
Abstract
The immediate alterations following lesions cannot be investigated by using fixed tissues. Here, we employed two-photon microscopy to study the alterations to the permeability of blood-brain barrier and to glio-vascular connections in vivo during the first minutes following cortical lesions in mice. Four models were used: (1) cryogenic lesion, (2) photodisruption using laser pulses, (3) photothrombosis, and (4) bilateral carotid ligation. Sulforhodamine101 was used for supravital labeling of astrocytes and dextran-bound fluorescein isothiocyanate for the assessment of extravasation. Transgenic mice, in which the endothelium and astrocytes expressed a yellow fluorescent protein, were also used. Astrocytic labeling in vivo was verified with postmortem immunostaining against glial fibrillary acidic protein (GFAP). Summary of results: (1) the glio-vascular connections were stable in the intact brain with no sign of spontaneous dynamic attachment/detachment of glial end-feet; (2) only direct vascular damage (photodisruption or cryogenic) resulted in prompt extravasation; (3) even direct damage failed to provoke a prompt astroglial response. In conclusion, the results indicate that a detachment of the astrocytic end-feet does not precede the breakdown of blood-brain barrier following lesions. Whereas vasogenic edema develops immediately after the lesions, this is not the case with cytotoxic edemas. Time-lapse recordings and three-dimensional reconstructions are presented as supplemental materials.
Introduction
The aim of the study was to investigate the events during the first minutes following cortical lesions in mice in vivo. These events cannot be studied by using traditional histological methods because of the delay due to fixation. Some observations, however, indicated that these minutes may be important. Following cryogenic lesions, extravasation was observed in a few minutes.1–5 In our experiments, the shortest available postlesion time until fixation was about 3 min. In these brains, extravasation was already detected around the vessels. 5 Signs indicating glio-vascular detachment were also observed: The cerebrovascular laminin immunoreactivity had become detectable and electron microscopic investigation revealed a separation of astroglial and vascular basal laminae. 5
The postlesion appearance of cerebrovascular laminin immunoreactivity refers to the separation of the astroglial and vascular basal laminae which makes the otherwise hidden laminin epitopes available for antibodies.6,7 Electron microscopic investigations of Jaeger and Blight 8 and Kwon et al. 9 reported detachment of astrocytic end-feet from vessels.
The detachment may have a role in the postlesion extravasation since the glio-vascular coupling has a basic importance in the maintenance of blood-brain barrier.10,11 In our former experiments, 5 it was impossible to clarify whether a glio-vascular detachment preceded the extravasation because delay caused by fixation made the events of the first minutes inaccessible for investigation.
In the present study, we applied two-photon microscopy. It allows imaging of the brain to depths of hundreds of micrometers and makes examination of early postlesion alterations possible in vivo. Two-photon microscopy uses a fluorochrome that can be excited by the simultaneous absorption of two photons of twice the wavelength than the wavelength required for a single photon to excite the same fluorochrome.12,13 It means that two simultaneous red-infrared range photons can excite molecules with absorption maximum in the UV-blue-green range. Due to the low probability of such excitation, a high flux of photons is necessary, which is provided by a focused femtosecond laser. Longer excitation wavelength involves less scattering and therefore deeper tissue penetration, which makes observations possible inside living tissues.12,13
The in vivo investigation requires in vivo staining. Sulforhodamine 101 (SR101)14,15 was applied for the supravital fluorescent labeling of astrocytes whereas the extravasation was visualized using fluorescein isothiocyanate (FITC) bound 70 kDa dextran, which can pass only impaired blood-brain barrier. 16 Transgenic mice, in which the endothelium and astrocytes expressed a yellow fluorescent protein (YFP), were also used. Astrocytes labeled in vivo were verified with postmortem immunofluorescent staining against glial fibrillary acidic protein (GFAP).
Besides cryogenic lesion our investigation was extended to other lesion types which result in different (vasogenic or cytotoxic) types of cerebral edema. Four experimental models were investigated: (1) cryogenic lesion,2,17 (2) photodisruption of vessels using the laser of the two-photon microscope,18,19 (3) photothrombosis following administration of Rose Bengal dye,19,20 and (4) bilateral carotid artery ligation.21,22
The observation started before the lesion and continued immediately following the lesion during the next 2 hr. This way our observation period overlapped with the investigation periods following traditional histological methods involving fixation.
The aim of the prelesion observation was to observe whether the glio-vascular connections show any dynamic attachment/detachment in the intact brain. Postlesion observation focused on extravasation and perivascular astroglial responses especially a withdrawal of vascular processes. Besides conventional figures, 3D reconstruction and time-lapse recordings are presented as supplemental material.
Materials and Methods
Young adult mice (16 of C57BL/6J strain, 15–20 g) of either sex were used. The animals were supplied with food and water ad libitum, and kept in artificial 12/12 h light-and-dark periods. Five transgenic mice were also used in which endothelium and astrocytes expressed YFP. Experiments were performed according to the Committee on the Care and Use of Laboratory Animals of the Council on Animal Care at the Semmelweis University of Budapest, Hungary (22.1/3491/003/2008) in accordance with the guidelines of European Union Directive (EU Directive 2010/63/EU). The animals were randomly collected from several cages.
The transgenic mice (FVB/AntTg(CAG-yfp sb)27 strain) were produced in the Institute of Experimental Medicine of the Hungarian Academy of Sciences. The marker gene (YFP) and the promoter (CAG) flanked with inverted terminal repeats (SB ITR) were co-injected as circular plasmid DNA with SB100X mRNA (SB—Sleeping Beauty transposase) into fertilized FVB/Ant mouse eggs. Eggs were transplanted into B6CBA F1 recipient females. In vivo YFP expression was monitored with FHS/T01 lamp through appropriate filters (BLS ltd.) in P0 pups.
