
Abstract
Interleukin-6 (IL-6) is a multifunctional cytokine considered to modulate bone homeostasis. Based on previous contradictory studies, we aimed to verify the influence of IL-6 deficiency on bone remodeling using an IL-6 knockout (IL-6-/-) murine model. Eight-month-old male mice, homozygous for the disrupted IL-6 gene, and their wild type (WT) littermates (control), were used. After transcardiac perfusion, tibiae were removed for histochemical analysis. Compared with the control group, IL-6 deficiency increased tartrate resistant acid phosphatase (TRAP)-positive osteoclast numbers and up-regulated the alkaline phosphatase (ALP) activity of osteoblasts in the metaphysis of the tibia. However, further analysis of serial histological sections from IL-6-/- mice found a significant discrepancy in osteoclast number, with the higher number of TRAP-positive osteoclasts conflicting with the lower number of cathepsin K-positive osteoclasts. Moreover, TUNEL staining identified a significantly higher rate of osteoclast apoptosis in IL-6-/- mice as compared with their WT controls. IL-6 deficiency induced abundant TRAP-positive osteoclasts but delayed bone remodeling by significantly inhibiting the bone resorption activity of osteoclasts and promoting osteoclast apoptosis.
Introduction
Interleukin-6 (IL-6), one of the most widely studied cytokines of the IL-6 family, is multifunctional and is involved in the regulation of various physiological processes, including immune response, acute-phase reaction, hematopoiesis, and cancer (Hirano et al. 1989). More recent studies have reported that IL-6 also functions in the maintenance of bone homeostasis; however, evidence for this remains controversial (Blanchard et al. 2009; Terpos et al. 2011; Jones et al. 2013; Ni et al. 2014). Both osteoblasts and osteoclasts are influenced by IL-6 family cytokines via signaling of glycoprotein 130 (gp130), a signal transmembrane transducer (Sims and Walsh 2010). IL-6 is widely accepted to stimulate osteoclasts. This is based on an abundance of previous reports that have shown IL-6 to induce the differentiation of osteoclast precursor cells into mature and active osteoclasts, both in vitro and in vivo (Ishimi et al. 1990; Binkley et al. 1994; Naka et al. 2002; Hong et al. 2007). However, other reports describe how IL-6 plays an inhibitory role in osteoclast formation and bone resorption by down-regulating enzymatic expression of osteoclasts and inhibiting RANKL signaling pathways (Duplomb et al. 2008; Yoshitake et al. 2008). Meanwhile, osteoblasts are found to express IL-6 receptor (IL-6R), which, when bound to IL-6, could promote osteoblast differentiation by increasing the levels of alkaline phosphatase (ALP) and osteocalcin, resulting in enhanced bone nodule formation and extracellular matrix mineralization in vitro (Nishimura et al. 1998; Heymann and Rousselle 2000; Erices et al. 2002; Franchimont et al. 2005). However, Kaneshiro et al. (In Press) found that IL-6 and its soluble receptor (sIL-6R) were able to obviously reduce the activity of osteoblasts, as well as the degree of mineralization in a dose-dependent manner in vitro, which indicates that IL-6 has negative effects on osteoblast differentiation. Taken together, these results suggest that the exact effect of IL-6 on osteoblasts and osteoclasts remains an enigma.
The role of IL-6 in bone remodeling in vivo has been studied using various transgenic and knockout animal models. In one IL-6-deficient model, generated by gene targeting, mice were reported to have a normal amount of trabecular bone, with no changes in osteoblast and osteoclast cell numbers but with a higher rate of bone turnover than the wild-type control (Poli et al. 1994). Conversely, other studies have reported that IL-6-deficient mice display increased bone formation (Heymann and Rousselle 2000; Franchimont et al. 2005). In addition, De Benedetti et al. (2006) found osteopenia in prepubertal IL-6 transgenic mice, with severe alterations in the cortical and trabecular bone microarchitecture, as well as decreased osteoblast and increased osteoclast cell numbers and activity. Furthermore, researchers have shown that an absence of IL-6 signaling results in a lower tissue mineral density, bone volume fraction, mineral/matrix ratio, and crystallinity of bone (Yang et al. 2007).
