
Abstract
Thymic mesenchymal cells are known to be important for the development of the early fetal thymus into a functionally mature organ supporting T cell differentiation. We examined the expression of mesenchymal markers: pan-mesenchymal marker ER-TR7, desmin, α-smooth muscle actin (α-SMA), and α- and β-chain of platelet-derived growth factor receptor (PDGFRα, PDGFRβ) in thymi of normal adult mice. Desmin and ER-TR7 revealed specific staining in the capsule, septa, and perivascular cells. Most perivascular cells highly expressed PDGFRβ at the same levels as desmin. Low expression of PDGFRα was detected in the capsule, intralobular septa, and some perivascular cells of normal adult thymi. α-SMA, used to identify vascular smooth muscle cells, was detectable on arterioles and some large venules but not on capillaries. Thus, desmin, PDGFRα, and PDGFRβ were localized in the capsule, septa, and perivascular cells in thymus of adult mouse, although there were differences in the expression level among these markers. On the other hand, the expression of mesenchymal markers was detectable in the region of the thymic medullary epithelium of lymphotoxin β receptor–deficient mice and plt/plt mice, indicating that mesenchymal cells were abnormally localized in the region. These results suggest that disorganization of the medullary epithelium may be accompanied by aberrant distribution of mesenchyme in adult mouse thymus.
T
Blood vessels are composed of endothelial cells and perivascular cells. Perivascular cells include pericytes and vascular smooth muscle cells (VSMCs) (Hirschi and D'Amore 1996). Small blood vessels are composed of endothelial cells surrounded by a basal lamina and are loosely covered by a single layer of pericytes. Large veins are irregularly covered by pericytes and/or VSMCs, whereas arteries have strong, elastic vessel walls with dense layers of concentrically formed VSMCs (Cleaver and Melton 2003). VSMCs are commonly identified by α-smooth muscle actin (α-SMA). Perivascular cells are known to express mesenchymal markers, such as desmin, PDGFRα, and PDGFRβ, but the expression of these markers is highly varied among vessel types in tissues and organs (Franklin et al. 1990; Nehls and Drenckhahn 1993; Kitami et al. 1995).
