
Abstract
Glioblastoma usually recurs after therapy consisting of surgery, radiotherapy, and chemotherapy. Recurrence is at least partly caused by glioblastoma stem cells (GSCs) that are maintained in intratumoral hypoxic peri-arteriolar microenvironments, or niches, in a slowly dividing state that renders GSCs resistant to radiotherapy and chemotherapy. Because the subventricular zone (SVZ) is a major niche for neural stem cells (NSCs) in the brain, we investigated whether GSCs are present in the SVZ at distance from the glioblastoma tumor. We characterized the SVZ of brains of seven glioblastoma patients using fluorescence immunohistochemistry and image analysis. NSCs were identified by CD133 and SOX2 but not CD9 expression, whereas GSCs were positive for all three biomarkers. NSCs were present in all seven samples and GSCs in six out of seven samples. The SVZ in all samples were hypoxic and expressed the same relevant chemokines and their receptors as GSC niches in glioblastoma tumors: stromal-derived factor-1α (SDF-1α), C-X-C receptor type 4 (CXCR4), osteopontin, and CD44. In conclusion, in glioblastoma patients, GSCs are present at distance from the glioblastoma tumor in the SVZ. These findings suggest that GSCs in the SVZ niche are protected against radiotherapy and chemotherapy and protected against surgical resection due to their distant localization and thus may contribute to tumor recurrence after therapy.
Keywords
Introduction
Glioblastoma is the most common malignant and aggressive brain tumor in adults with poor survival of approximately 20 months after diagnosis despite optimal therapy (i.e. surgical resection of the tumor, radiotherapy, temozolomide chemotherapy, and magnetic tumor-treating fields) in the fittest patient population.1,2
Poor patient survival is at least partly caused by the presence of a small fraction of slowly dividing glioblastoma stem cells (GSCs) in the tumor that reside in protective microenvironments called niches. In GSC niches, GSCs are maintained in a slowly dividing or quiescent state that contributes to their therapy-resistance.3–14 We have characterized GSC niches in patient-derived glioblastoma tissue samples and elucidated that GSC niches in glioblastoma tumors are functionally similar to hematopoietic stem cell (HSC) niches in normal human bone marrow. Both niche types are exclusively peri-arteriolar and hypoxic.4,15–18 The chemokines stromal-derived factor-1α (SDF-1α) and osteopontin (OPN) and their receptors C-X-C receptor type 4 (CXCR4) and CD44, respectively, are involved in homing of GSCs and HSCs in their specific niches and maintenance of their stem cell characteristics. Hypoxia in the niches induces stemness of GSCs and HSCs as the transcription factors hypoxia-inducible factor (HIF)-1α and HIF-2α upregulate expression of SDF-1α, CXCR4, OPN, and CD444,18–20 as well as stem cell genes such as prominin-1 (CD133) and sex-determining region Y-box 2 (SOX2).21–25
In mouse models, GSCs do not only home in hypoxic peri-arteriolar GSC niches in the tumors but also in the subventricular zone (SVZ) of the brain, where they are protected from radiotherapy and chemotherapy.26–28 Clinical outcome in patients is worse when glioblastoma tumors are in contact with the SVZ compared with when this is not the case.29–37 Furthermore, glioblastoma recurrence in patients frequently occurs in close proximity of the SVZ. 38
The SVZ is the largest neurogenic niche that harbors neural stem cells (NSCs) and is located at the border of the lateral ventricles in the cerebrum where NSCs divide occasionally to generate differentiated neurons, astrocytes, and oligodendrocytes.28,39,40 Mouse and human SVZ has been described to consist of three layers. The first layer consists of a monolayer of ependymal cells (ECs) that contain microvilli and separates the SVZ from the ventricles.28,41,42 The ependymal layer is important for transport of small molecules between the cerebrospinal fluid (CSF) and neuropil and forms a barrier between CSF and the brain parenchyma.28,42,43 The second layer is called the hypocellular gap consisting of ependymal and astrocytic cell processes and is hypothesized to be involved in the exchange of signals to establish metabolic homeostasis and to control NSC proliferation and differentiation. The third layer is a ribbon of glial fibrillary acidic protein (GFAP)-positive astrocyte-like neural progenitor cells28,42–44 and CD133-positive NSCs. The next layer is the transitional zone that consists of myelinated axons and oligodendrocytes.28,42,43
GSCs have been shown to home in the SVZ of the brain in mouse models where they are maintained as slowly dividing cells.26,27,45 Homing of GSCs in the SVZ region occurs via the SDF-1α–CXCR4 axis. CXCR4-positive GSCs are attracted to the SVZ where levels of SDF-1α are high. In the SVZ, GSCs are protected from radiotherapy.26,27,45 When CXCR4–SDF-1α interactions are disrupted by the use of the CXCR4 inhibitor plerixafor, GSCs are able to migrate out of the niches, differentiate, and proliferate and are sensitized to radiotherapy in mouse models.26,27,45 In a single-arm phase I/II clinical trial, the CXCR4 inhibitor plerixafor was tested in combination with chemotherapy and radiotherapy in 29 glioblastoma patients, which yielded a median overall survival of 21.3 months.46,47 A phase II clinical trial is currently recruiting patients to evaluate the efficacy of plerixafor in combination with whole-brain radiotherapy and temozolomide chemotherapy (ClinicalTrials.gov Identifier: NCT03746080). The primary endpoint is 6-month progression-free survival.
OPN in the SVZ acts as a chemokine 48 for CD44-positive NSCs49,50 and is involved in survival of NSCs. Thus, the OPN–CD44 axis may also play a role in homing of GSCs in the SVZ.
As far as we know, the SVZ of glioblastoma patients has not yet been investigated with respect to the presence of GSCs. Because GSC niches in glioblastoma tumors are likely to be removed by surgery, the aim of this study was to investigate whether GSCs are also hiding in the protective SVZ at distance from the tumor mass and thus escape from removal by surgery.
To address this aim of the study, a thorough characterization of the SVZ of human glioblastoma patients was performed to understand how and where GSCs are localized in the SVZ, how to distinguish NSCs from GSCs, and to determine which cell types, molecules, and tissue structures are involved in the SVZ in the maintenance of stemness of NSCs and GSCs. GSCs express stem cell markers CD133 and SOX2.15–17,51–55 We demonstrated in our previous study that the CD9 gene that encodes for tetraspanin, a cell surface glycoprotein, is expressed on GSCs derived from patient biopsies, whereas CD9 expression was not found on NSCs, astrocytes, and in normal brain. 56 We aimed to test the hypothesis that the SVZ as NSC niche is morphologically similar to hypoxic peri-arteriolar GSC niches in glioblastoma and that GSCs are present in the SVZ of brains of glioblastoma patients.
Materials and Methods
Cell Cultures
Human NCH421K and NCH644 GSCs56,57 were a generous gift from Prof. Christel Herold-Mende (Heidelberg University, Heidelberg, Germany) and K26 GSCs were isolated and propagated from a patient-derived glioblastoma sample and were cultured as 3D tumor spheres in Neurobasal medium (Gibco Life Technologies, Carlsbad, CA, USA) containing 1% penicillin/streptomycin (Sigma, St. Louis, MO, USA), 1% L-glutamine (Sigma), 2% B27, 0.08% basic fibroblast growth factor (bFGF; Gibco), 0.01% epidermal growth factor (EGF; Gibco) and 0.01% heparin (Sigma) at 37C in a 5% CO2 incubator. U87 and U373 differentiated glioblastoma cells (ATTC, Teddington, Middlesex, UK) were cultured in Dulbecco’s modified Eagle’s medium (DMEM) medium (Sigma) containing 1% penicillin/streptomycin, 1% L-glutamine, and 10% fetal bovine serum (Gibco) at 37C in a 5% CO2 incubator. The U87 glioblastoma cells were authenticated as described by Torsvik et al. 58
Immunocytochemistry on Human GSCs and Differentiated Glioblastoma Cells
Human U87 glioblastoma and U373 cells were cultured on glass slides in an incubator at 37C. On each glass slide, 500,000 cells were seeded. Two days after U87 and U373 cells had been seeded on glass slides, cells were fixed in 4% paraformaldehyde (Merck, Darmstadt, Germany) for 10 min, followed by a washing step using phosphate-buffered saline (PBS; Gibco) containing 1% bovine serum albumin (BSA; Sigma-Aldrich). Cells were permeabilized and nonspecific background staining was reduced by incubating cell preparations in PBS containing 1% BSA, 10% normal goat serum (Dako Glostrup, Denmark), and 0.1% Triton-X (Sigma-Aldrich) for 1 hr at room temperature (RT). Cells were incubated overnight at 4C with primary antibodies diluted in PBS containing 1% BSA, as indicated in Table 1.
Details of Primary Antibodies Used for Fluorescence Immunohistochemistry.