Because of the diversity and contradictory data of previous studies, the majority of preceding IL-6 knockout studies have been performed using young mice (6–8 weeks); however, limited information is available concerning the different immune responses in middle-aged (8–12 months) mice. Therefore, we aimed to collect histological evidence of bone remodeling influenced by IL-6 deficiency using a middle-aged IL-6-/- murine model.
Materials & Methods
Animals and Tissue Preparation
All animal experiments in this study were conducted according to the Guidelines for Animal Experimentation of Shandong University and Hokkaido University. Eight-month-old, male, strain-matched, IL-6-/- mice and their wild type (WT) littermates (Jackson Laboratory, Bar Harbor, ME) were used in these experiments. IL-6-/- mice were backcrossed onto the C57BL/6 background for 10 generations before being intercrossed to produce homozygotes. For statistical analysis, eight animals were present in each group. All mice were subjected to transcardiac perfusion with 4% paraformaldehyde in a 0.1 M phosphate buffer (pH 7.4). Tibiae were removed and immersed in the same fixative for an additional 12 hr. Samples were then decalcified with 10% EDTA-2Na solution for 3 weeks at 4C. Specimens were then dehydrated through an ascending ethanol series and embedded in paraffin using standard procedures. Serial sections (5-µm thickness) were prepared for histochemical analysis.
Histological Examination and Image Analysis
Hematoxylin and eosin staining was performed to investigate the morphology of the metaphyses in both groups. Stained sections were observed and digital images were taken with a light microscope (Olympus BX-53, Tokyo, Japan). Using Image Pro Plus 6.2 software (Media Cybernetics, Silver Spring, MD, USA), trabecular bone volume (BV/TV, bone volume/tissue volume × 100%) and cartilage bone volume (CV/TV, cartilage volume/tissue volume × 100%) were measured. Ten slices for each sample were used for quantitative histomorphometric analysis to obtain the mean value.
Immunohistochemistry Examination and Image Analysis
Sections were dewaxed in xylene and rehydrated in a graded ethanol series. Endogenous peroxidases were blocked by incubating sections in 0.3% hydrogen peroxide for 30 min at room temperature. Sections were then pre-incubated with 1% bovine serum albumin in phosphate-buffered saline (BSA-PBS) for 20 min to reduce non-specific staining. Following this, sections were incubated for 2 hr at room temperature with a rabbit antiserum against ALP, generated by Oda et al. (1999), at a dilution of 1:150, or an anti-cathepsin K antibody (Abcam, Cambridge, MA) at a dilution of 1:200. After rinsing with PBS, sections were incubated with horseradish peroxidase (HRP)-conjugated secondary antibodies (DaKo, Glostrup, Denmark) at a dilution of 1:100 for 1 hr at room temperature. The immunoreaction was visualized with diaminobenzidine (Sigma-Aldrich, St. Louis, MO). Double staining of ALP and TRAP was performed as previously reported (Li et al. 2013). In brief, after satisfactory immunostaining, sections treated with the ALP antibody were rinsed with PBS and submerged in a mixture of 3.0 mg of naphthol AS-BI phosphate, 18 mg of red violet LB salt, and 100 mM L(+) tartaric acid (0.36 g), diluted in 30 mL of 0.1 M sodium acetate buffer (pH 5.0), for 15 min at 37C. Staining was observed by light microscopy (Olympus) after faintly counterstaining with methyl green. The primary antibody was replaced with 1× PBS for the negative control.
Ten tissue sections were selected for each sample group for the statistical assessment of osteoclast number and ALP activity. This was conducted in three randomly selected, non-overlapping microscopic fields of view, using Image Pro Plus 6.2 software. Additionally, cathepsin K-positive osteoclasts and TRAP-positive osteoclasts were counted in the same serial section views at an original magnification of ×1000.