SR101 (20 mg/kg body weight, Sigma-Aldrich, St. Louis, MO) and/or FITC-dextran 70 kDa (100 mg/kg body weight, Sigma-Aldrich, St. Louis, MO) were diluted (10 mg/ml and 100 mg/ml, respectively) in physiological sodium chloride solution and administered through the left jugular vein 2 hr before the experiment.
Examinations were performed in deep anesthesia with ketamine (Calypsol 50 mg/ml, Richter Gedeon, Budapest, Hungary) and xylazine (Nerfasin 20 mg/ml, LeVet. Pharma, Oudewater, The Netherlands), 100 and 20 mg/kg body weight, respectively, intramuscularly. A 3 × 3 mm bone piece was removed by drilling so that a “window” was formed on the skull corresponding to the dorsoparietal cortex on the right side. Lesions were performed as described later. Monitoring started 30 min before the lesion and continued immediately following the lesion for 2 hr. The animals were covered with a small blanket to prevent cooling, so their temperature remained between 36C to 37C. The in vivo labeling was combined with postmortem immunofluorescent staining for the astroglial marker GFAP (see later).
Observations were performed with a Femto2D (Femtonics Ltd., Budapest, Hungary) laser scanning microscope which contained a tunable ultrafast laser (MaiTai DeepSee Laser, Spectra Physics, Santa Clara, CA). Laser intensity was adjusted by turning a polarizer filter. In addition to single images, stacks of images taken at series of focal planes (Z-stacks) were created to reconstruct three-dimensional (3D) structures. For imaging, a 10× objective (UPLSAPO 10X, N. A. 0.4 Olympus) was used. The excitation wavelengths were 920 nm for the combination of SR101 and FITC and 950 nm for SR101 plus YFP. Fluorescent emission signals were detected in two channels, for green (480–560 nm) and red (600–700 nm) wavelength domains. The measurement system was controlled by a software package developed by Femtonics Ltd. Image processing was performed with the open source ImageJ program (ImageJ 1.48v, NIH, Wayne Rashband, http://imagej.nih.gov/ij). This setup allows observations in 3 to 400 µm depth from the surface of the pia mater, for 3D reconstructions and time-lapse recordings. The time-lapse recordings and 3D reconstructions are attached as supplemental materials; their starts are given at the corresponding parts of Results and Legends in min:sec.
During observation respiration was detectable in the form of horizontal color strips on the display, whereas blood flow evoked a characteristic stripe pattern in the vessels, which was generated by moving red blood cells. Both patterns are visible in figures (Figs. 1A and 2A).
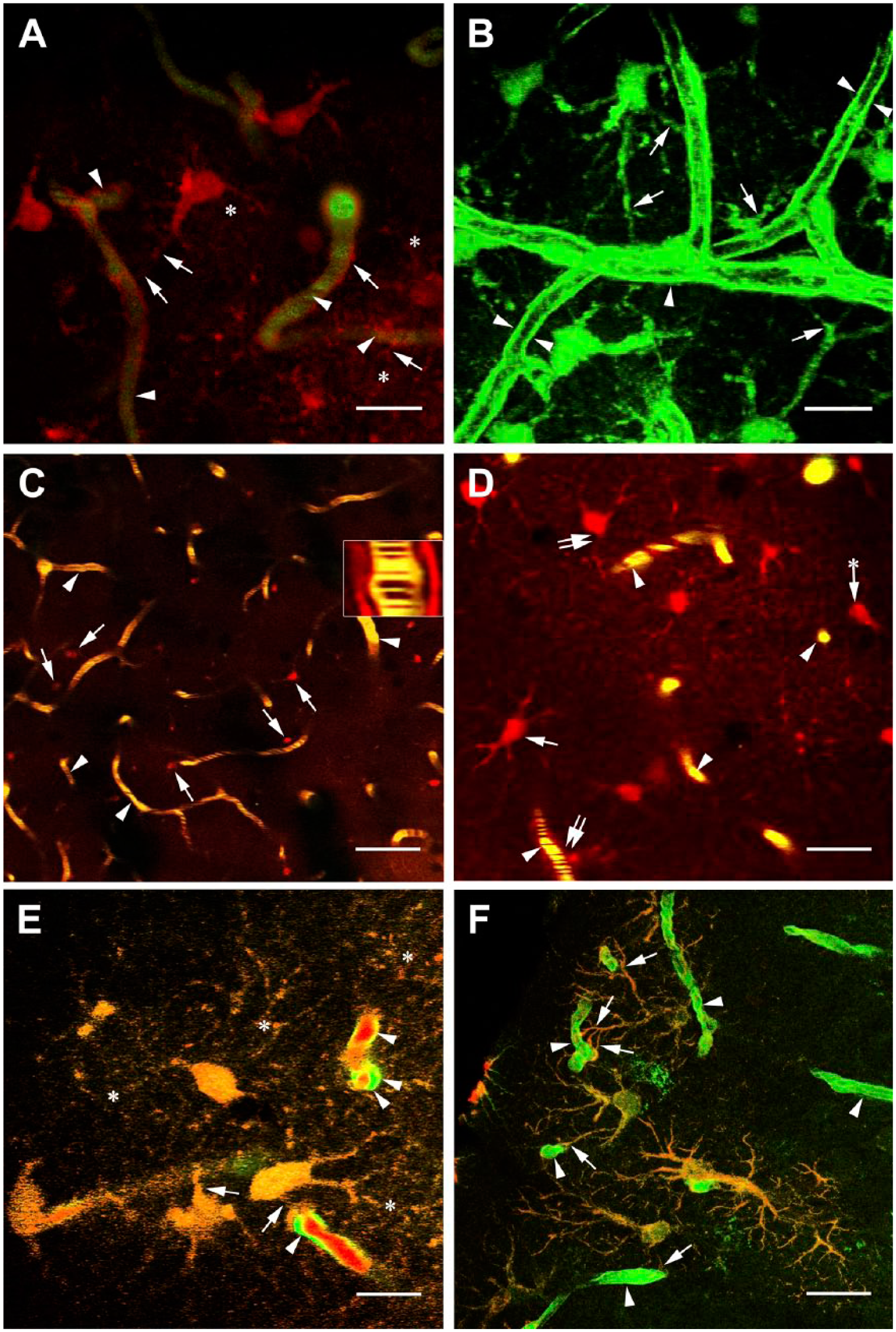