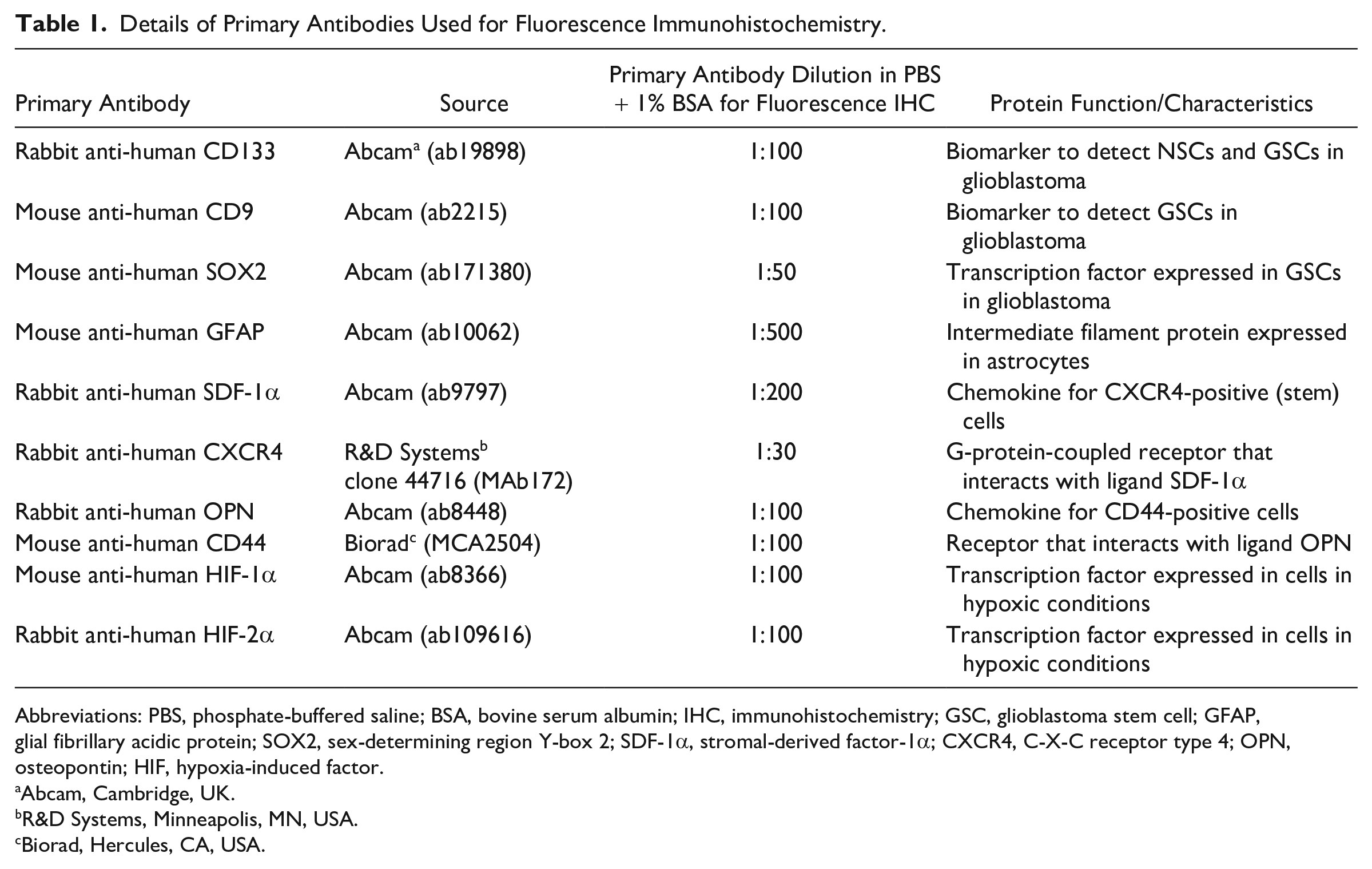
Abbreviations: PBS, phosphate-buffered saline; BSA, bovine serum albumin; IHC, immunohistochemistry; GSC, glioblastoma stem cell; GFAP, glial fibrillary acidic protein; SOX2, sex-determining region Y-box 2; SDF-1α, stromal-derived factor-1α; CXCR4, C-X-C receptor type 4; OPN, osteopontin; HIF, hypoxia-induced factor.
Abcam, Cambridge, UK.
R&D Systems, Minneapolis, MN, USA.
Biorad, Hercules, CA, USA.
After 5 days of culture, tumor spheres of approximately 106 NCH421K, NCH644, and K26 GSCs were placed in eppendorf tubes and the entire immunostaining procedure was performed in the eppendorf tubes for the GSCs to maintain the three-dimensional (3D) structure of the tumor spheres as described before. 16 GSC tumor spheres were fixed in the eppendorf tubes in 4% paraformaldehyde (Merck) for 10 min, followed by a washing step using PBS containing 1% BSA. Cells were permeabilized and nonspecific background staining was reduced by incubating cell preparations in PBS containing 1% BSA, 10% normal goat serum (Dako Glostrup, Denmark), and 0.1% Triton-X (Sigma-Aldrich) for 1 hr at RT. Cells were incubated overnight at 4C with primary antibodies diluted in PBS containing 1% BSA, as indicated in Table 1.
Alexa Fluor 488-conjugated goat anti-rabbit antibodies (Life Technologies, Carlsbad, CA, USA) and Alexa Fluor 546-conjugated goat anti-mouse antibodies (Thermo Fisher Scientific, Waltham, MA, USA) were used as secondary antibodies in a dilution of 1:200 in PBS containing 1% BSA as described before. 16 Incubation lasted 1 hr at RT. Cells were washed in PBS, followed by nuclear counterstaining using 4′,6-diamidino-2-phenylindole (DAPI; Sigma-Aldrich) for 5 min at RT. Cells were washed in PBS for 5 min and coverslipped using Prolong Gold mounting medium (Life Technologies). Control incubations were performed in the absence of primary antibodies.
SVZ-invaded Glioblastoma Patient Samples
Brain tissue material was obtained from seven patients (aged 27–75 years at the time of glioblastoma diagnosis: six IDH1-wild type and one IDH1-mutated glioblastoma at autopsy at the Department of (Neuro)Pathology of Amsterdam UMC, Academic Medical Center, University of Amsterdam, The Netherlands). Informed consent was obtained for the use of brain tissue for research purposes. All procedures performed using human brain tissue were in accordance with the ethical standards of the Amsterdam University Medical Center (location AMC, reference number W14_224 # 14.17.0286), local protocols, the 1964 Helsinki declaration and its later amendments, or comparable ethical standards. Human brain tissue was fixed in 10% buffered formalin and embedded in paraffin. Histological diagnosis was confirmed following the current World Health Organization classification guidelines 2 by two independent neuropathologists. Sections of paraffin-embedded tissue (including the SVZ region from the site of tumor localization) were cut (5 µm thick), mounted on pre-coated glass slides (Star Frost, Waldemar Knittel, Braunschweig, Germany) and processed for immunohistochemical staining. Consent for removal of the tissue and its storage in the tumor bank for research purposes was obtained and documented in the medical charts of the patients. Patient characteristics are shown in Table 2. The seven glioblastoma samples were from patients who were treated before the advent of molecular characterization as standard practice in glioblastoma care. Therefore, the samples were not genetically fully characterized and thus next generation sequencing data of these samples was not available.
Glioblastoma Patient Characteristics.
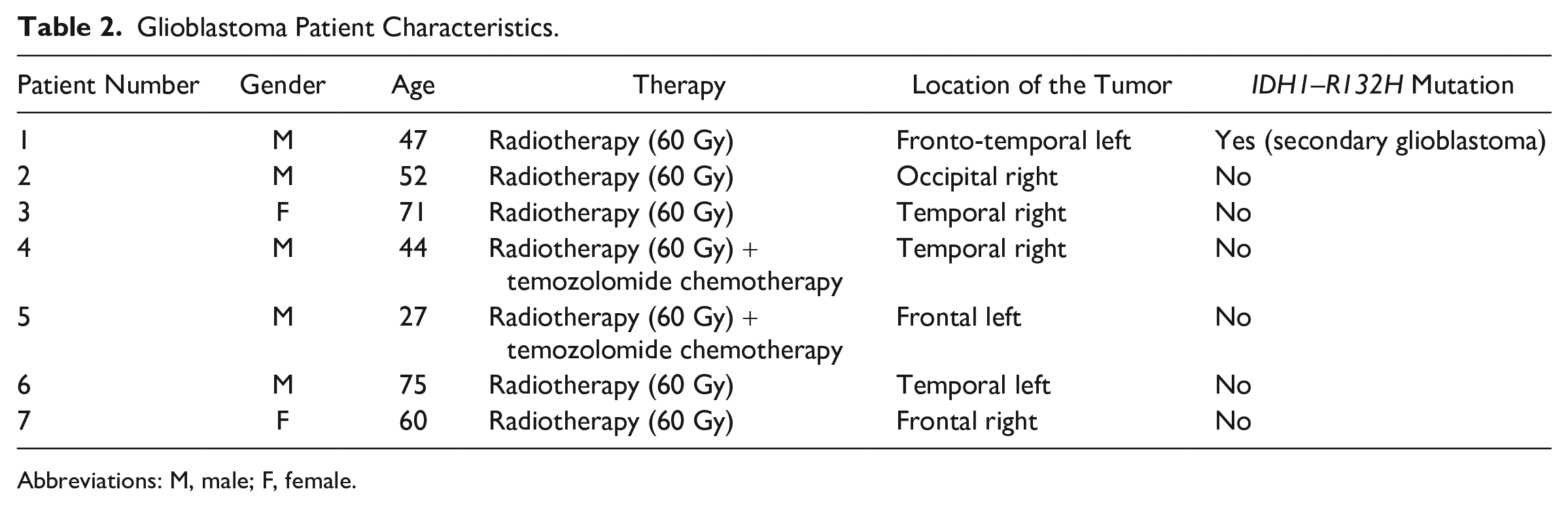
Abbreviations: M, male; F, female.