TUNEL Detection and Quantification
For the detection of apoptotic cells, the TACS 2TdT-Blue Label In Situ Apoptosis Detection Kit (Trevigen Inc., Gaithersburg, MD), for terminal deoxynucleotidyl transferase-mediated deoxyuridine triphosphate nick end labeling (TUNEL), was employed. Dewaxed sections were incubated with 1% proteinase K (Trevigen Inc.) diluted 1:200 at 37C for 15 min, followed by inhibition of the endogenous peroxidases at room temperature for 5 min. After treatment with TdT enzyme at a dilution of 1:50 at 37C for 1 hr, sections were incubated with HRP-conjugated streptavidin at room temperature for 15 min. The reaction was made visible with the blue label solution provided in the kit. Apoptotic osteoclasts were determined as both TUNEL- and TRAP-positive in the same field of serial sections at an original magnification of ×1000.
Statistical Analysis
All statistical analyses were performed using SPSS software (Chicago, IL) and all values are presented as the mean ± standard deviation (SD). Differences among groups were assessed by the unpaired t-test and considered statistically significant when p<0.01.
Results
Histological Alterations of the Tibial Metaphysis
IL-6-/- mice showed significantly more trabecular bone and a wider growth plate than their WT controls (Fig. 1A, 1B). Moreover, abundant residual cartilage cores were seen in the metaphyses of IL-6-/- mice as compared with WT counterparts (Fig. 1C, 1D). As shown in Fig. 1E-1G, statistical analysis revealed significant differences (p<0.001) between the IL-6-/- group and their WT controls with regard to trabecular bone volume (41.10 ± 0.68 vs 14.82 ± 0.30), cartilage volume (14.11 ± 0.13 vs 2.04 ± 0.03), and height of growth plate (98.63 ± 0.85 vs 58.86 ± 0.36), respectively.