Mesenchymal cells of fetal thymus are shown to originate primarily in neural crest–derived cells (Bockman and Kirby 1984; Jiang et al. 2000). A contribution of neural crest to perivascular mesenchyme was noted earlier for the developing chick thymus (Le Douarin and Jotereau 1975). Furthermore, neural crest provides pericytes and smooth muscle cells to all blood vessels of the head and neck region (Noden et al. 1995; Etchevers et al. 2001). Recent studies have demonstrated that neural crest–derived cells enter the thymus, and these cells differentiate into perivascular cells, which contribute to a postulated thymus–blood barrier (Foster et al. 2008; Müller et al. 2008). In the thymus of adult mouse, mesenchymal cells, including perivascular cells, fibroblasts, and the cells residing in capsule and septa, are detected with an antibody, ER-TR7 (van Vliet et al. 1986). However, the expression of other mesenchymal markers in adult thymus has not been studied in detail.
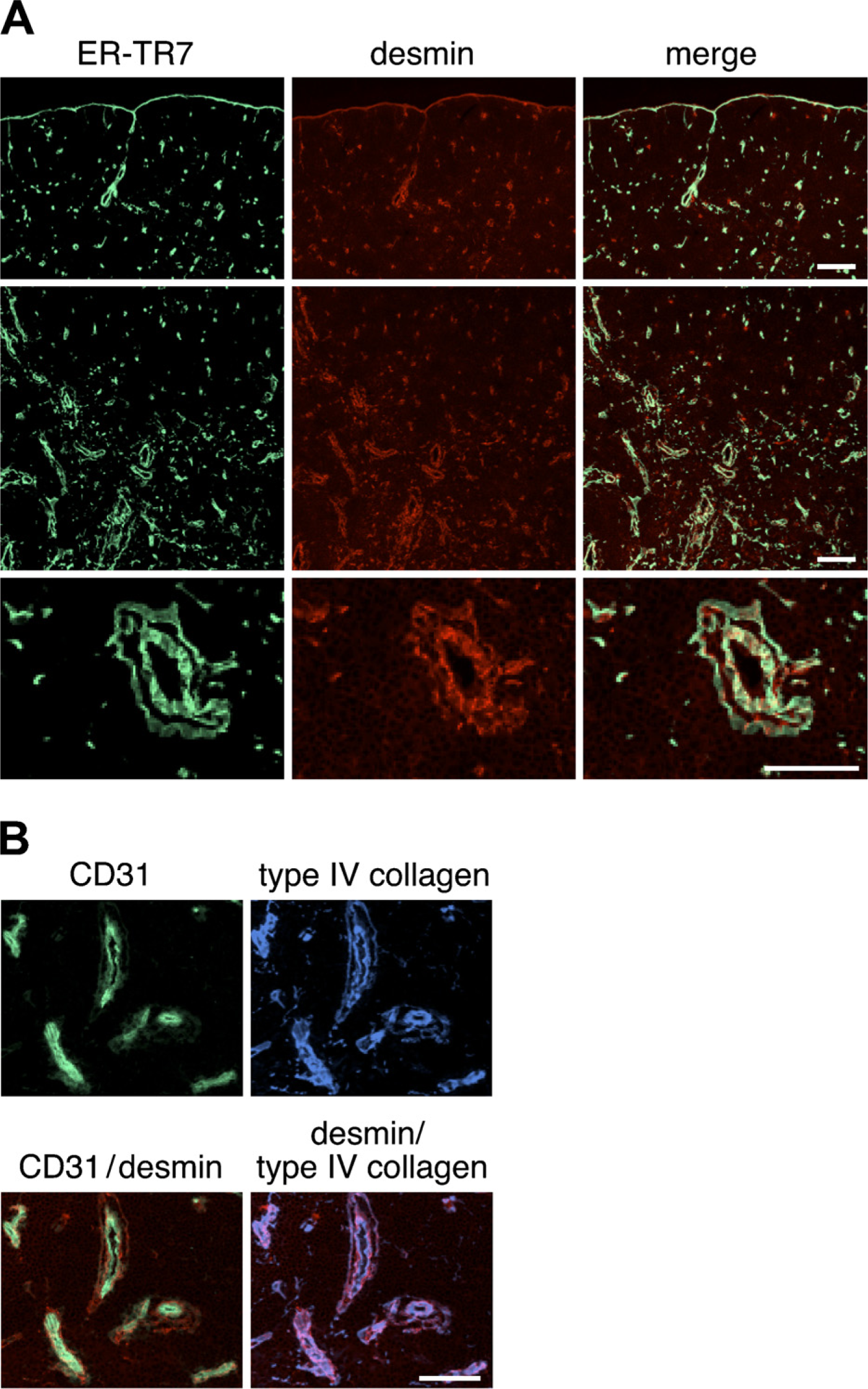
Expression of desmin in the thymus of adult mouse. Immunofluo-rescence staining of thymic sections of adult BALB/c mice was performed to detect ER-TR7 (
The lymphotoxin (LT) pathway is critical for the development and maintenance of peripheral lymphoid organs (Ware 2005). Mice with deficiencies in members of this pathway lack lymph nodes and Peyer's patches and have abnormal spleen architecture. These animals also develop autoantibodies and lymphocytic infiltrates of multiple organs, provoking speculation that the LT pathway may play a role in central tolerance induction (Chin et al. 2006). LT β receptor (LTβR) deficiency leads to abnormally differentiated thymic medullary epithelium (Boehm et al. 2003). In addition, the thymi of mice deficient in chemokine (C-C motif) receptor 7 (CCR7) or in its ligands CCL19 and CCL21-ser (plt/plt mice; Vassileva et al. 1999; Luther et al. 2000) also display altered architecture (Misslitz et al. 2004).
In this study, we examined thymic sections of normal adult mice by immunofluorescence analysis with antibodies against several mesenchymal markers to clarify the localization of mesenchyme in the thymi. Moreover, the analysis for thymi of LTβR-deficient (LTβR-/-) mice and plt/plt mice revealed that mesenechymal cells were abnormally distributed in the medullary region.
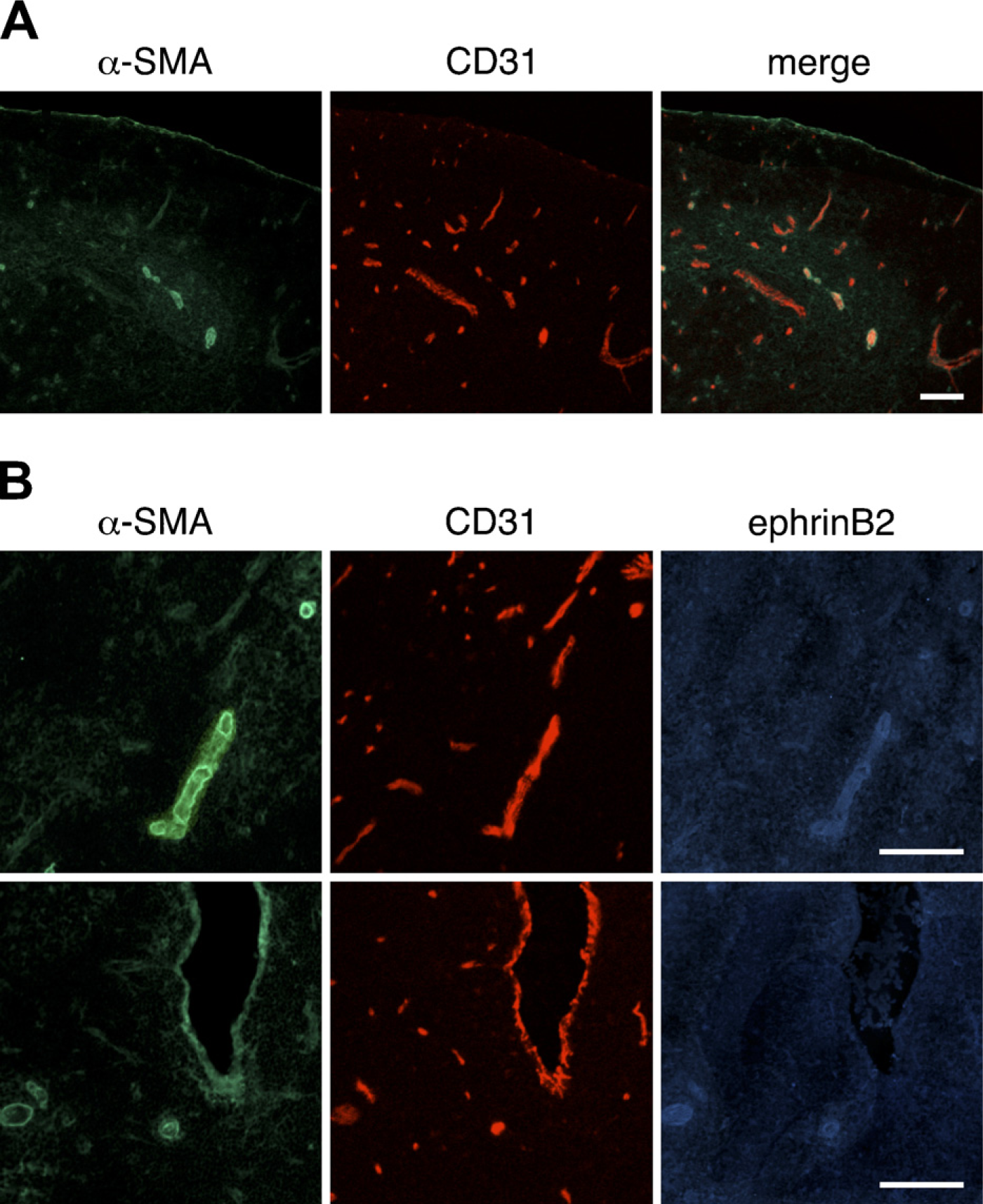
Expression of α-smooth muscle actin (α-SMA) in the thymus of adult mouse. Immunofluorescence staining of thymic sections of adult BALB/c mice was performed to detect α-SMA (
Materials and Methods
Mice Thymi
C57BL/6 and BALB/c mice were obtained from Japan SLC (Hamamatsu, Japan). Mice were sacrificed, and thymi were dissected, embedded in OCT compound, and frozen on dry ice.