Vessels and perivascular astrocytes in intact mouse brains. (A) Detail of intact cortex of C57BL/6J “wild” (i.e., not transgenic) mouse. The vessels are filled with FITC-dextran (green, arrowheads) whereas the astrocytes are labeled with SR101 (red, arrows point to their vascular processes); a fine network of astrocyte processes is recognizable (asterisk). The stripe pattern in the vessels indicates blood flow. Scale bar: 20 µm. A 3D reconstruction in the supplemental material starts at 0:16 min. (B) Detail of intact cortex of YFP-transgenic mouse: vessels and perivascular astrocyte-like cells. Arrows point to perivascular processes of astrocytes arrowheads point to endothelium. The lumen seems to be empty: The vessels are not filled with dye. Scale bar: 15 µm. A 3D reconstruction in the supplemental material starts at 0:33 min. (C) SR101 (red) and FITC-dextran (green) labeling in the intact cortex of C57BL/6J “wild” mouse. The astrocytes (arrows) are red labeled with SR101; the vessels (arrowheads) are yellow, which is a combination of red and green fluorescence. Note the stripe pattern indicating blood flow (see also the inset and the supplemental material from 0:49 min); the gap seen in the inset between the blood flow and the perivascular astrocyte processes corresponds to the position of endothelium. Scale bar: 150 µm. (D) Enlarged detail of an intact cortex with similar labeling as before. Arrowheads: vessels, double arrows: vessel-contact astrocytes, arrows: noncontact astrocytes. Arrow with asterisk labels a noncontact astrocyte adjacent to a vessel within a distance of a diameter of a cell body. Astrocytes in larger distance from vessels were not taken into consideration for the statistical analysis shown in Table 1. Scale bar: 30 µm. (E) SR101 labeling applied on a YFP-transgenic mouse. The merge of red and green fluorescence results in the orange-yellow color of astrocytes; the vascular processes of astrocytes are pointed by arrows. Asterisk: network of fine astrocyte processes. The endothelium (arrowheads) is green as it expresses YFP, whereas the blood inside the vessels is stained red by SR101. Scale bar: 15 µm. (F) YFP-expressing astrocytes immunostained postmortem against GFAP. The colocalization (yellowish color) is conspicuous in the processes of astrocytes (arrows). The vessels (arrowheads) display only green color. This photomicrograph was taken by a confocal microscope. Scale bar: 30 µm. Abbreviations: FITC, fluorescein isothiocyanate; 3D, three-dimensional; YFP, yellow fluorescent protein.
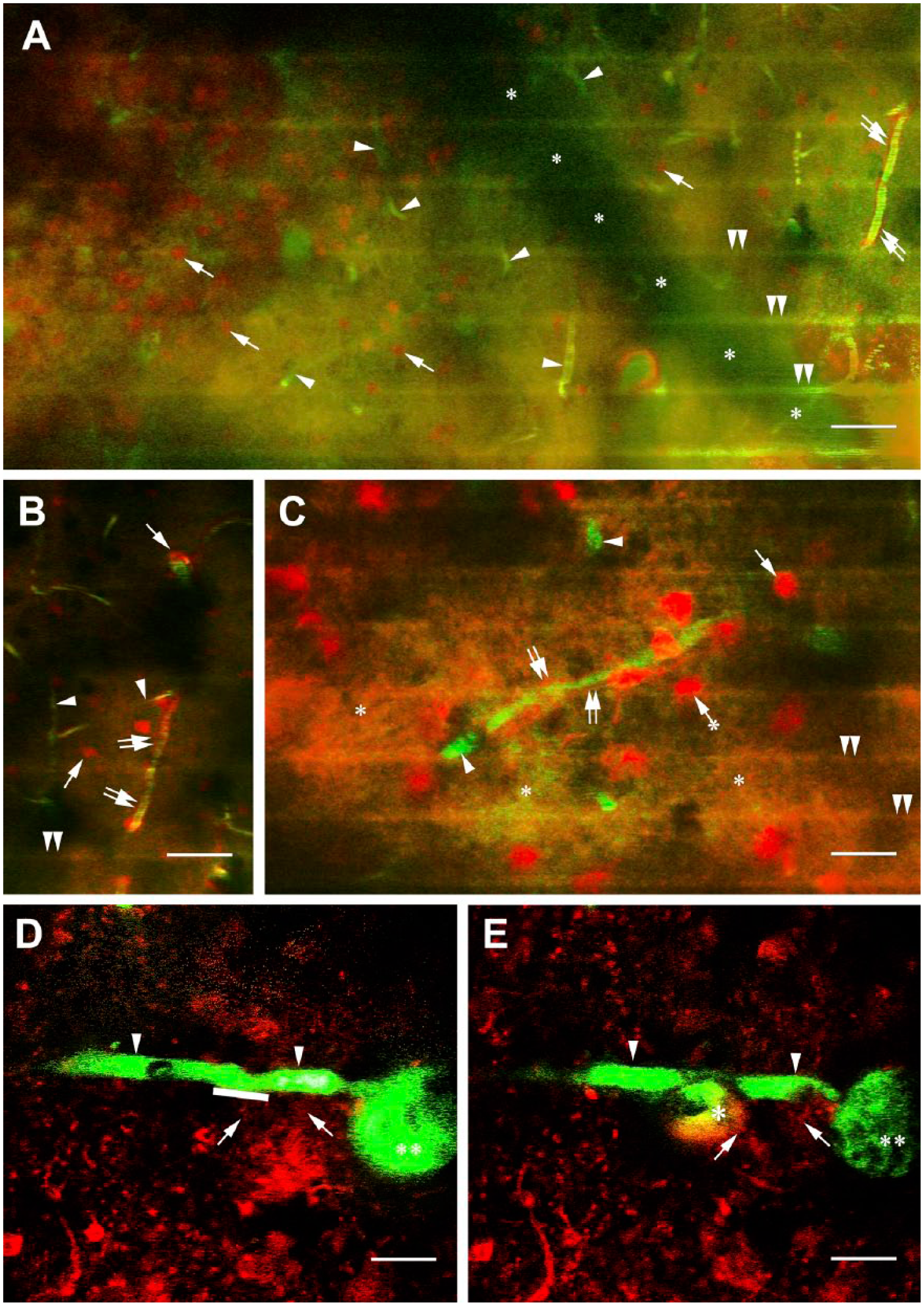
Effects of cryogenic lesion and photodisruption in the cortices of C57BL/6J “wild” mice. (A) Cryogenic lesion in the presence of labeling with SR101 (red) and FITC-dextran (green). Arrowheads: vessels almost covered by the greenish clouds of extravasation, arrows: astrocytes; their glio-vascular connections are still recognizable (double arrows). The horizontal green stripes (double arrowheads) are created by the respiration of mouse. The asterisks trace the shadow of a large vessel not in focus. Scale bar: 150 µm. (B) The vessel pointed by double arrows in panel A but color filtered and