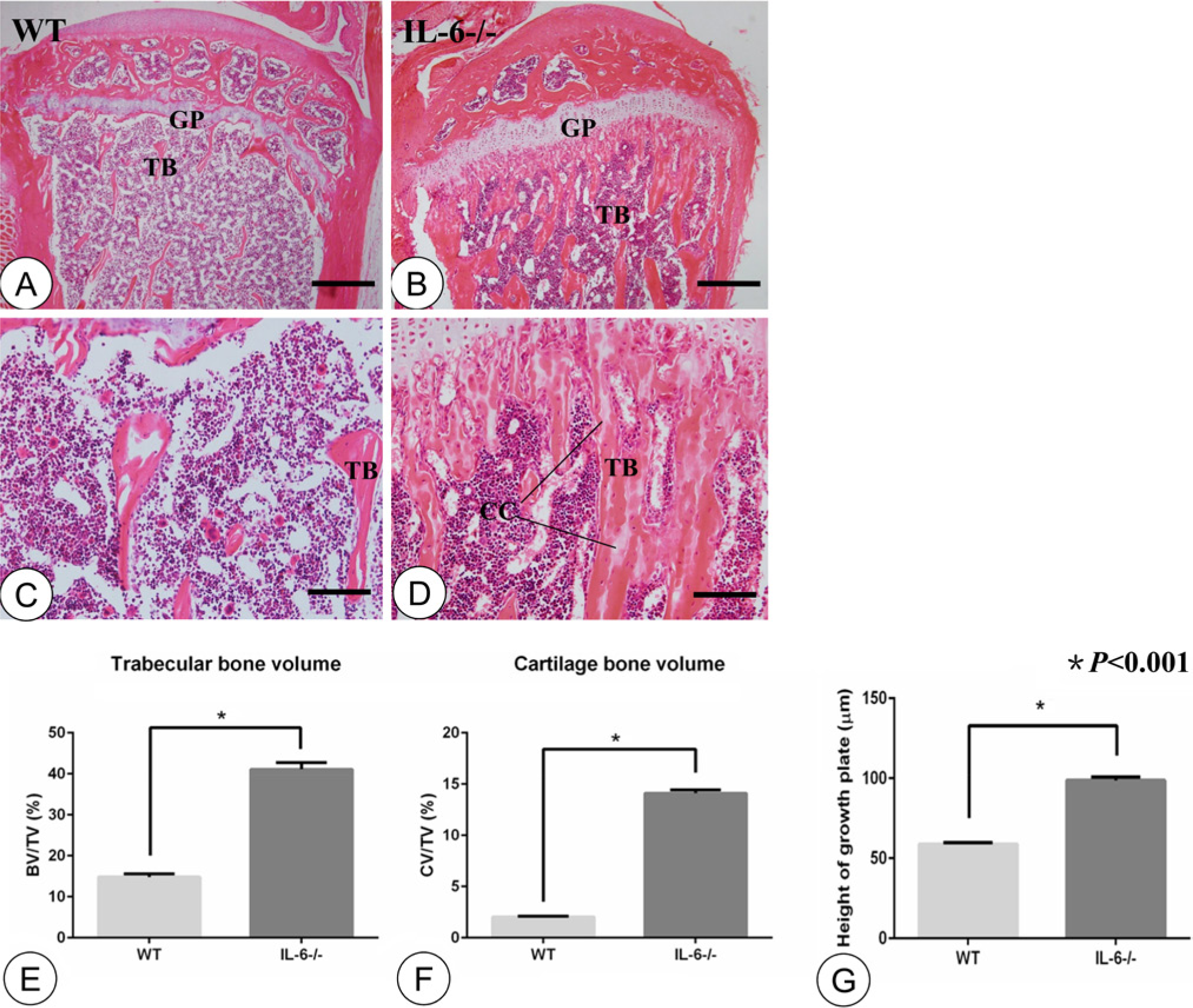
Hematoxylin-eosin staining and statistical analysis. Low-magnification images of the tibial metaphysis in the WT (A) and IL-6-/- (B) groups. High-magnification images of the tibial metaphysis in the WT (C) and IL-6-/- (D) groups. Statistical analysis of trabecular bone volume (E), cartilage bone volume (F), and height of growth plate (G) between the groups. The WT group manifested scarce trabeculae and a thin growth plate (A, C). The IL-6-/- group showed more immature trabecular bone containing abundant residual cartilage cores and an abnormal growth plate (B, D). Statistical analysis revealed significant differences in trabecular bone volume, cartilage volume as well as height of growth plates between the WT and IL-6-/- groups (E, F, G; p<0.001, n=8). TB: trabecular bone; GP: growth plate; CC: cartilage cores. Error bars indicate ± SD. Scale bars in A and B, 300 µm; C and D, 60 µm.
Effects of IL-6 Deficiency on Osteoblasts and Osteoclasts
As shown in Figure 2, ALP was faintly expressed and only a few TRAP-positive osteoclasts were found in the WT group (Fig. 2A). In contrast, ALP activity was significantly enhanced and abundant TRAP-positive osteoclasts were found on the surface of the trabecular bone in the metaphyses of the IL-6-/- group (Fig. 2B). Both ALP activity (0.30 ± 0.003 vs 0.19 ± 0.004) and TRAP-positive osteoclast cell number (60.3 3± 1.09 vs 10.50 ± 0.42) were significantly (p<0.001) increased in the IL-6-/- mice (Fig. 2C, 2D).