Fluorescence Immunohistochemistry Using Human SVZ-invaded Glioblastoma Paraffin Sections
Paraffin sections (5 µm thick) of human SVZ-invaded glioblastoma were stored at RT until use. Dewaxing was performed by incubation of the sections in xylene (VWR Chemicals, Atlanta, GA, USA) for 5 min and rinsing in 100%, 96%, and 70% ethanol (Merck), respectively.
Antigen-retrieval was performed in a microwave using 100 mM citrate buffer containing 0.1% Triton-X (Sigma-Aldrich), pH 6.0, for 20 min at 98C, followed by cooling down for 20 min and a washing step in PBS. 59
Sections were encircled with a PAP pen (Dako) and incubated with PBS (Gibco) containing 10% normal goat serum (Dako) and 0.1% Triton-X for 1 hr at RT to reduce nonspecific background staining and for permeabilization of the sections. Sections were subsequently incubated overnight at 4C with primary antibodies diluted in PBS containing 1% BSA (Sigma-Aldrich), as indicated in Table 1. Sections were washed three times using PBS containing 1% BSA.
Alexa Fluor 488-conjugated goat anti-rabbit antibodies (Life Technologies) and Alexa Fluor 546-conjugated goat anti-mouse antibodies (Thermo Fisher Scientific) were used as secondary antibodies in a dilution of 1:200 in PBS containing 1% BSA for 1 hr at RT. Sections were then washed in PBS, followed by nuclear counterstaining using DAPI (Sigma-Aldrich) in PBS for 5 min at RT. Next, sections were washed in PBS for 5 min and coverslipped using Prolong Gold mounting medium (Life Technologies). Afterward, sections were sealed with nail polish and dried overnight. Control incubations were performed in the absence of primary antibodies.
Hematoxylin and Eosin (HE)
For HE staining, human SVZ-invaded glioblastoma paraffin sections (5 µm) were dewaxed in xylene and 100% ethanol. Sections were fixed with freshly prepared Formol-Macrodex (4% formaldehyde, 7.2 mM CaCl2, 0.12 M Dextran-70, 0.12 M NaCl, and 7.96 mM CaCO3) for 10 min, followed by a washing step in distilled water for 5 min. Nuclei were stained with hematoxylin (Sigma-Aldrich) for 30 sec and then, sections were placed in running tap water for 5 min, after which the sections were placed in distilled water. Sections were then stained with eosin (Merck) for 20 sec, dipped five times in distilled water, 15 times in 70% ethanol, 15 times in 96% ethanol, and 10 times in 100% ethanol. Afterward, sections were rinsed three times for 5 min in xylene. The sections were covered with Pertex (Histolab, Götenburg, Sweden). All steps were performed at RT.
Periodic Acid Schiff (PAS) Staining
PAS staining was performed by the Pathology Department at the Amsterdam UMC. Human SVZ-invaded glioblastoma paraffin sections (5 µm) were used and the staining procedure was performed automatically using an Artisan Link Pro machine (Dako, Glostrup, Denmark) and staining kits were purchased from Agilent (Santa Clara, CA, USA).
Imaging
Fluorescence imaging as well as light microscopy for the histochemical staining was performed using an Eclipse Ti-E inverted microscope and NIS-Elements AR 4.13.04 software (Nikon Instruments, Melville, NY, USA). Fluorescence staining patterns of the SVZ-glioblastoma sections and glioblastoma cells were analyzed by three independent observers (V.V.V.H., B.B., C.J.F.V.N.). Morphology and histological characteristics of the SVZ were analyzed by two independent observers on the basis of light microscopical images (V.V.V.H., C.J.F.V.N.). Fluorescence staining of GSCs in 3D tumor spheres was imaged using a confocal microscope (Leica DFC 7000 T, Wetzlar, Germany). Staining patterns were analyzed by three independent observers (V.V.V.H., B.B., C.J.F.V.N.).
The total number of NSCs and GSCs were quantified in the seven samples using image analysis and Image J software. 60 Three images per patient sample were analyzed. NSCs were quantified by the presence of CD133 and SOX2 expression. GSCs were detected and quantified by the presence of CD133 and CD9 expression. GFAP-positive cells were also quantified in the seven SVZ samples.
Results
CD133 and SOX2 as GSC and NSC Biomarkers and CD9 as Selective GSC Biomarker
To determine which biomarkers can be used to detect GSCs in glioblastoma tissue samples, immunocytochemical analysis was performed on cell lines (Fig. 1). Monolayers of U87 and U373 differentiated glioblastoma cell lines and spheroids of NCH421K, NCH644, and K26 GSC lines were characterized in vitro using immunocytochemical analysis. Monolayers of U87 differentiated glioblastoma cells did not express the stem cell biomarkers CD133 on the surface and SOX2 in the nuclei (Fig. 1A), but expressed the astrocyte marker GFAP (Fig. 1B) and CD9 (Fig. 1C). The expression patterns of U373 were similar (data not shown). In all tested conditions described above and throughout the “Results” section, control staining in the absence of the primary antibodies was negative (Supplementary Fig. 1).
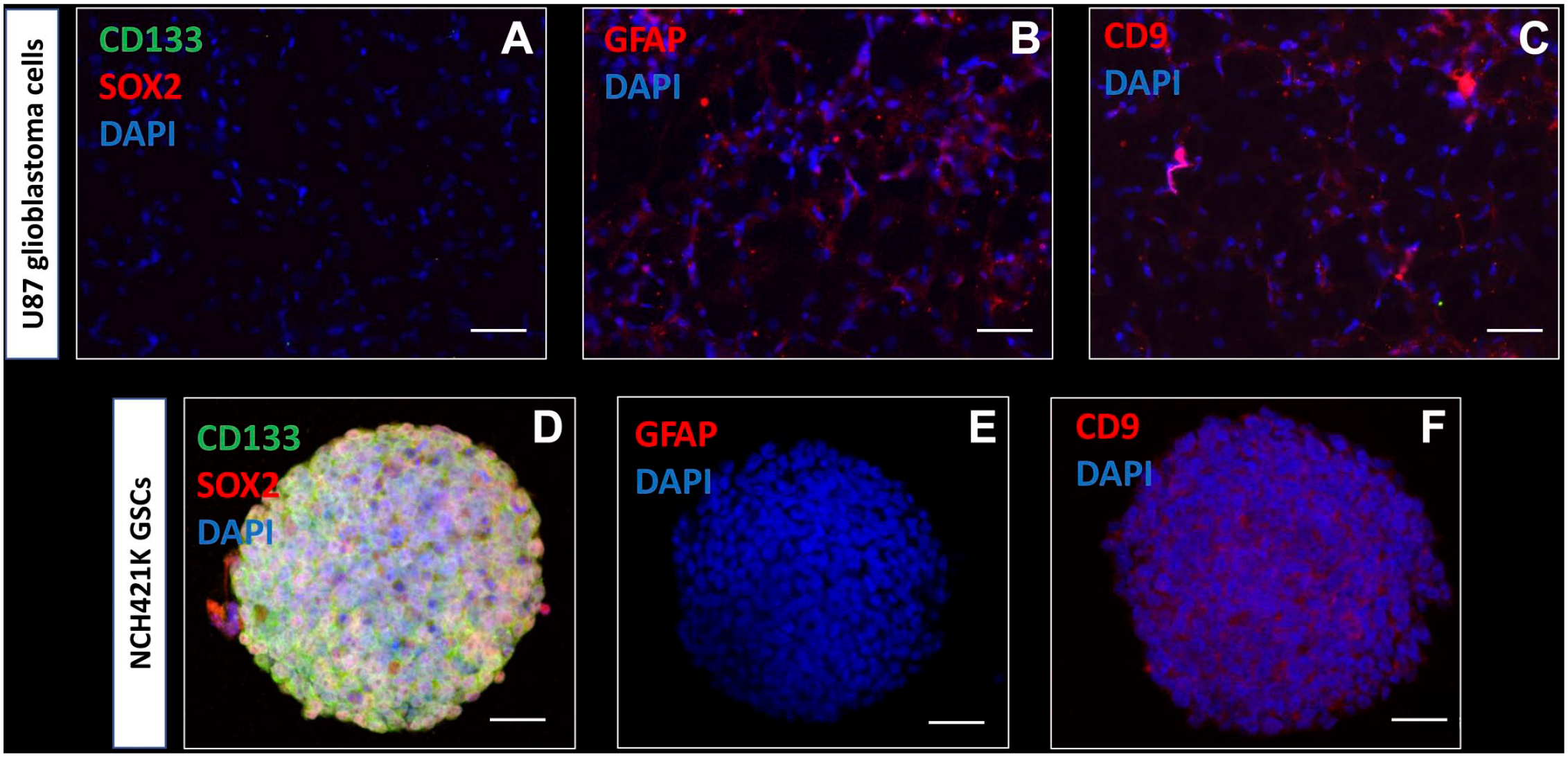
Immunocytochemical characterization of U87 differentiated glioblastoma cells and NCH421K GSCs. Wide-field fluorescence microscopy reveals that monolayered U87 glioblastoma cells do not express the stem cell biomarkers CD133 (green) and SOX2 (red) (A), express the astrocyte marker GFAP (B) and cell surface biomarker tetraspanin CD9 (C). Confocal microscopy demonstrates that NCH421K GSCs express the stem cell biomarkers CD133 (green) and SOX2 (red) in vitro in tumor spheres (D). GFAP is not expressed in the NCH421K GSCs in vitro in tumor spheres (E). CD9 cell surface expression is detected in NCH421K GSCs in vitro in tumor spheres (F). DAPI is used for nuclear counterstaining. Abbreviations: DAPI, 4′,6-diamidino-2-phenylindole; GFAP, glial fibrillary acidic protein; GSC, glioblastoma stem cell; SOX2, sex-determining region Y-box 2. Scale bar A–C = 100 µm; scale bar D–F = 50 µm.