Thymi of LTβR-/- mice on the B6 background (Chin et al. 2006) and plt/plt mice were gifts from Dr. Y-X. Fu (University of Chicago, Chicago, IL) and Dr. J.G. Cyster (University of California, San Francisco, CA). Mice were used at the age of 6 or 7 weeks. Animal care and experiments were reviewed and approved by the animal care committees of our institutes.
Antibodies and Reagents
The following antibodies and reagents were used: rat ER-TR7 (BMA Biomedicals; Augst, Switzerland), mouse anti-desmin (clone D33; DakoCytomatin, Glostrup, Denmark), rat anti-PDGFRα (CD140α) (clone APA5; eBioscience, San Jose, CA), rat anti-PDGFRβ (CD140β) (clone APB5; eBioscience), mouse anti-α-SMA (clone 1A4; Lab Vision, Fremont, CA), rat anti-CD31/platelet endothelial cell adhesion molecule 1 (clone MEC13. 3; eBioscience), goat anti-ephrinB2 (R and D Systems, Minneapolis, MN), rabbit anti-keratin 5 (Covance; Berkeley, CA), and rabbit anti-collagen type IV (LSL CO., LTD.; Tokyo, Japan). Alexa Fluor–labeled donkey secondary antibodies (Molecular Probes; Eugene, OR) were also used. The binding to biotinylated Ulex europaeus agglutinin-1 (UEA-1; Vector Laboratories, Burlingame, CA) was followed by allophycocyanin-conjugated streptavidin (eBioscience).
Immunofluorescence Staining and Confocal Microscopy
Cryosections were prepared, fixed in ice-cold acetone for 15 min, and air-dried. After washing with PBS, sections were blocked with 20% Block Ace (Dainippon Pharmaceutical Co.; Osaka, Japan) in PBS for 1 hr. For mouse monoclonal antibodies, the Mouse on Mouse (M.O.M.) Kit (Vector Laboratories) was used, following the manufacturer's instructions. Sections were then incubated with primary antibodies at 4C overnight, followed by washing for 5 min three times with PBS. Secondary antibodies were then applied and