slightly enlarged. Labeling is the same. Scale bar: 100 µm. (C) A vessel from a similar experiment. The extravasation (asterisks) is colored by both FITC-dextran and SR101. Arrowheads: vessels, double arrows: vessel-contact astrocytes, arrow: noncontact astrocytes. Arrow with asterisk labels a noncontact astrocyte adjacent to a vessel. Astrocytes in larger distance from vessels were not taken into consideration for the statistical analysis shown in Table 1. The horizontal green stripes (double arrowheads) are created by the respiration of mouse. Scale bar: 20 µm. (D and E) The effect of photodisruption, panel D: before lesion, panel E: after lesion. Arrowheads: the targeted vessel, asterisk: marks the extravasation, double asterisk: large vessel perpendicular to the focus plane, arrows: contacts of astrocytes. The stripe along the vessel indicates the exact site of irradiation. Scale bar: 10 µm. See a time-lapse recording in the supplemental material from 1:38 min. Abbreviation: FITC, fluorescein isothiocyanate.
Cryogenic lesions were performed in 6 C57BL/6J mice with a copper rod (2 mm thick) cooled in frozen carbon dioxide. The frozen carbon dioxide was mixed with acetone and was in a plastic cylinder around the copper rod, therefore, it did not contact the brain. The rod with the cylinder was adjusted to a stereotaxic instrument. This way its smooth, polished tip was positioned gently to the leptomeninx-covered brain surface. Former experiments 23 suggested that the optimal contact time is 20 sec.
For photodisruption (2 C57BL/6J and 2 YFP mice) the laser of the two-photon microscope (780 nm, 60–80 mW) was focused on a vessel, scanning it repeatedly for 3 sec according to Davalos et al. 18
For photothrombosis (4 C57BL/6J mice), Rose Bengal dye (40 mg/kg body weight, Sigma-Aldrich, St. Louis, MO) was diluted (10 mg/ml) in physiological sodium chloride solution and was administered through the jugular vein. Thrombosis was induced with 5 min illumination following the 30-min prelesion observation. The light source was a halogen lamp with fiberoscope for operations (Motic MLC-150C Microscope Cold Light Source, 5000Kelvin, diameter: 3 mm), which does not warm tissues.
Bilateral carotid ligation (4 C57BL/6J mice) was prepared in advance and tightened after the abovementioned 30-min prelesion observation of the intact brain.
Photos were taken as soon as possible: within seconds following laser combustions and photothromboses, and within 1 min following bilateral carotid occlusions and cryolesions.
The vessel-contact and noncontact astrocytes were counted on photomicrographs. The significance of the quantitative differences before and following lesions was tested according to the χ2 and Mann–Whitney tests at a significance level of p<0.05. (For details see Results, Quantitative analysis and Table 1.)
Comparison of Vessel-Contact and Noncontact Astrocytes Before and After Lesions.