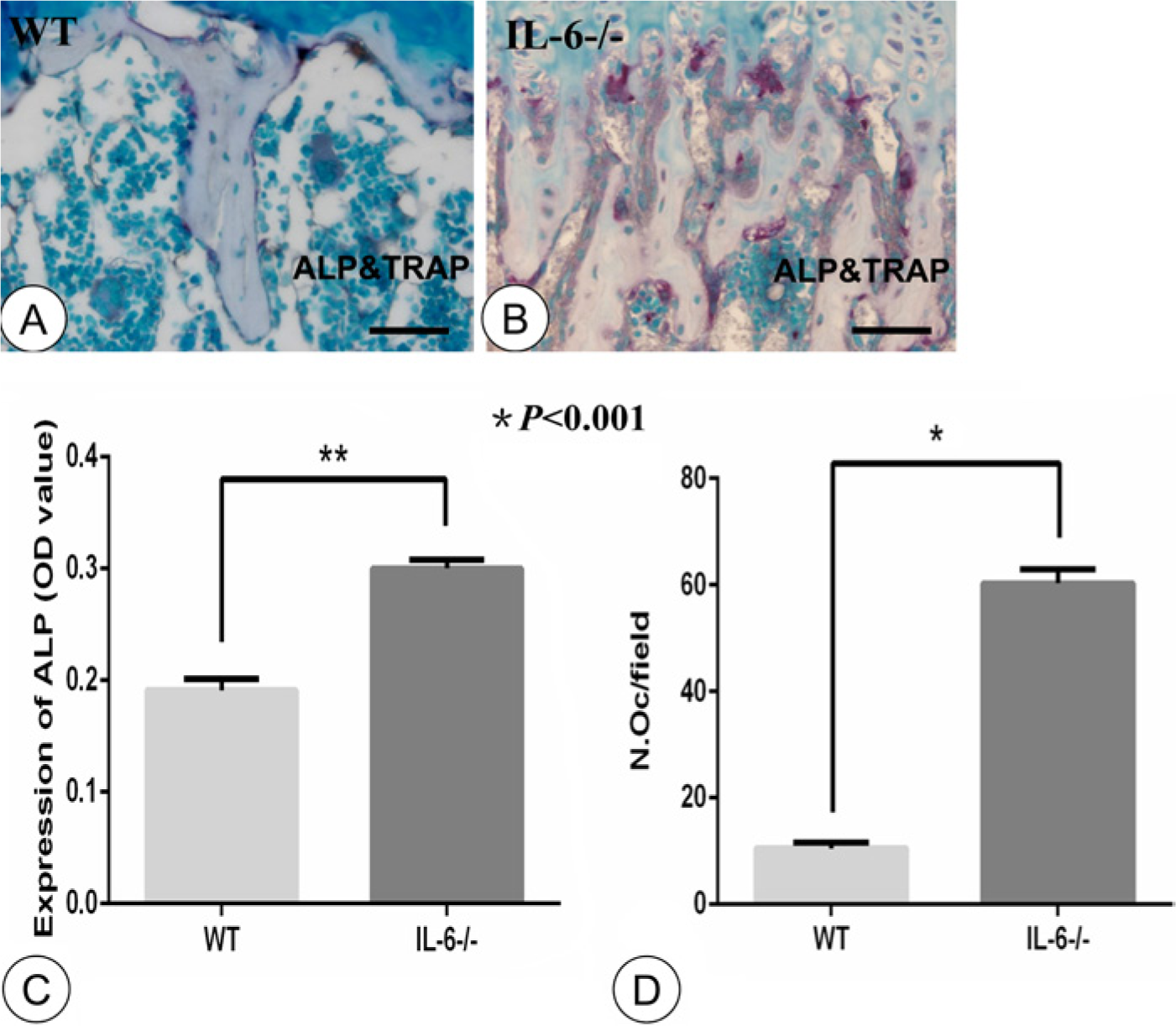
Double-staining of alkaline phosphatase (ALP) and tartrate resistant acid phosphatase (TRAP) and statistical analysis. Double staining for ALP (brown) and TRAP (red) in trabecular bone of the WT (A) and IL-6-/- (B) groups. Statistical analysis for expression of ALP- (C) and TRAP-positive osteoclasts (D). ALP was faintly expressed and few TRAP-positive osteoclasts were observed in the WT group (A). Strong ALP and abundant TRAP-positive osteoclasts were observed in the IL-6-/- group (B). Expression of ALP- and TRAP-positive osteoclast numbers were significantly increased in the IL-6-/- group (C, D; n=8, p<0.001). OD: optical density; OC: osteoclast. N.Oc, number of osteoclasts. Error bars indicate ± SD. Scale bars in A and B, 30 µm.