incubated for 1 hr. Sections were washed and mounted with fluorescent mounting medium (Vector Laboratories) using coverslips. Confocal laser-scanning microscopy analysis was performed on a Zeiss LSM 510 (Carl Zeiss; Oberkochen, Germany). Negative controls were performed by replacement of first-step antibodies with isotype-matched monoclonal antibodies or species-matched antibodies. Representative images were chosen from each experiment for figure editing.
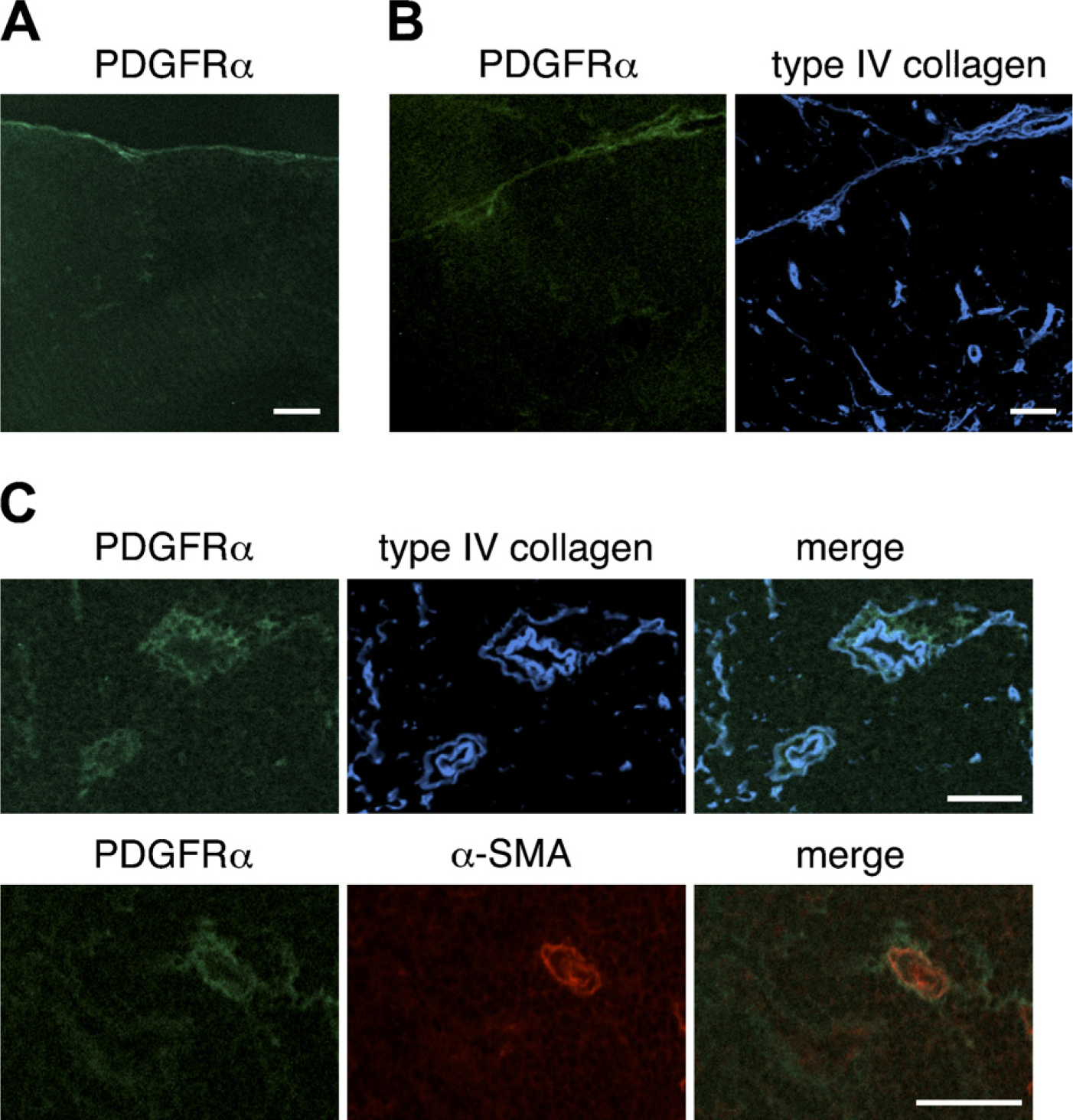
Expression of the α-chain of platelet-derived growth factor receptor (PDGFRα) in the thymus of adult mouse. Immunofluorescence staining of thymic sections of adult BALB/c mice was performed to detect PDGFRα (
Results
Expression of Desmin
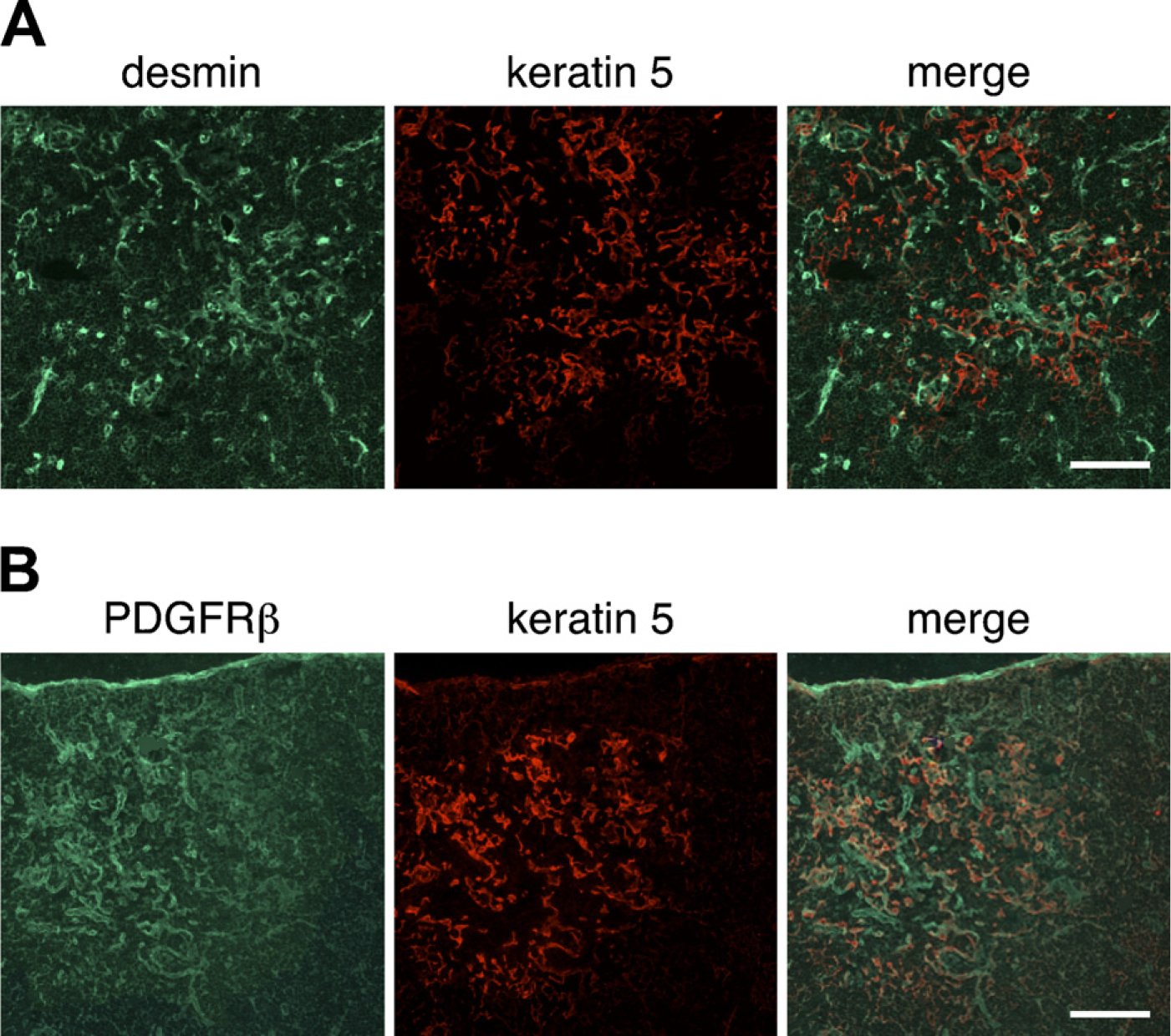
Frozen thymus sections from normal 6-week-old BALB/c mice were stained with ER-TR7 and antidesmin antibody, and the distribution of mesenchymal cells was analyzed by confocal microscopy. As expected from a previous study (van Vliet et al. 1986), ER-TR7 particularly marked the cells in the capsule, septa, and around blood vessels (Figure 1A). The distribution of desmin was similar to that of ER-TR7–positive reticular fibroblasts. Desmin-immunoreactive cells were situated near vasculatures including capillaries of the cortical region and postcapillary venules (PCVs) of the cortico–medullary junction (Figure 1A). Comparison with the pan-endothelial marker CD31 revealed that desmin-positive cells were localized in the wall of CD31-positive blood vessels including capillaries (Figure 1B), and desmin was restricted to perivascular cells in almost all blood vessels. When an antibody to type IV collagen was used to detect basement membranes, desmin-positive perivascular cells were seen to be associated with them (Figure 1B).
