvement of sensory pathways in conveying non-circulating microorganism-related signals has already been shown for the vagus nerve (Goehler et al. 2005,2007), and because its sensory ganglion expresses TLR4 (Hosoi et al. 2005), a direct activation by pathogens has been hypothesized (Goehler et al. 2007). It has also been shown that electrophysiological responses of DRGs sensory neurons projecting to the intestine can be modulated by oral inoculation of probiotics (Kamiya et al. 2006).
As to their efferent function, our findings suggest that TLRs might be part of the signaling pathways through which sensory neurons participate in rapid protective actions of the intestinal wall. These actions include activation of local axon reflexes such as the anti-inflammatory reflex and peripheral release from sensory endings of different mediators and neuropeptides in response to local threats (Holzer and Maggi 1998; Holzer 2006,2007).
Footnotes
Acknowledgements
We thank Dr. Michele Sciarabba for help with quantitative analysis of TLR4 immunostaining, Marianna Gaman for help in tissue processing, and Dr. Elena Donetti for kindly revising the manuscript.