Discrepancy between the Number of TRAP-positive and Cathepsin K-positive Osteoclasts
The numbers of TRAP-positive and cathepsin K-positive osteoclasts in the same microscopic visual fields were consistent in the WT group (Fig. 3A, 3C), but obvious differences in osteoclast positivity were observed for sections from the IL-6-/- mice, as shown in Figure 3B and 3D. Statistically, the numbers of TRAP-positive and cathepsin K-positive osteoclasts were proportionate in the WT control group (5.5 ± 1.08 vs 5.45 ± 1.05, p>0.05, Fig. 3E), but an obvious discrepancy between these two markers was measured for samples taken from IL-6-/- mice (16.5 ± 1.05 vs 6.17 ± 0.75, p<0.001; Fig. 3E).
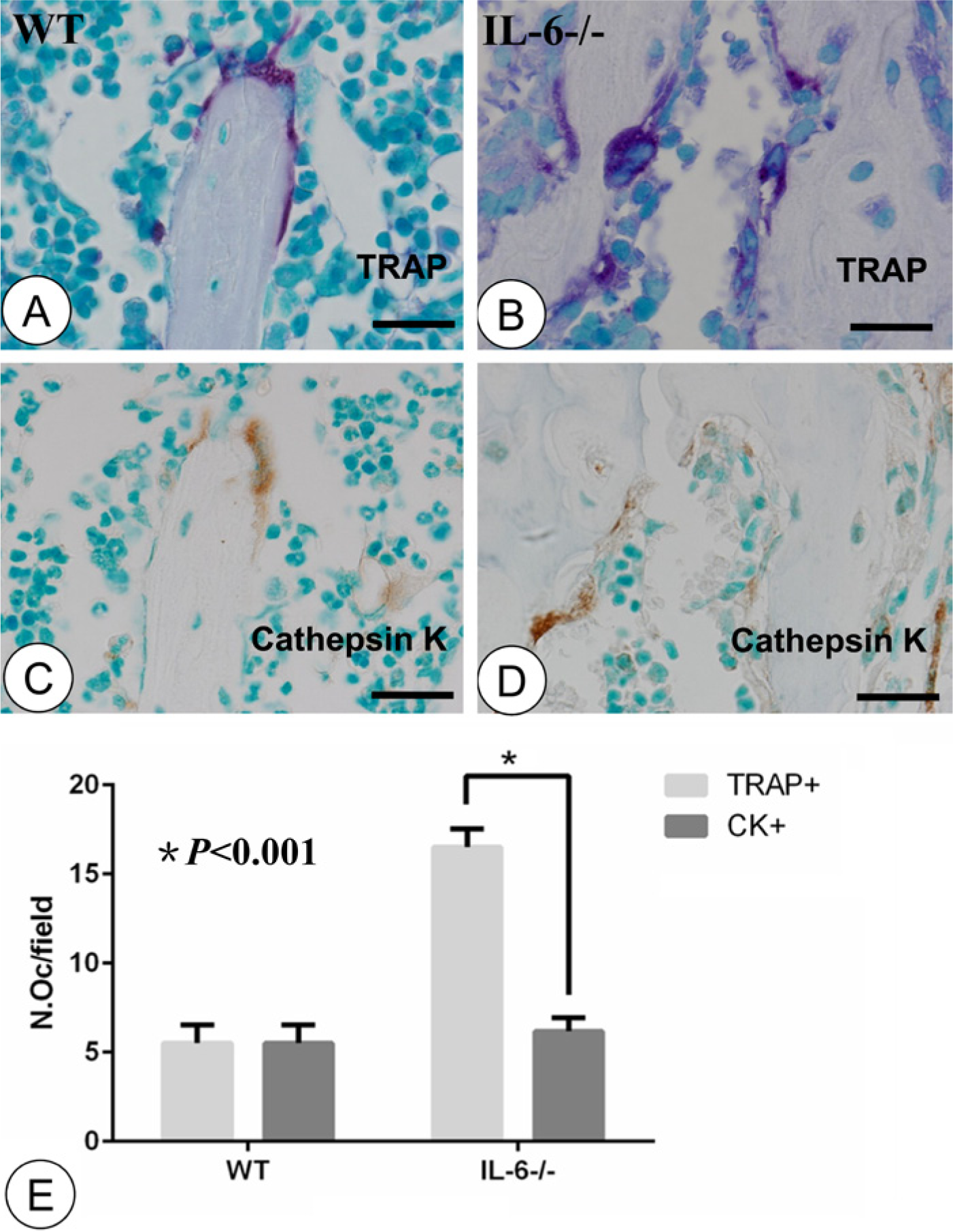
Staining for TRAP and cathepsin K and statistical analysis in serial sections. TRAP-positive osteoclasts (red) in the WT (A) and IL-6-/- (B) groups. Immunolocalization of cathepsin K (brown) in microscopically identical visual fields in the WT (C) and IL-6-/- (D) groups. (E) Statistical analysis for TRAP-positive and cathepsin K-positive osteoclasts. WT group showed consistent expression of TRAP-positive and cathepsin K-positive osteoclasts in the same microscopic visual fields (A, C). In the IL-6-/- group, many TRAP-positive osteoclasts showed faint or undetectable cathepsin K activity (B, D). There was an obvious discrepancy between the TRAP-positive and cathepsin K-positive osteoclast numbers in the IL-6-/- group (E, n=8, p<0.001). OC, osteoclast; CK, cathepsin K; N.Oc, number of osteoclasts. Error bars indicate ± SD. Scale bars, 12 µm.