Expression of α-SMA
Thymus sections of BALB/c mice were stained for VSMCs with an anti-α-SMA antibody, and the resulting α-SMA immunoreactivity was found to be restricted to some vasculature-like structures (Figure 2A). Arterial and venous endothelial cells are phenotypically distinct, and ephrinB2 is selectively expressed on arteries (Gale et al. 2001; Shin et al. 2001). To clearly discern the distribution of α-SMA in thymic vasculatures, three-color immunofluorescence staining for α-SMA, CD31, and ephrinB2 was performed (Figure 2B). α-SMA-positive VSMCs were predominately localized on ephrinB2-positive arterioles. VSMCs on arterioles were uniformly shaped, closely packed, and tightly associated with the endothelium. VSMCs were also detectable on some large ephrinB2-negative PCVs, and PCVs partially covered with VSMCs were also detectable (Figure 2B).
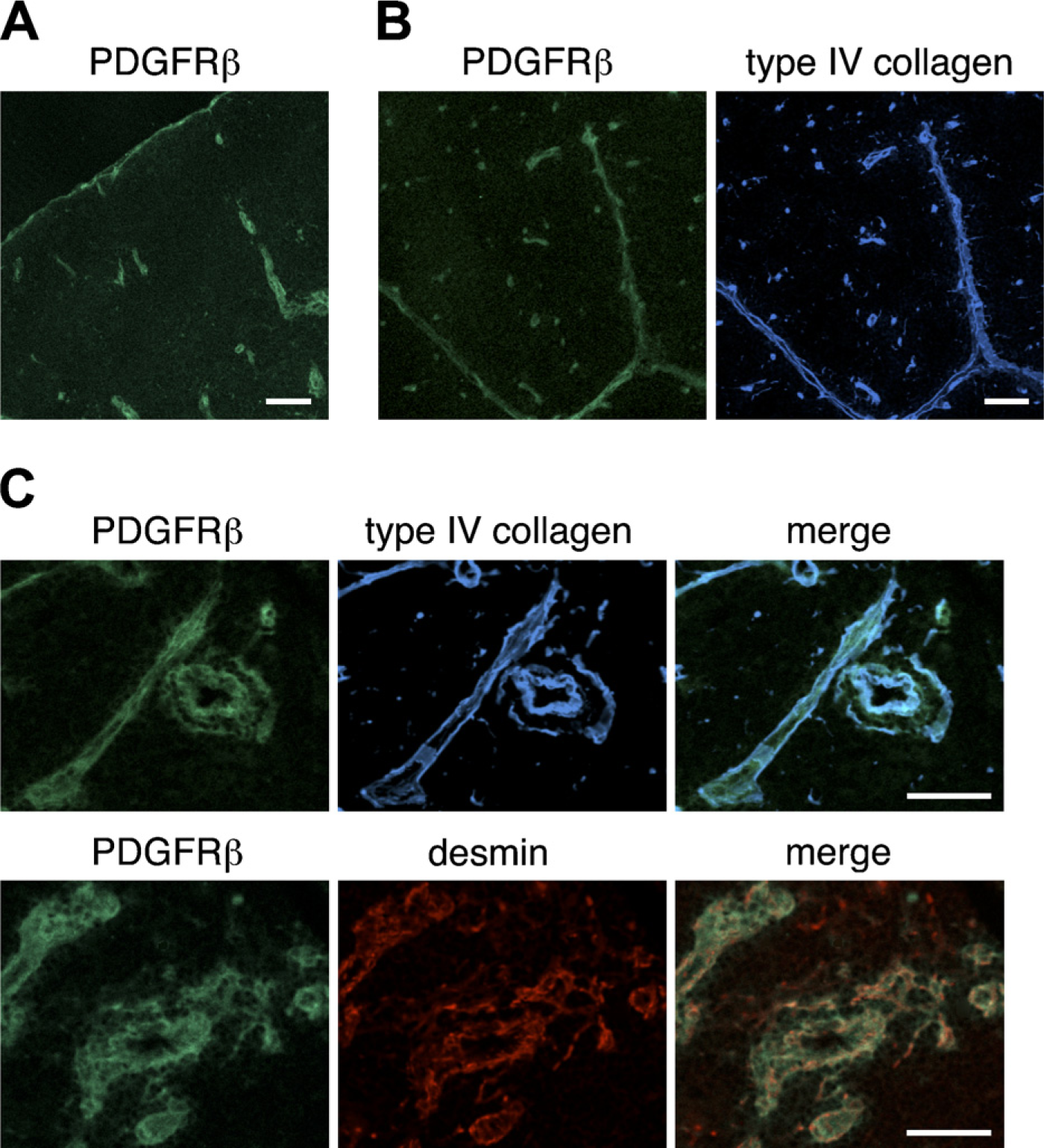
Expression of PDGFRβ in the thymus of adult mouse. Immunofluorescence staining of thymic sections of adult BALB/c thymi was performed to detect PDGFRβ (
Expression of PDGFRα and PDGFRβ
We next assessed the expression of PDGFRα and PDGFRβ in the thymi of BALB/c mice. The expression of PDGFRα was weak; nevertheless, PDGFRα-positive cells were occasionally located in the capsule (Figure 3A) and within intralobular septa (Figure 3B). Some perivascular cells expressed PDGFRα (Figure 3C). Comparison with type IV collagen reactivity indicated that PDGFRα-positive cells were frequently in close association with basement membranes around blood vessels (Figure 3C). The expression of PDGFRβ around thymic vasculatures was intense (Figure 4). Many PDGFRβ-positive cells were observed around almost all blood vessels, including capillaries in the cortical region, and desmin-positive perivascular cells highly expressed PDGFRβ (Figure 4C). PDGFRβ-positive cells were also detected in the capsule and septa (Figures 4A and 4B).