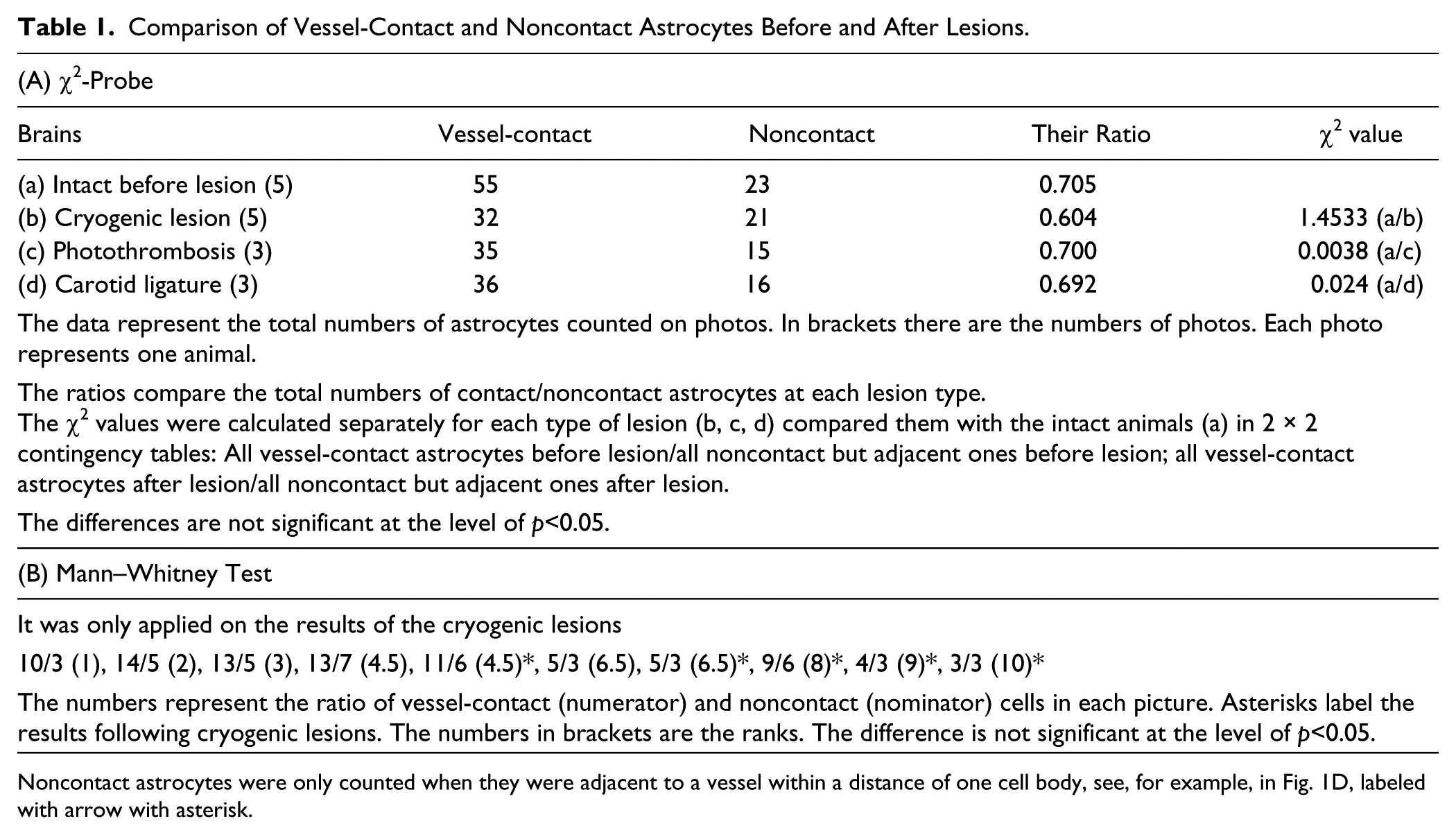
Noncontact astrocytes were only counted when they were adjacent to a vessel within a distance of one cell body, see, for example, in Fig. 1D, labeled with arrow with asterisk.
For postmortem immunohistochemistry the brains were fixed by immersion in 4.0% paraformaldehyde dissolved in 0.1 M phosphate buffer (pH 7.4) overnight at 4C. Coronal sections of 50 µm with the lesioned area were cut with a vibratome (Leica VT1000 S, Leica Microsystems GmbH, Wetzlar, Germany) and rinsed in phosphate buffered saline 0.1 M, pH 7.4 (PBS, Sigma-Aldrich, St. Louis, MO) overnight. Floating sections were pretreated with normal goat serum (diluted to 20% in PBS) for 90 min at room temperature. This and the following steps were followed by a rinse in PBS (30 min, at room temperature). The sections were then incubated with monoclonal mouse anti-GFAP antibody (Novocastra, Newcastle, England) for 40 h at 4C. The antibody was diluted to 1:100 (final concentration 100 µg/ml) in PBS containing 0,5% Triton X-100 and 0.01% sodium azide. Fluorescent secondary antimouse immunoglobulin (fluorogen: Alexa 555, red, Life Technologies, Eugene, OR) was applied at room temperature for 3 hr. The sections were finally rinsed in PBS (1 hr, at room temperature), mounted on slides, coverslipped and sealed with lacquer. Photomicrographs were taken with a Radiance-2100 (BioRad, Hercules, CA) confocal laser scanning microscope.