Effects of IL-6 Deficiency on Osteoclast Apoptosis
TUNEL staining was used to detect osteoclast apoptosis in different groups in identical microscopic fields of view. TRAP staining was used to visualize osteoclasts whereas TUNEL staining was applied to detect apoptotic cells on serial sections. Very few visible apoptotic osteoclasts were observed in the WT group (Fig. 4A, 4C). However, the number of apoptotic cells was significantly increased in the IL-6-/- group, especially at sites where TRAP-positive osteoclasts were located (Fig. 4B, 4D). Statistically, IL-6 deficiency caused an increase in the proportion of apoptotic osteoclasts (8.51 ± 0.56 in WT group vs 49.95 ± 0.37in IL-6-/- group, p<0.001; Fig. 4E).
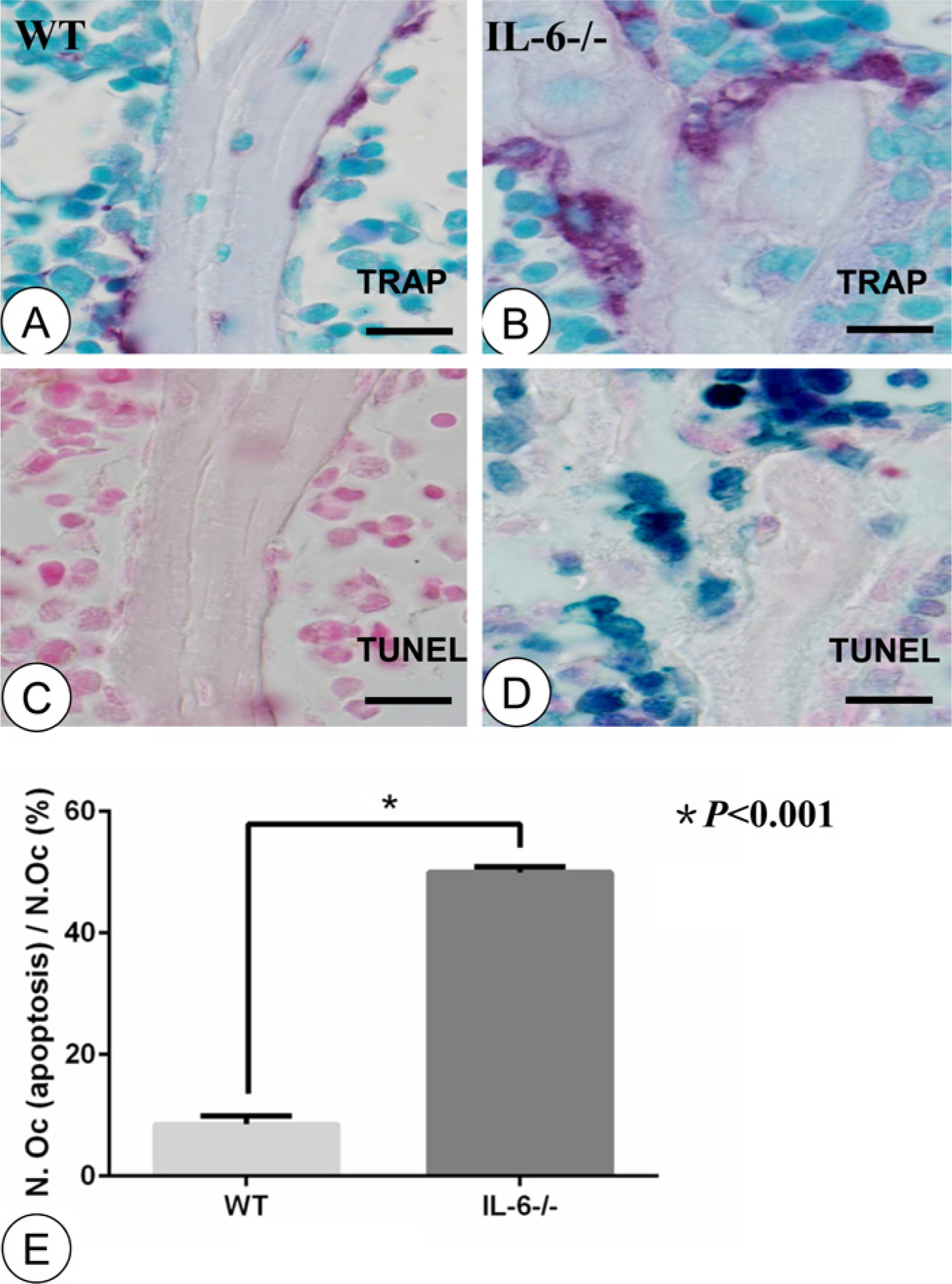
Staining of TRAP and TUNEL on serial sections. TRAP-positive osteoclasts (red) in the WT (A) and IL-6-/- (B) groups. TUNEL detection (dark blue) revealed osteoclast apoptosis in the WT (C) and IL-6-/- (D) groups. (E) Statistical analysis comparing the percentage of apoptotic osteoclasts. The WT group showed few TRAP-positive osteoclasts on the surface of trabecular bone (A) with numerous in the IL-6-/- group (B). There were few visible apoptotic bodies in the microscopic vision of the WT group (C). The IL-6-/- group displayed abundant apoptotic bodies, especially