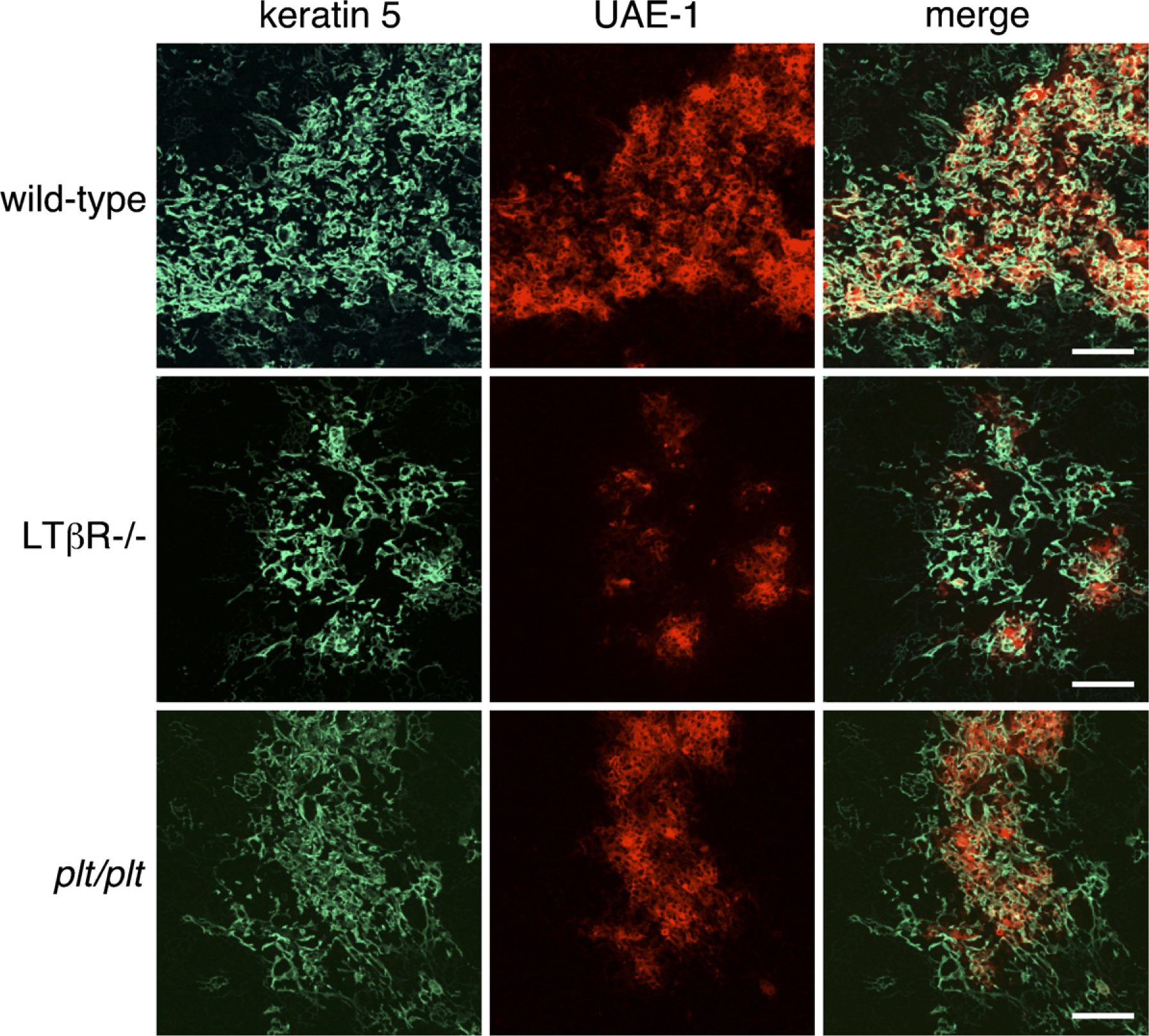
Organization of thymic medullary epithelium in adult C57BL/6 wild-type, lymphotoxin b receptor–negative (LTβR-/-), and plt/plt mice. Immunofluorescence staining of thymic sections of adult C57BL/6 wild-type, LTβR-/-, and plt/plt mice was performed to detect keratin 5 (green) and the binding to Ulex europaeus agglutinin-1 (UEA-1) (red). Aberrant medulla formation and marked reduction in UEA-1 binding to medullary epithelial cells are seen in LTβR-/- mice. Plt/plt thymi tend to have small medullary islets, but the clusters of medullary epithelial cells expressing keratin 5 and binding to UEA-1 are present. Bar = 100 μm.