Confocal microscopy of 3D tumor spheres of NCH421K GSCs showed that stem cell biomarkers CD133 and SOX2 were expressed on the surface and in the nuclei, respectively (Fig. 1D). The astrocyte biomarker GFAP was not expressed in GSCs (Fig. 1E). GSCs expressed biomarker CD9 on the surface (Fig. 1F). The expression patterns of NCH644 GSCs and K26 GSCs were similar (data not shown).
On the basis of the immunocytochemical data, CD133 and SOX2 were used for the detection of GSCs and NSCs in the SVZ of glioblastoma tissue sections. In addition, CD9 was used to discriminate between CD9-positive GSCs and normal NSCs, as NSCs and normal brain tissue have been shown to be CD9-negative in our previous study. 56
Morphological Characteristics of the SVZ of Glioblastoma Patients
To assess the morphological characteristics of the SVZ of glioblastoma patient samples, HE (Fig. 2A) and PAS (Fig. 2B) staining was performed. Figure 2 shows representative images of the morphology found in the seven SVZ samples at distance of the tumor of glioblastoma patients.
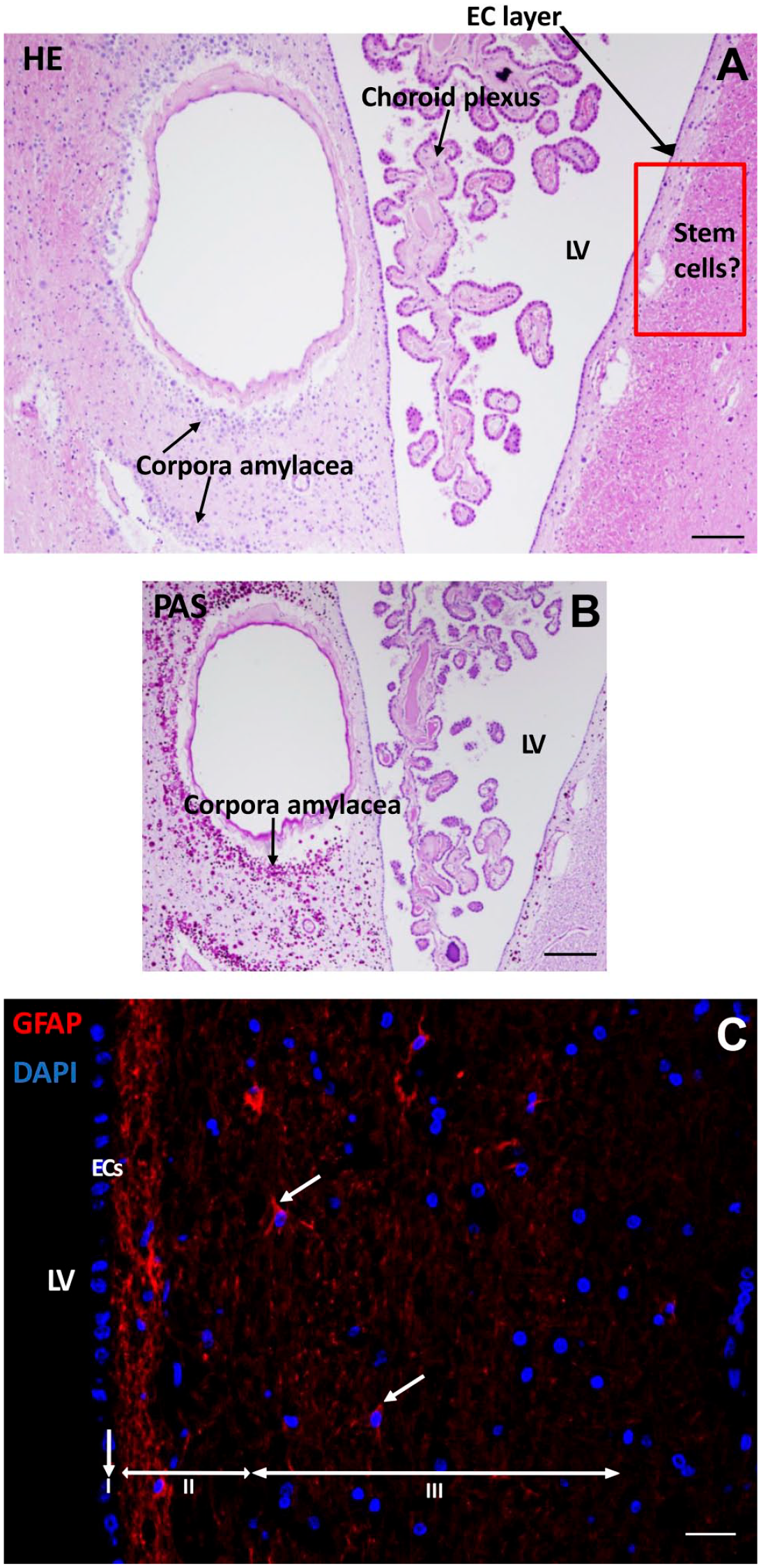
Morphological characteristics of the SVZ of a glioblastoma patient. HE staining shows the LV with choroid plexus inside the ventricle where CSF is produced with an EC layer lining the LV. Along the LV, corpora amylacea structures are found. The red rectangled area indicates the possible location of NSCs and GSCs (A). PAS staining demonstrates the corpora amylacea structures in the SVZ (B). Immunofluorescence staining of GFAP shows the presence of neural progenitor cells in the SVZ in red (white arrows). The three top layers of the SVZ are indicated: the monolayer of ECs (layer I), the hypocellular gap that contains EC and astrocyte processes (layer II), and the layer of NSCs and astrocytic-like neural progenitor cells that express GFAP (layer III). DAPI is used for nuclear counterstaining (C). Scale bars A, B = 100 µm, scale bar C = 50 µm. Abbreviations: DAPI, 4′,6-diamidino-2-phenylindole; EC, ependymal cell; HE, hematoxylin and eosin; LV, lateral ventricle; CSF, cerebrospinal fluid; GFAP, glial fibrillary acidic protein; NSC, neural stem cell; PAS, periodic acid–Schiff; SVZ, subventricular zone.
In the lateral ventricle, the choroid plexus was present that produces the CSF. Corpora amylacea were present in the SVZ of the glioblastoma patients (Fig. 2B). The numbers of corpora amylacea structures varied between the patient samples, but were found in all seven samples. The exact nature of corpora amylacea is not known but they accumulate in the human brain during aging and in neurodegenerative disorders and likely contain non-degradable waste products. 61
The SVZ was found to be characterized by a monolayer of ECs (Fig. 2A–C) that separates the SVZ from the ventricles (Fig. 2C). GFAP staining showed the EC and astrocytic processes in the hypocellular gap (second layer) and the astrocytic-like neural progenitor cells and astrocytes in the third layer of the SVZ (Fig. 2C). All seven SVZs showed a similar morphological pattern. However, the width of the hypocellular gap varied among patient samples.