Results
Intact Brain
In wild-type mice vessels were delineated by administering FITC-dextran and astrocytic processes were visualized by SR101, so glio-vascular connections were visible (Fig. 1A, see also 3D reconstruction in the supplemental material from 0:16 min). In the transgenic mice, both endothelial cells and astrocytes were visualized by yellow fluorescence but they were distinguished by their characteristic morphology (Fig. 1B, see also 3D reconstruction in the supplemental material from 0:33 min). In the vessels filled with FITC-dextran a characteristic stripe pattern was observed. This pattern was generated by moving red blood cells and indicated blood flow (Fig. 1C, see time-lapse recording in the supplemental material from 0:49). In general, in medium-power visual fields like in Fig. 1C 25 to 30 perivascular astrocytes were seen with processes to vessels. Figure 1D demonstrates a part at higher magnifications, where the close glio-vascular contacts can be observed. A time-lapse recording proves that they remain stable; no spontaneous dynamic detachment/attachment of processes was observed (see the supplemental material from 0:05 min). YFP-expressing astrocytes were also labeled by intravenous application of SR101 (Fig. 1E). Postmortem GFAP immunohistochemistry confirmed that both YFP and SR101 labeled astrocytes (Fig. 1F).
Cryogenic Lesions
The cryogenic lesions led to hemostasis immediately; blood flow was only observed away from the area of the lesion whose diameter was about 2 mm. The vessels became “leaky” so the FITC-dextran tracer penetrated the surrounding area immediately (Fig. 2A–C). Glio-vascular connections were visible, but no motion of astroglial processes was observed during the 2-hr postlesion observation period.
Other Models
Photodisruptions provoked only modest damage (Fig. 2D and E). The vessels became immediately leaky, but the extravasation remained confined as compared with the extended extravasation found following cryogenic lesions. When astrocytes were labeled with SR101 or using YFP-transgenic mice no astroglial response was observed even when the laser was focused directly on an astrocyte (see also time-lapse recording in the supplemental material from 1:38 min).
Both photothromboses (Fig. 3A–C, see also 3D reconstruction and time-lapse recording in the supplemental material from 0:59 min) and bilateral carotid artery ligations (Fig. 3D) interrupted the cerebral circulation immediately, but neither extravasation nor change of glio-vascular connections, nor other astrocyte response was seen either immediately after lesion or during the further 2-hr postlesion observation period.
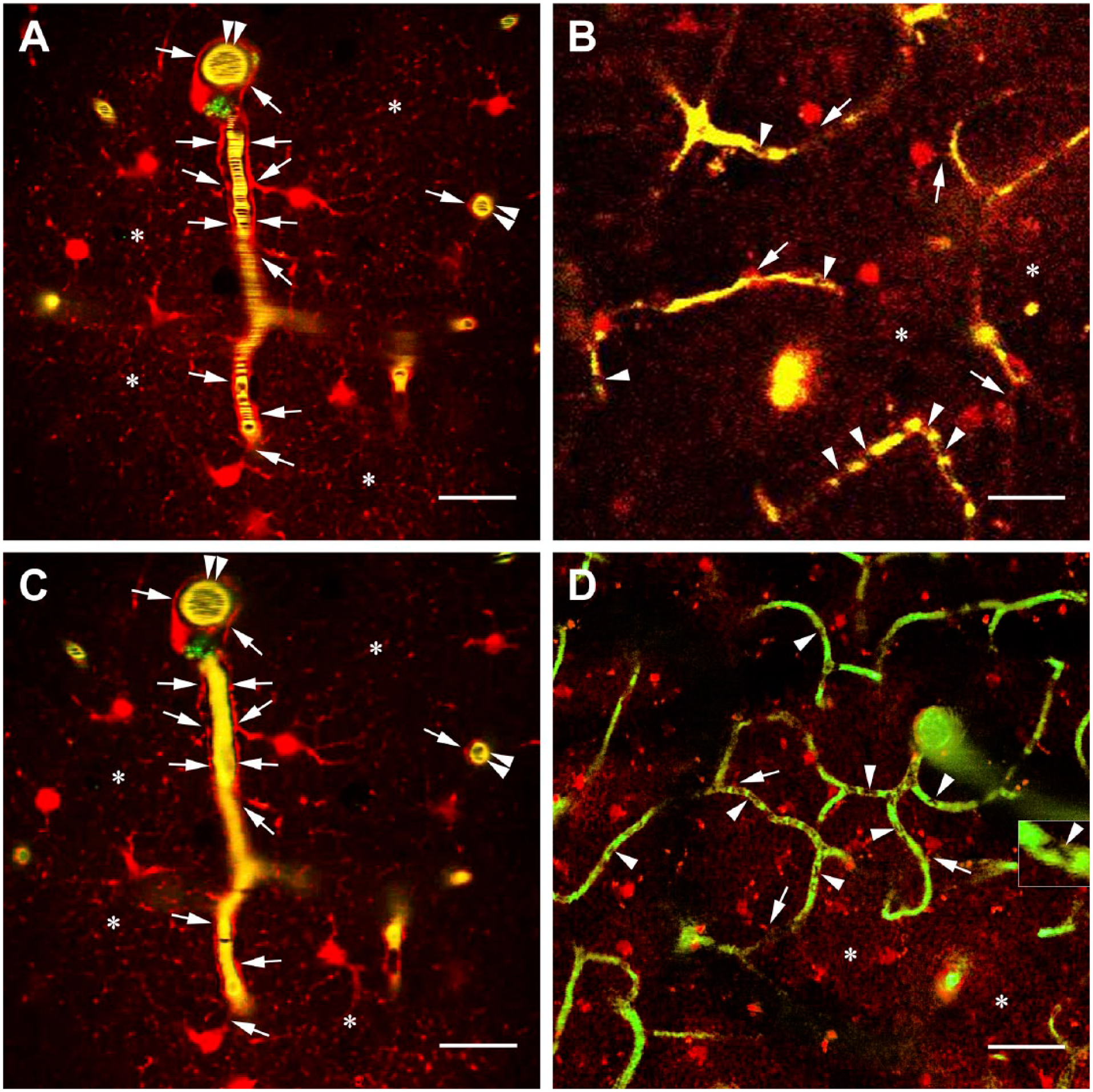
Effects of photothrombosis and bilateral common carotid ligature in the cortices of C57BL/6J “wild” mice. (A) An intact cortex with SR101 and FITC-dextran labeling before a Rose Bengal-induced photothrombosis. Note the stripe pattern within the vessel, which indicates blood flow. Arrows: perivascular astrocyte processes, double arrowheads: a vessel perpendicular to the focus plane, asterisks: network of fine astrocytic processes. Scale bar: 50 µm. (B) A cortex with SR101 and FITC-dextran labeling after photothrombosis. No stripe pattern indicating blood flow is visible in the vessels. The dense intravascular bodies (arrowheads) are stagnant red blood cells (see also a time-lapse recording in the supplemental material from 0:59 min). Arrows: perivascular astrocytes with vascular processes, asterisks: network of fine astrocytic processes. Scale bar: 100 µm. (C) The same area as in panel A, but following photothrombosis. No stripe pattern indicating blood flow is visible in the vessels. The positions of astrocyte processes (arrows) did not change. Asterisks: network of fine astrocytic processes. Scale bar: 50 µm. A 3D reconstruction in the supplemental material starts at 1:19 min. (D) The effect of the bilateral ligature of the common carotid arteries. No stripe pattern indicating blood flow but deformed red blood cells are visible in the vessels (arrowheads, see also inset) like in panel B. The glio-vascular contacts persist (arrows). Asterisks: network of fine astrocytic processes. Scale bar: 120 µm. Abbreviation: FITC, fluorescein isothiocyanate.