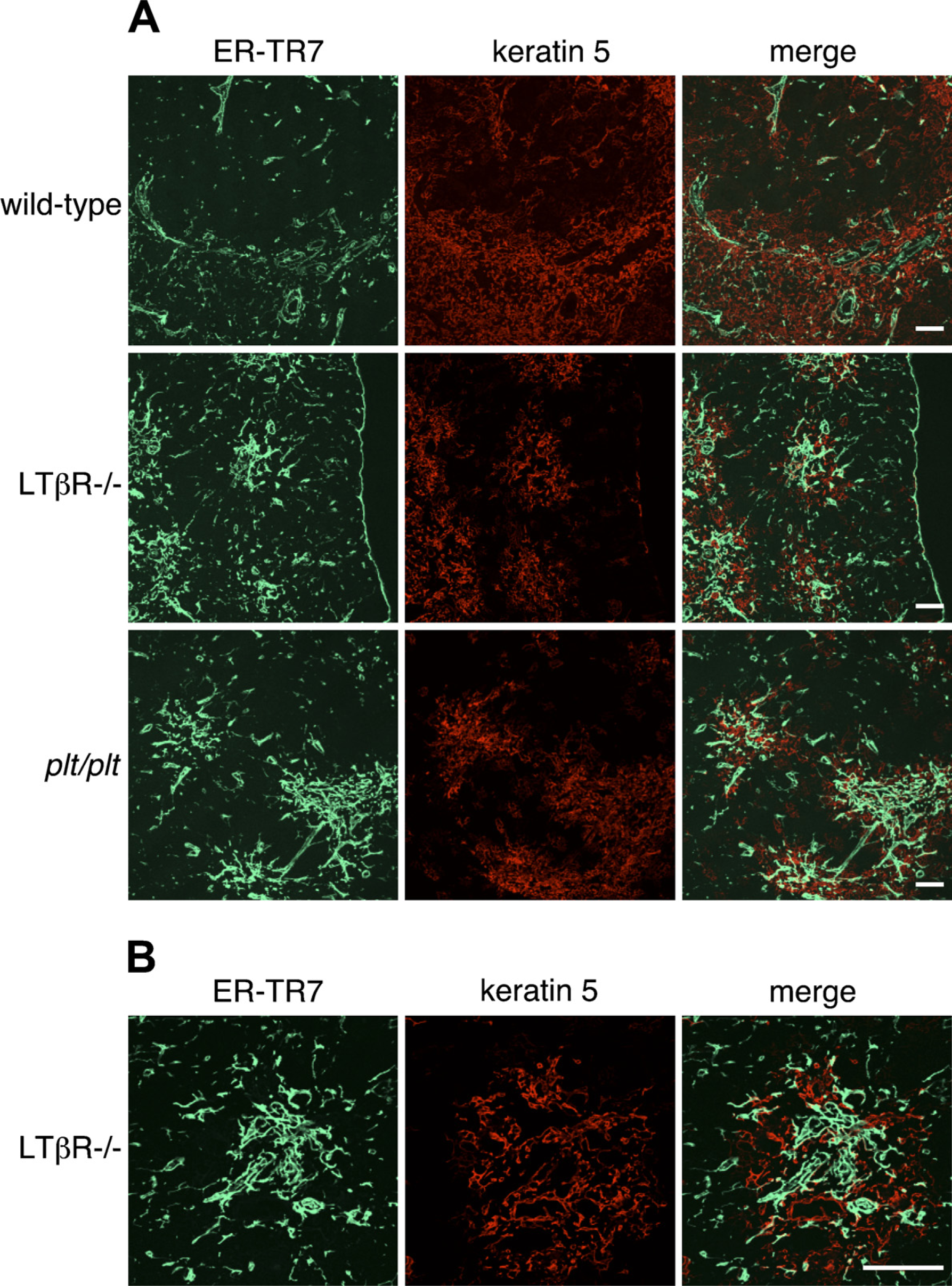
Localization of ER-TR7 and medullary epithelium in the thymi of adult C57BL/6 wild-type, LTβR-/-, and plt/plt mice. (
Localization of Medullar Epithelium and Mesenchyme in the Thymi of LTβR-deficient Mice and plt/plt Mice
We stained thymus sections from C57BL/6 wild-type, LTβR-/-, and plt/plt mice with reagents recognizing thymic medullary epithelium and analyzed them by confocal microscopy. The distribution of keratin 5 allows distinction of medullary epithelial cells, and the mature populations of thymic medullary epithelial cells bind the lectin UEA-1. In wild-type mice, UEA-1–positive epithelial cells formed a network of stellate cells that builds the thymic medulla (Figure 5). As expected, the medullary regions of LTβR-/- thymi were smaller and more sparsely populated by keratin 5–positive medullary epithelial cells than were those of wild-type thymi (Figure 5). UEA-1–positive medullary epithelial cells were detected on LTβR-/- thymic sections at a lower frequency than on wild-type sections, and were localized in small clusters, consistent with previous observations showing that the deficiency of LTβR signaling affects UEA-1–positive medullary epithelial cells (Boehm et al. 2003). Plt/plt thymi tended to have small foci of medullar epithelium, but were not disrupted to the extent of LTβR-/- thymi, and the clusters of medullary epithelial cells expressing keratin 5 and binding UEA-1 were detectable.
We next examined the localization of medullary epithelium and mesenchyme in wild- type, LTβR-/-, and plt/plt thymi by double staining with anti–keratin 5 and ER-TR7. Surprisingly, aberrant foci in labeling with ER-TR7 were evident in the medullary regions of both LTβR-/- and plt/plt thymi (Figure 6). Many ER-TR7–positive cells were situated in epithelial cell– free regions of the medulla as well as perivascular cells, capsule, and septa, and they were found to be frequently adjacent to the epithelial cells, but coexpression of ER-TR7 and keratin 5 was rarely detectable in LTβR-/- and plt/plt thymi. The numbers of ER-TR7–expressing cells in the medulla of LTβR-/- thymi appeared to be higher than those of plt/plt thymi. In addition, we examined the expression of desmin or PDGFRβ in wild-type, LTβR-/-, and plt/plt thymi. In wild-type mice, desmin expression was restricted in perivascular pericytes, capsule, and septa, like BALB/c thymi (data not shown). On the other hand, desmin was also detected in epithelial cell–free regions of the medulla in LTβR-/- mice (Figure 7A). Expression of PDGFRβ was found in the same area (Figure 7B). The labeling patterns of desmin and PDGFRβ were indistinguishable from those of ER-TR7 in plt/plt sections (data not shown). PDGFRα was expressed at low levels in the regions of both LTβR-/- and plt/plt thymi, whereas α-SMA was undetectable in the regions (data not shown). These mesenchymal marker–expressing cells in epithelial cell–free regions of the medulla were not associated with blood vessels, as assessed by CD31 expression (data not shown). Altogether, we conclude that mesenchymal cells are abnormally localized in epithelial cell–free regions of thymic medulla in LTβR-/- and plt/plt mice.