NSCs in the SVZ Express CD133 and SOX2
In the seven samples, NSCs were detected in the SVZ. NSCs expressed biomarker CD133 in the cytoplasm and on the plasma membrane (Fig. 3A) and approximately half of the CD133-positive NSCs co-expressed transcription factor SOX2 in the nuclei (Fig. 3B; Table 3). Both CD133 and SOX2 were also found to be highly expressed in the EC layer of the SVZ (Fig. 3). The ECs showed differences in cellular morphology that are most likely related to their activity. In Fig. 3A, the ECs are columnar and highly active whereas in Fig. 3B, the ECs are cuboid and more quiescent. 62
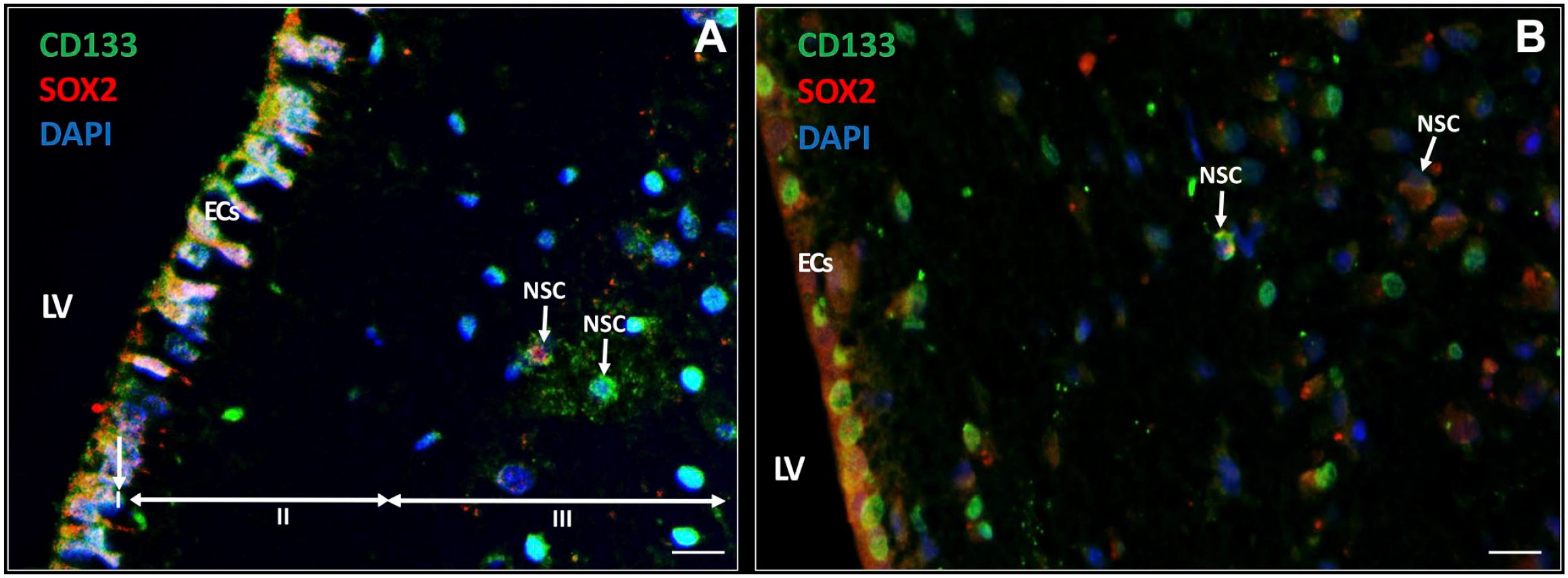
Immunofluorescence staining of NSCs in the SVZ of glioblastoma patients. NSCs (white arrows) express CD133 (green) and approximately half of the CD133-positive NSCs co-express SOX2 (red) in the SVZ. ECs also express CD133 and SOX2 (A, B). The ECs in (A) show characteristics of cellular activity, whereas the ECs in (B) are more quiescent. The three top layers of the SVZ are indicated: the monolayer of ECs (layer I), the hypocellular gap that contains EC and astrocyte processes (layer II), and the layer of NSCs and astrocyte-like neural progenitor cells (layer III). DAPI is used for nuclear counterstaining. Scale bar = 100 µm. Abbreviations: DAPI, 4′,6-diamidino-2-phenylindole; EC, ependymal cell; LV, lateral ventricle; NSC, neural stem cell; SOX2, sex-determining region Y-box 2; SVZ, subventricular zone.
Numbers of GSCs and NSCs in the SVZ at Distance of the Tumor of Glioblastoma Patients.
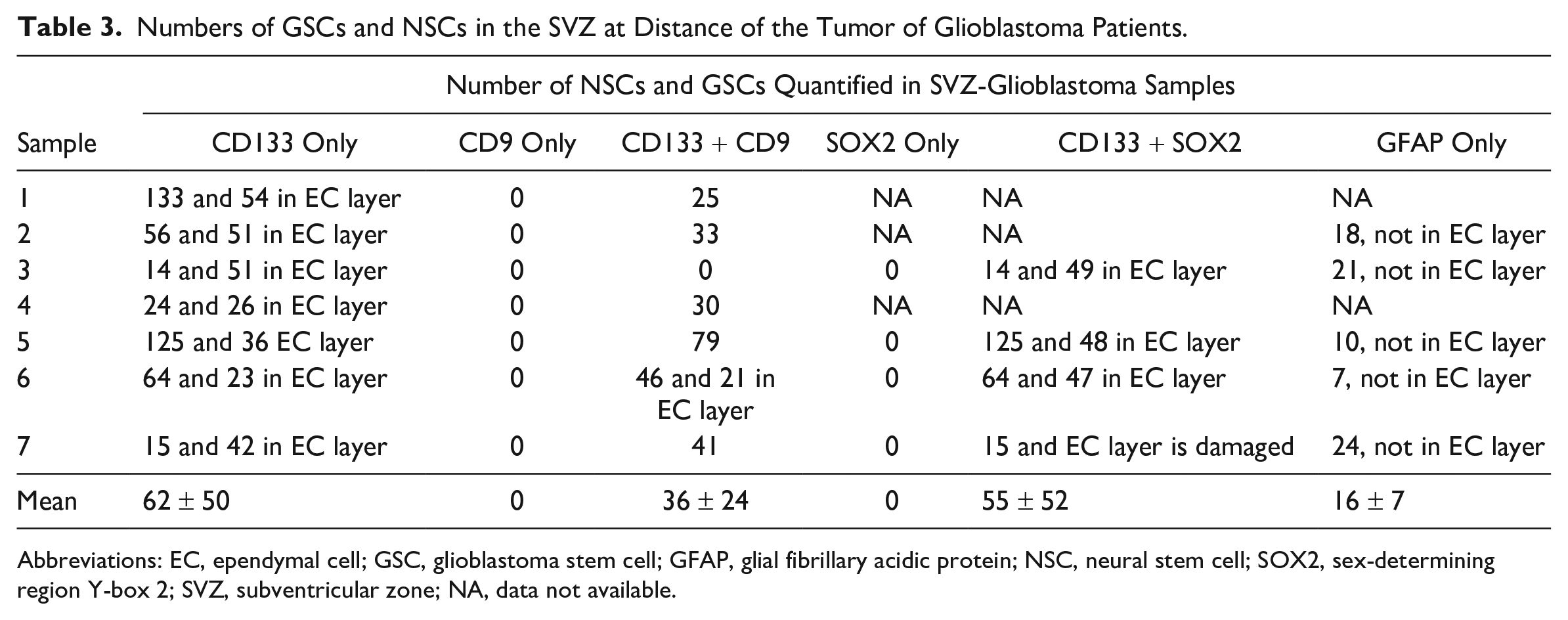
Abbreviations: EC, ependymal cell; GSC, glioblastoma stem cell; GFAP, glial fibrillary acidic protein; NSC, neural stem cell; SOX2, sex-determining region Y-box 2; SVZ, subventricular zone; NA, data not available.
NSCs and GSCs Can Be Distinguished in the SVZ
Next, we determined whether GSCs were present in the SVZ samples of glioblastoma patients. GSCs were detected using the biomarkers CD133, SOX2, and CD9, as GSCs expressed all three biomarkers in vitro (Fig. 1D and F). In the SVZ samples, NSCs did not express the biomarker CD9 (Fig. 4A), whereas GSCs expressed CD133 and CD9 on the plasma membrane (Fig. 4B and C). Thus, with the use of CD133 and CD9, NSCs and GSCs were distinguished in patient tissue samples. NSCs were found in all seven samples and GSCs were found in six out of seven patient samples. The number of NSCs and GSCs were quantified using image analysis (Table 3). The data show that the number of GSCs and NSCs varied among the patient samples. CD9 and SOX2 expression was always found in combination with CD133 expression. In addition, CD133 and SOX2 were abundantly expressed in the EC layer, whereas CD9 expression was found in the EC layer of only 1 patient sample. The number of GFAP-positive cells varied among the samples and was never found to be expressed in the EC layer.