Quantitative Analysis
In the case of cryogenic lesion, photothrombosis and carotid ligature two types of astrocytes were counted on photomicrographs taken before and after lesion like Figs. 1C and 2C. Each animal was represented by one photo. The counting was performed by screening the photo along its zoomed rectangles similar to Fig. 1D.
One cell type was the vessel-contact astrocyte the other type was a noncontact astrocyte which, however, was adjacent to a vessel within a distance of a diameter of a cell body, that is, a distance from which a vessel-contact could be formed. This latter cell type is demonstrated in Fig. 1D labeled with an arrow plus asterisk. Astrocytes at larger distances from vessels were not taken into consideration.
These data were analyzed according to the χ2 test (Table 1A) in a 2 × 2 contingency table. The analysis was performed separately for each lesion type.
In the case of cryogenic lesions, Mann–Whitney test was also applied (Table 1B), since the difference was nearly significant according to the χ2 test. Both tests indicated that lesions resulted in no significant difference in the number of vessel-contact cells.
In the case of photodisruption, the lesioned areas were so small they involved only one or two astrocytes. As mentioned above, the astrocytes did not show any visible alteration even in the case of direct combustion (see a time-lapse recording in the supplemental material from 1:38 min). Therefore quantitative analysis was not performed here.
Discussion
The Mechanisms of the Lesions
Cryogenic lesion is a widely used model of cerebral damage. It results in a reproducible tissue damage, which shares many similarities with models of neurotrauma and it is applied frequently to study the role of blood-brain barrier in experimental lesions.1–4,24–27
The early appearance of extravasation following cryogenic lesions was attributed to a direct effect of freezing on vessels. 2 In our models, however, direct lesions of vessels with photodisruption resulted in only minimal extravasation as compared with cryogenic lesions. An increase in the trans-endothelial vesicular transport 4 and membrane phospholipid peroxidation 28 are thought to be factors responsible for blood-brain barrier breakdown in cryogenic lesions.
“Sham” cryogenic lesions, that is, applying the copper rod without dry ice were performed in a previous study. 5 They resulted in no considerable extravasation.
In photothrombosis lesions, the photoactivated Rose Bengal dye produces singlet oxygen. It damages the endothelium and leads to thrombus formation without a disruption of the vascular wall. 29
In the case of carotid ligature, a drawback is its inconsistency due to the irregular occurrence of the posterior communicating artery.21,22 This anastomosis makes rats resistant to bilateral common carotid occlusion. In gerbils, which lack this anastomosis, bilateral occlusion results in ischemia. In mice, the anastomosis is highly variable and strain-dependent. 21 In our experiments, the cerebral circulation stopped immediately when carotids were ligated and did not return during the observation period.
Stab wound is a frequent and simple method of lesion. However, according to our former observations23,30 stab wounds result in quite different “individual” forms of damage, and the extravasation is complicated by bleeding. Therefore this model was not applied.
The Labeling
In studies based on the in vivo labeling of astrocytes with SR101, it was smeared on the brain, 15 injected into it,31,32 administered intraperitoneally 33 or intravenously14,34 like in our experiments. According to a recent publication, 35 the multispecific thyroid hormone transporter underlies the uptake of SR101 into astrocytes.