Localization of desmin, PDGFRβ, and medullary epithelium in the thymi of adult LTβR-/- mice. Immunofluorescence staining of thymic sections of adult LTβR-/- mice was performed to detect desmin (
Discussion
The expression of pan-mesenchymal marker ER-TR7, desmin, α-SMA, PDGFRα, and PDGFRβ was observed in the thymus of normal adult mouse by immunofluorescence staining, indicating colocalization of ER-TR7 and desmin along the capsule, intra- and interlobular septa, and around vasculature. Desmin-positive cells were present on almost all segments of thymic CD31-positive blood vessels. The distribution of pericytes or VSMCs in blood vessels varies among the tissues (Hirschi and D'Amore 1996; Bergers and Song 2005). Thymic arterioles identified by the expression of ephrinB2 were completely covered by α-SMA–positive VSMCs. Some large venules were almost covered by VSMCs, whereas some were only partially covered. α-SMA was undetectable in capillaries of adult thymus. This observation is consistent with previous studies, which demonstrated that pericytes on normal capillaries typically express desmin but not α-SMA, whereas normal venules express both molecules (Nehls and Drenckhahn 1993).
In embryos, PDGFRα is expressed on the mesenchymal layer of fetal thymus (Morrison-Graham et al. 1992; Takakura et al. 1997). These mesenchymal cells also express PDGFRβ. In the thymus of adult mouse, the distribution of PDGFRβ was similar to that of desmin, and PDGFRβ was highly expressed on perivascular cells, i.e., pericytes and α-SMA–positive VSMCs (data not shown). Recruitment and coverage of vessels with pericytes is essential for the development of vasculature, as has been shown by genetic ablation of PDGFRβ signaling (Soriano 1994; Hellstrom et al. 1999). Therefore, the presence of PDGFRβ in thymic perivascular cells also might contribute to the yascular formation and morphogenesis. Because ER-TR7 is not a cell surface marker, PDGFRα has been used as a marker for thymic mesenchymal cells instead (Jenkinson et al. 2003, 2007). Although a small population of PDGFRα-positive mesenchymal cells is present in the neonatal and adult thymus (Jenkinson et al. 2007), in adult thymus, the distribution of PDGFRα was different from that of the pan-mesenchymal marker ER-TR7. The expression of PDGFRα in the thymus was weak, and PDGFRα-positive cells were occasionally located in the capsule and within the intralobular septa. Some perivascular cells expressed PDGFRα. Our observation that PDGFRα expression was low in thymic perivascular cells of adult mouse is consistent with other studies showing that neural crest–derived cells within the parenchyma of the thymus downregulate the expression of PDGFRα, whereas they maintain PDGFRβ expression (Foster et al. 2008; Müller et al. 2008). A similar downregulation of PDGFRα has also been observed during pericyte differentiation in the brain (Bondjers et al. 2006). Migration of neural crests into the thymus during embryogenesis and their differentiation into perivascular cells has been demonstrated (Foster et al. 2008; Müller et al. 2008). However, the process through which thymic mesenchymal cells differentiate into perivascular cells during thymus organogenesis remains to be clarified.
In addition, we examined the expression of mesenchymal markers in both LTβR-/- and plt/plt thymi that display the disorganization of medullary epithelium, and observed abnormal distribution of mesenchyme in these thymi. Aberrant foci of mesenchyme were found in epithelial cell–free regions of the thymic medulla. This finding is more prominent in LTβR-/- mice than in plt/plt mice. These results suggest that abnormal medullary architecture may accompany aberrant distribution of mesenchyme in adult thymus. Further studies are necessary in other mice displaying altered disruption of thymus morphology. Epithelial-mesenchymal transition (EMT) is well known as the process whereby cells undergo a switch from epithelial cells into mesenchymal phenotypes (Hay 2005). Up-regulation of Snail or Slug, transcription factors for EMT, and α-SMA were undetectable in the medullary mesenchyme of LTβR-/- and plt/plt thymi by immuno-histology analysis (data not shown), and so far, we do not have evidence of EMT in the thymi of these mice.
In summary, our present study indicates similar localization among pan-mesenchymal marker ER-TR7, desmin, and PDGFRβ in thymus of normal adult mouse. Expression of PDGFRα in the mesenchyme was weak, compared with the other mesenchymal markers. Almost all perivascular cells were detected with ER-TR7, desmin, and PDGFRβ, but PDGFRα was expressed in only some of perivascular cells. In several organs, such as the brain, liver, and kidney, pericytes have been shown to perform specific functions (Bergers and Song 2005), and it is therefore of interest to examine the possibility that thymic perivascular cells have specialized functions. In addition, we show abnormal distribution of thymic mesenchyme in LTβR-/- and plt/plt mice. Further studies are needed to determine the mechanism by which thymic mesenchymal cells take the place of medullary epithelial cells in LTβR-/- and plt/plt mice.
Footnotes
Acknowledgements
We are grateful to Dr. Y-X. Fu, Dr. M. Zhu, and Dr. J. G. Cyster for providing LTβR-/- and plt/plt mice thymi.