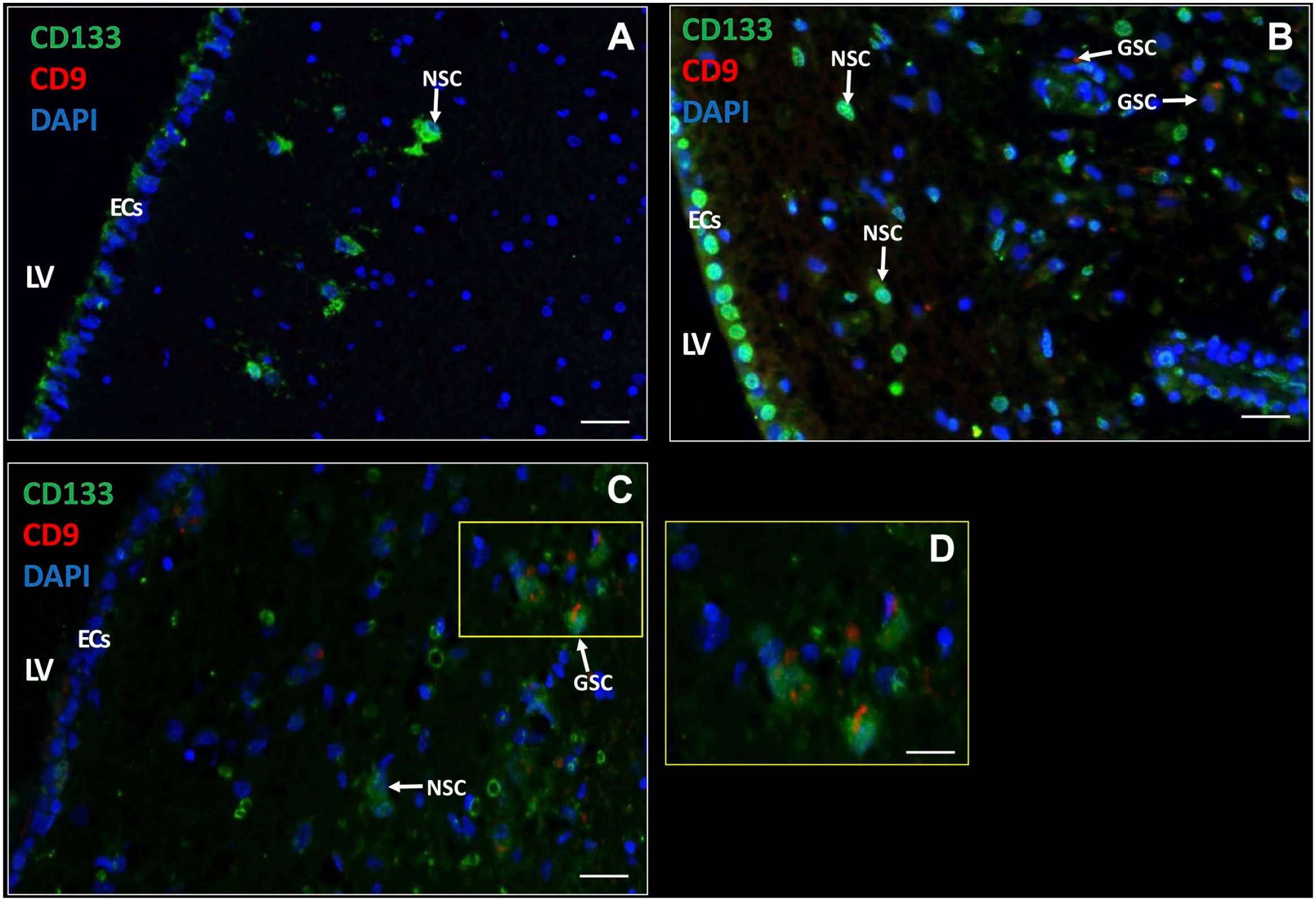
Immunofluorescence staining of NSCs and GSCs in the SVZ of glioblastoma patients. CD133 and CD9 expression in the SVZ of glioblastoma patients (A–D) with active ECs (A, C) and quiescent ECs (B) with variable CD133 expression (medium in A, high in B, and low in C). NSCs are characterized by CD133-positive CD9-negative and GSCs by CD133-positive CD9-positive. D is a magnification of section in C indicated by a yellow rectangle. DAPI is used for nuclear counterstaining. Scale bars: A–C = 100 µm; D = 50 µm. Abbreviations: DAPI, 4′,6-diamidino-2-phenylindole; EC, ependymal cell; LV, lateral ventricle; NSC, neural stem cell. SVZ, subventricular zone.
Chemokines and Their Receptors in the SVZ
Expression of chemokines and their receptors was determined in the SVZ. The chemokine SDF-1α was found to be highly expressed extracellularly as well as intracellularly (Fig. 5A and B). In addition, SDF-1α was also expressed intracellularly in ECs (Fig. 5C). The receptor CXCR4 was found to be expressed on the cell surface and/or nuclei of many cell types in the SVZ, including ECs in an active state (Fig. 5A) and in a quiescent state (Fig. 5B).
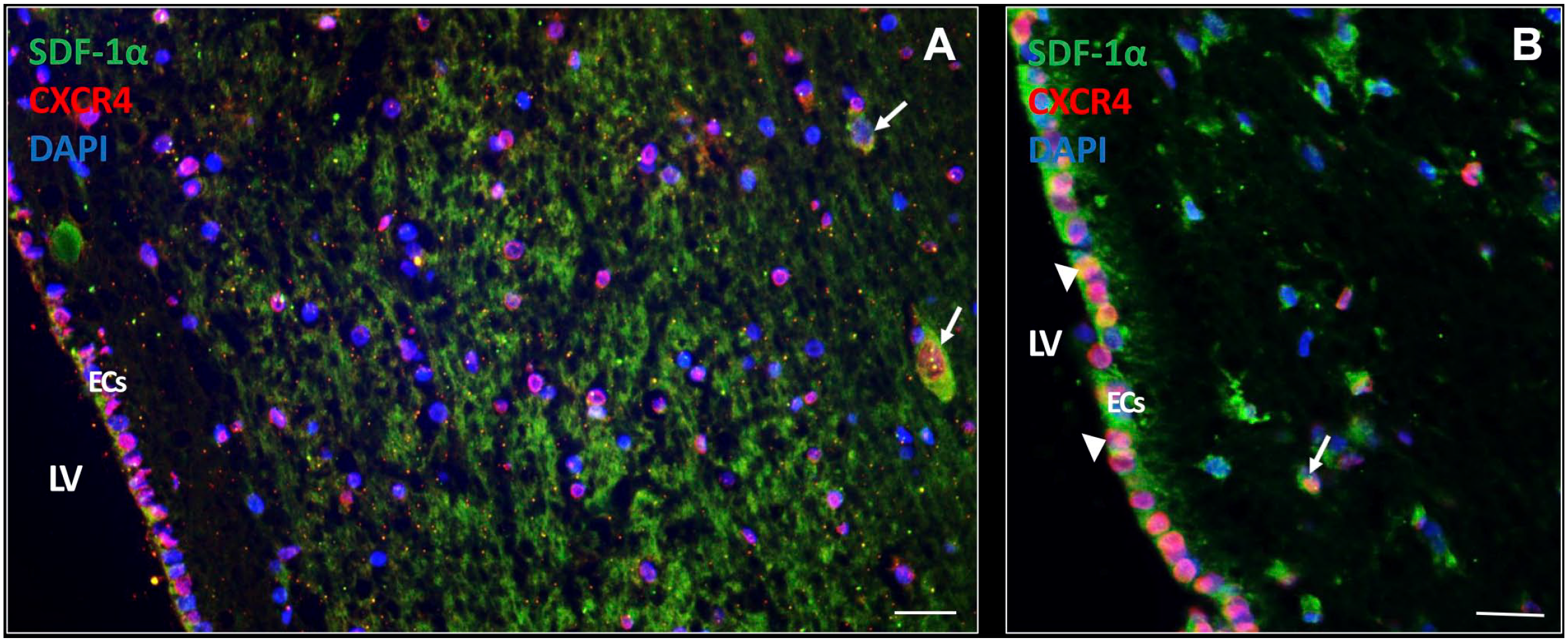
SDF-1α and CXCR4 expression in the SVZ of glioblastoma patients. In the SVZ of glioblastoma patient samples, expression of the chemokine SDF-1α is found abundantly expressed both extracellularly and intracellularly (white arrows). CXCR4 expression is present in various cell types in the SVZ, both in the cytoplasm and nucleus CXCR4 expression (A, B). SDF-1α and CXCR4 protein expression is also found in ECs of the SVZ both in an active state (A) and a more quiescent state (white arrow heads) (B). DAPI is used for nuclear counterstaining. Scale bar A, B = 100 µm. Scale bar C = 200 µm. Abbreviations: CXCR4, C-X-C receptor type 4; EC, ependymal cell; DAPI, 4’,6-diamidino-2-phenylindole; LV, lateral ventricle; SDF-1α, stromal-derived factor-1α; SVZ, subventricular zone.
We also investigated the expression of chemokine OPN, which was highly expressed intracellularly (Fig. 6A) and extracellularly (Fig. 6B) in the patient samples. Receptor CD44 was expressed on a subset of cells in the SVZ and also on ECs (Fig. 6). These expression patterns were found in all samples investigated.