FITC-dextran (of 70 kDa) labeling of extravasation was applied, for example, by Hultström et al., 16 for two-photon microscopy by Mc Caslin et al. 33 It only passes impaired blood-brain barrier; therefore, it is a proper marker of leakage.
Fixation by Perfusion or Immersion?
It is an important problem, whether perfusion may press the extravasation deeper into the brain tissue. In the case of intrinsic markers (e.g. immunoglobulins, plasma fibronectin), perfusion has frequently been used.26,36,37 However, these markers are usually unable to be visualized in vivo. Especially, Stewart et al. 4 does not recommend perfusion when an extrinsic marker (e.g., horse radish peroxidase) is applied following a short postlesion period.
Blood-brain Barrier and Glio-vascular Connections
Previous studies revealed postlesion cerebrovascular laminin immunopositivity following various experimental lesions: stab wounds,23,30,38 cryogenic lesion, 23 excitotoxic lesion, 39 arterial occlusion, 40 which can refer to impaired glio-vascular connections.6,7 A similar importance may be attributed to the postlesion disappearance of β-dystroglycan.23,40,41 Electron microscopic observations on glio-vascular decoupling have been mentioned in the “Introduction” section.8,9 These phenomena might play a role in the postlesion extravasation, since glio-vascular connections have a basic importance in the maintenance of blood-brain barrier.10,11
Several earlier two-photon studies investigated postlesion astroglial responses31,33,42–45 and found swelling of astrocytes 46 and extension of their processes toward the site of lesion. 47 These investigations, however, did not focus on the first postlesion minutes.
The present investigations do not support an early withdrawal of the vascular astrocyte end-feet or any other conspicuous astroglial movement. Together with our previous ones, 5 they suggest that it may be the extravasation which separates the astroglial and vascular basal laminae evoking laminin immunoreactivity and “detaching” astrocytes.
Different Roles of Astrocytes in Different Lesions
Cryogenic lesions result in vasogenic edema.2,3,24–27 In this type of cerebral edema, the perivascular astrocytes move the excess water from the intercellular space into the vessels via the aquaporin 4 localized in their end-feet.48–50
In the other form of cerebral edema, the cytotoxic (intracellular) edema, aquaporin 4 promotes water influx into the perivascular astrocytes from the vessels; it takes place, for example, in cerebral ischemia. 51 Note that in ischemic models (carotid ligature, photothrombosis) no extravasation was seen during the observation period.
It seems to be surprising, since edema regularly appears following cerebral ischemia. However, it was observed only following longer postlesion periods than used in our experiments. Following middle cerebral artery occlusion Garcia et al. 52 did not find extravasation in 30 min but in 3 hr. Following photothrombosis Hoff et al. 53 observed extravasation in 1 hr applying FITC-dextran of 40 kDa but only in 3 hr applying FITC-dextran of 70 kDa as we did. Summarizing the results (1) in intact brain: the glio-vascular connections were stable and there was no sign of dynamic attachment/detachment of astroglial end-feet; (2) only direct vascular damage (photodisruption or cryogenic) resulted in prompt extravasation; (3) even direct damage failed to provoke visible astroglial responses. The results indicate that a postlesion detachment of the astrocytic end-feet does not precede the breakdown of the blood-brain barrier. Whereas vasogenic edemas develop immediately after the lesion, this is not the case with cytotoxic edemas.
Supplemental Material
DS_10.1369_0022155418788390 – Supplemental material for The First Postlesion Minutes: An In Vivo Study of Extravasation and Perivascular Astrocytes Following Cerebral Lesions in Various Experimental Mouse Models
Supplemental material, DS_10.1369_0022155418788390 for The First Postlesion Minutes: An In Vivo Study of Extravasation and Perivascular Astrocytes Following Cerebral Lesions in Various Experimental Mouse Models by László Tóth, Dávid Szöllősi, Katalin Kis-Petik, István Adorján, Ferenc Erdélyi and Mihály Kálmán in Journal of Histochemistry & Cytochemistry
Footnotes
Acknowledgements
We thank Á. Dénes and G. Szalay (Institute of Experimental Medicine of the Hungarian Academy of Sciences) for technical advices to improve our methods. In addition, we thank Á. Ádám for the wild type mice and A. Z. Őz, S. Deák and Z. Gróti (Department of Anatomy, Histology, and Embryology, Semmelweis University, Budapest, Hungary) for the technical assistance. For style and grammar corrections thanks to J. Zerkowitz.
Competing Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Author Contributions
The participation was as follows: L. Tóth operations and two-photon microscopy; K. Kis-Petik two-photon microscopy; D. Szöllősi confocal microscopy and immunohistochemistry; I. Adorján elaboration of operation techniques; F. Erdélyi (Institute of Experimental Medicine of the Hungarian Academy of Sciences) providing the transgenic mice; and M. Kálmán (senior) for experimental design, evaluation of data, and formation of manuscript.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: The work was supported by the scientific budget of the Department of Anatomy, Histology, and Embryology, Semmelweis University, Budapest, Hungary; the Department of Biophysics and Radiation Biology, Semmelweis University; the MTA-SE Molecular Biology Research Group, Budapest, Hungary, where the authors are affiliated; and by the National Research, Development and Innovation Office under the contract VKSZ_14-1-2015-0052.
Ethical Approval
This article does not contain any studies on humans. All procedures involving animals were in accordance with the Committee on the Care and Use of Laboratory Animals of the Council on Animal Care at the Semmelweis University of Budapest, Hungary (22.1/3491/003/2008) and with the guidelines of European Union Directive (EU Directive 2010/63/EU).
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.