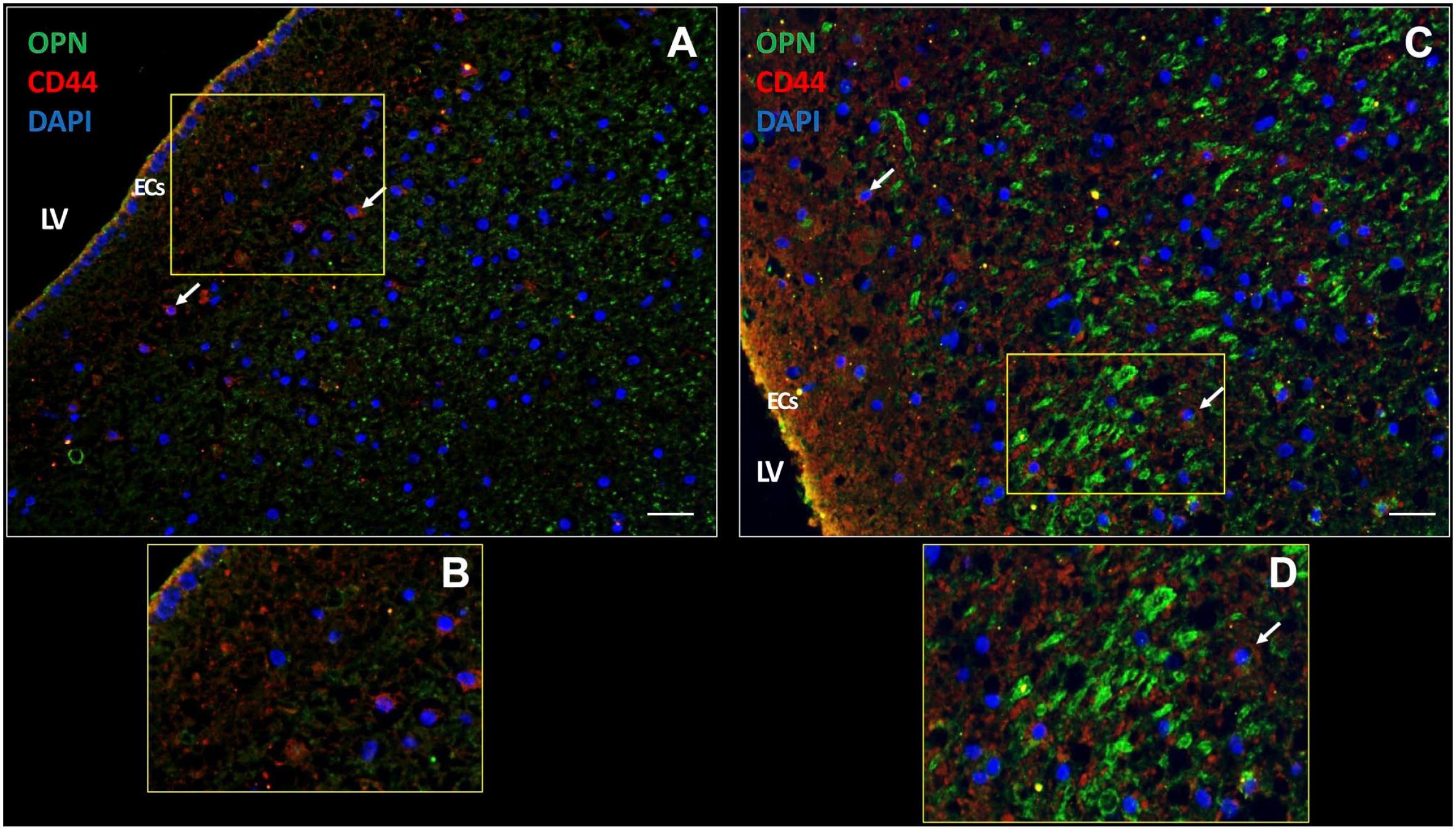
OPN and CD44 expression in the SVZ of a glioblastoma patient. In the SVZ, chemokine OPN is expressed extracellularly (A) as well as intracellularly (yellow dots) (C). CD44 receptor expression is present on the plasma membrane of cells in the SVZ (white arrows) (A, C). OPN and CD44 are expressed in ECs of the SVZ (yellow; C, D). B and D are magnifications of sections in A and C, respectively. DAPI is used for nuclear counterstaining. Scale bars: A = 100 µm; B = 200 µm. Abbreviations: DAPI, 4’,6-diamidino-2-phenylindole; EC, ependymal cell; LV, lateral ventricle; OPN, osteopontin; SVZ, subventricular zone.
The SVZ Is Hypoxic
The SVZ is hypoxic because the transcription factors HIF-1α and HIF-2α were present in the cytoplasm and in nuclei of cells including ECs in all seven SVZ samples. HIF-1α was more abundantly expressed than HIF-2α (Fig. 7). Remarkably, walls of blood vessels were strongly positive for HIF-1α, whereas HIF-2α expression was virtually absent in blood vessels walls (Fig. 7).
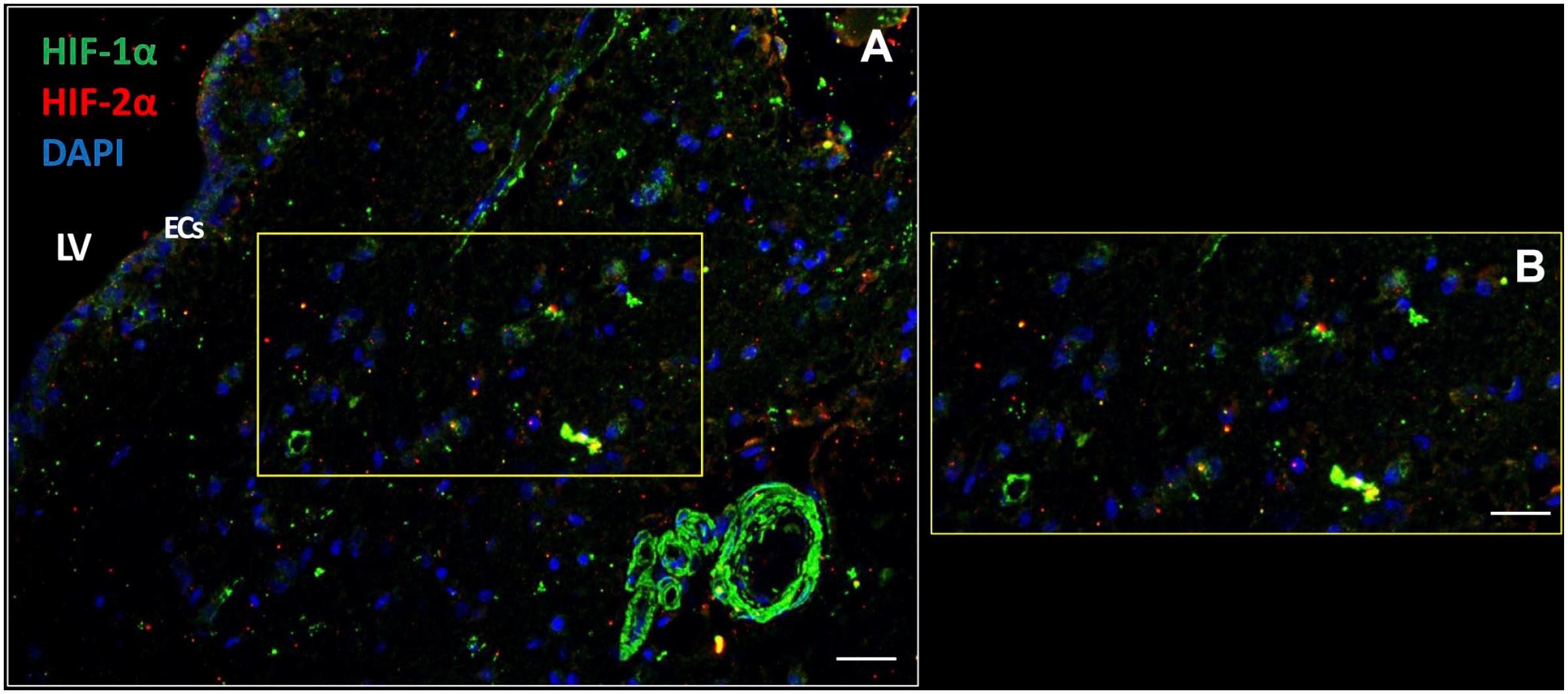
The SVZ niche is hypoxic. Transcription factors HIF-1α and HIF-2α are expressed intracellularly in the cytoplasm and nuclei of various types of cells in the SVZ of glioblastoma patient samples (A, B) including the ECs (A). HIF-1α expression is also strongly expressed in blood vessel walls (A). DAPI is used for nuclear counterstaining. Scale bars: A = 100 µm; B = 200 µm. Abbreviations: DAPI, 4′,6-diamidino-2-phenylindole; EC, ependymal cell; LV, lateral ventricle; HIF, hypoxia-inducible factor; SVZ, subventricular zone.
Discussion
We have immunohistochemically characterized the SVZ of glioblastoma patients to show whether the SVZ as NSC niche is morphologically similar to hypoxic peri-arteriolar GSC niches in glioblastoma and whether GSCs are present in the SVZ of brains of glioblastoma patients. To the best of our knowledge, this study is the first to demonstrate that both NSCs and GSCs are localized specifically in the human SVZ at distance of the glioblastoma tumor. Discrimination between NSCs and GSCs was performed on the basis of expression of the biomarker CD9. Furthermore, hypoxia was found to be an essential element of the SVZ, whereas the chemokines SDF-1α and OPN and their receptors CXCR4 and CD44, respectively, were abundantly expressed in the SVZ. In Fig. 8, a schematic illustration demonstrates the morphological similarities and differences between the peri-arteriolar GSC niche in glioblastoma (Fig. 8A)15–18 and the SVZ as NSC niche in glioblastoma patients (Fig. 8B). Similarities between peri-arteriolar GSC niches in glioblastoma tumors and NSC niches in the SVZ are the hypoxia and the same chemokines and their receptors. The major difference is that the central role of arterioles in GSC niches is missing in the SVZ. In the SVZ, ECs and GFAP-positive cells are relevant elements of the SVZ (Fig. 8).
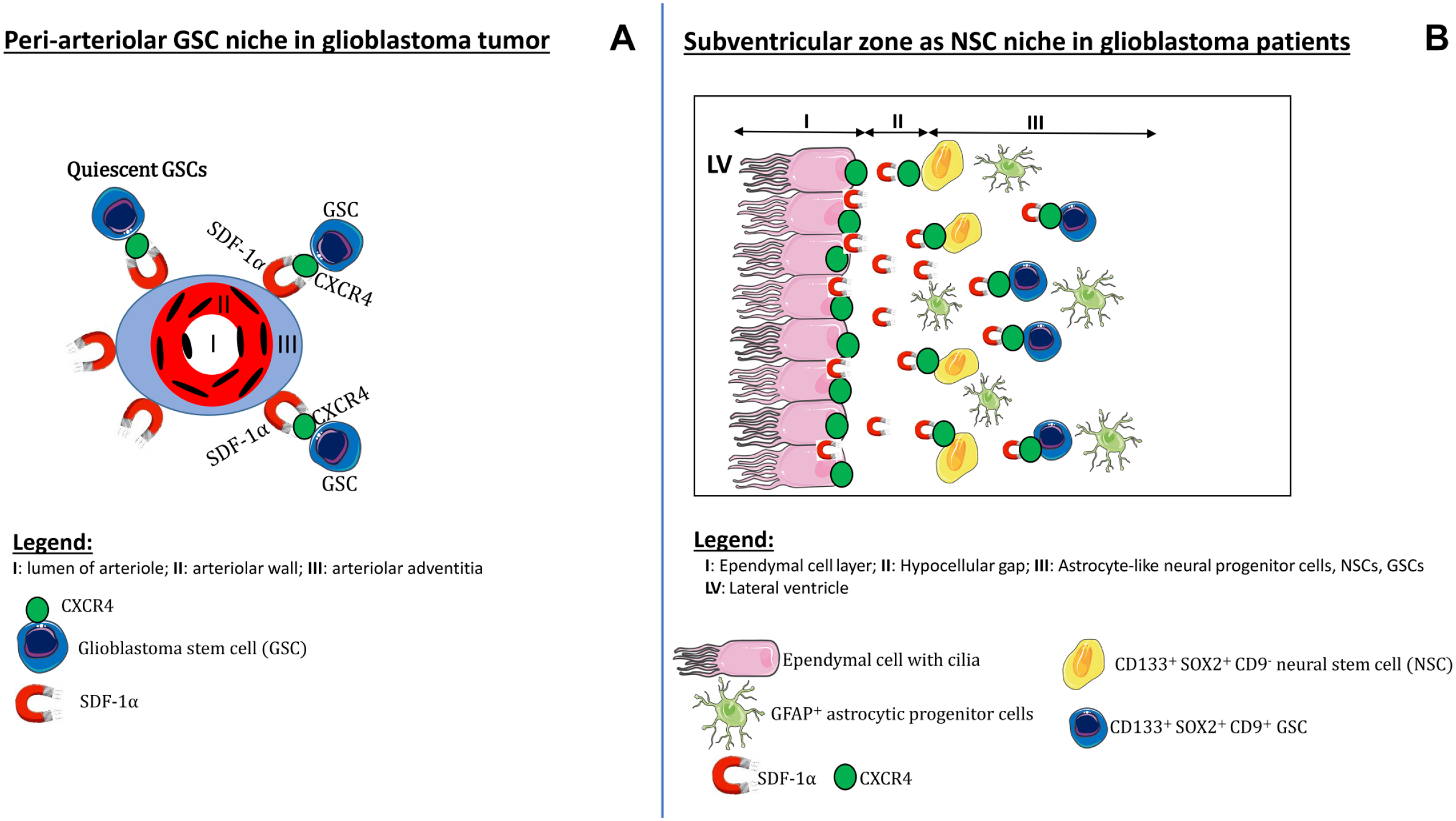
Schematic illustrations of niches. The peri-arteriolar GSC niche in glioblastoma tumors (A) and the SVZ as NSC niche in glioblastoma patients, containing NSCs and GSCs (B). Abbreviations: GSC, glioblastoma stem cell; NSC, neural stem cell; SVZ, subventricular zone.
SVZ studies have been performed thus far almost exclusively in mouse models26,63–68 to elucidate the identity of NSCs. Initially, it was assumed that the monolayer of ECs of the SVZ are NSCs.43,69 But then, it became generally accepted that GFAP-positive cells in the SVZ are NSCs, as elimination of GFAP-positive cells resulted in loss of neurosphere-forming cells in mice.41,70 However, a few studies reported that GFAP-positive cells are neural progenitor cells,43,71 whereas NSCs express CD133 and SOX2 and can generate neurospheres that are multipotent.43,63,64
On the basis of our data in human SVZ, we conclude that the CD133-positive SOX2-positive but CD9-negative cells in the third layer of the SVZ are NSCs (Fig. 4A, B) and that CD133-positive SOX2-positive CD9-positive cells in the SVZ are GSCs (Fig. 4C, D). Because GFAP is not a stem cell biomarker but a biomarker of differentiated astrocytes,43,66 we conclude that the GFAP-positive cells in the SVZ are astrocytes and possibly astrocytic-like neural progenitor cells.
NSCs can differentiate into neurons, astrocytes, and oligodendrocytes 71 and these cell types are part of the microenvironment that is essential for the maintenance of stemness of NSCs. 72 In line with the above, our in vitro immunocytochemical analyses revealed that differentiated glioblastoma cells express GFAP (Fig. 1B) and do not express stem cell biomarkers CD133 and SOX2 (Fig. 1A), whereas GSCs do not express GFAP (Fig. 1E), but express CD133, SOX2, and CD9 (Fig. 1D, F). We also demonstrated that GSCs express receptors CXCR4 and CD4416,17 which are stem cell biomarkers too73,74 although not very specific, because these receptors are expressed by various other cell types as well (Figs. 5, 6). Heterogeneity is a hallmark of glioblastoma tumors in which various subtypes of GSCs are present. 75 GSCs that express biomarker CD44 belong to the mesenchymal subtype of GSCs and are therefore most aggressive and therapy-resistant76,77 and their presence in the SVZ (Fig. 6) may well promote GSC survival and therapy-resistance.
The exact functions of ECs in the SVZ are still unclear. As stated above, ECs have been considered to be NSCs43,69 and our immunofluorescence images demonstrate that ECs express CD133 and SOX2 (Figs. 3, 4) which would be in line with their stemness. However, ECs also express chemokines SDF-1α and OPN (Figs. 5, 6) and this expression pattern resembles that of endothelial cells which are the center of peri-arteriolar GSC niches in glioblastoma tumors. These endothelial cells also express CD13378–80 and SOX2 and secrete chemokines such as SDF-1α and OPN for the maintenance of GSC stemness.16–18 We suggest that the ECs in the SVZ are required for the maintenance of NSCs in a similar way as endothelial cells are in peri-arteriolar GSC niches in glioblastoma. Endothelial cells have been shown to secrete factors that are essential for the maintenance of GSC stemness.14,81
Lee et al. 82 provided evidence that glioblastoma tumors originate from SVZ-residing NSCs. It was shown in patient-derived brain tissues that NSCs in the SVZ contain driver mutations that are also present in the matching tumors at a distance from the SVZ. Moreover, NSCs that carry driver mutations migrate out of the SVZ to develop into a malignant tumor at distance in the brain.82,83
The question arises whether GSCs in the SVZ are mutated NSCs82,83 or that they have migrated from the glioblastoma tumor into the SVZ on the basis of chemokines SDF-1α, OPN, and/or pleiotrophin that is highly expressed by neural progenitor cells in the SVZ as was demonstrated recently in mouse models. 68
In conclusion, the SVZ as NSC niche is morphologically different from peri-arteriolar GSC niches in glioblastoma tumors but the hypoxic conditions and presence of the same chemokines and their receptors in the SVZ indicate that the mechanisms involved in the protective conditions are similar in GSC niches and the SVZ. The presence of GSCs in the SVZ, that may well be the cause of recurrence of glioblastoma tumors after therapy, emphasizes the necessity to develop novel therapeutic treatment strategies with the aim to mobilize GSCs out of their protective peri-arteriolar GSC niches in the tumor and out of the SVZ to sensitize GSCs to chemotherapy and radiotherapy.
Supplemental Material
sj-pdf-1-jhc-10.1369_0022155421994679 – Supplemental material for Immunohistochemical Detection of Neural Stem Cells and Glioblastoma Stem Cells in the Subventricular Zone of Glioblastoma Patients
Supplemental material, sj-pdf-1-jhc-10.1369_0022155421994679 for Immunohistochemical Detection of Neural Stem Cells and Glioblastoma Stem Cells in the Subventricular Zone of Glioblastoma Patients by Vashendriya V.V. Hira, Remco J. Molenaar, Barbara Breznik, Tamara Lah, Eleonora Aronica and Cornelis J.F. Van Noorden in Journal of Histochemistry & Cytochemistry
Footnotes
Acknowledgements
The authors thank Jasper Anink and Theo Dirksen from the Department of (Neuro)Pathology of the Amsterdam UMC at the location Academic Medical Center in The Netherlands for their technical assistance.
Competing Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Author Contributions
All authors have contributed to this article as follows: conception and design (VVVH, CJFVN), collection and/or assembly of data (VVVH, BB, CJFVN), data analysis and interpretation (VVVH, BB, EA, TL, RJM, CJFVN), manuscript writing (VVVH, CJFVN), final approval of manuscript (VVVH, BB, EA, TL, RJM, CJFVN), provision of study material (EA), and supervision of the entire study (CJFVN).
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This study was financially supported by the Dutch Cancer Society (KWF; UVA 2014-6839 and UVA 2016.1-10460; V.V.V.H., R.J.M., C.J.F.V.N.), the Slovenian Research Agency (Project J3-2526; CJFVN, BB, TL), the European Program of Cross-Border Cooperation for Slovenia-Italy Interreg TRANS-GLIOMA (Program 2017; T.L.), the IVY Interreg Fellowship (V.V.V.H.), and the Slovenian Research Agency (Program P10245; T.L., and Postdoctoral project Z3-1870 to B.B.), and R.J.M. was supported by the Fondation pour la Recherche Nuovo-Soldati 2019.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.