at sites of TRAP-positive osteoclasts (D). Statistical analysis revealed a significant difference in apoptotic bodies between the WT and IL-6-/- groups (E; n=8, p<0.001). Error bars indicate ± SD. Scale bars, 12 µm.
Discussion
In this study, we investigated bone remodeling in the metaphysis of the tibia using an 8-month-old IL-6-/- murine model. Previous studies report that young (aged 6–8 weeks) IL-6-/- mice have no significant bone abnormalities (Kopf et al. 1994; Sims et al. 2004) or only a higher rate of bone turnover than WT controls (Bonewald 2007). In this study, we found that middle-aged IL-6-deficient mice had a more immature bone structure than WT controls in the metaphysis of the tibia, which was characterized by thicker trabeculae containing abundant residual cartilage cores, as well as a wider growth plate. Previous studies have found that plasma IL-6 concentration was higher in middle-aged subjects than in younger ones, both in mice and humans (Wei et al. 1992; Roubenoff et al. 1998; Okamura et al. 2012). Moreover, an age-related osteoporosis animal model has shown decreased bone mineral density accompanied by increased IL-6 (Halade et al. 2010). Given these data, we speculated that the inconformity between our results and that of previous studies may be contributed to age.
To further explore the underling mechanism of how IL-6 deficiency influences bone remodeling, we performed double staining of ALP and TRAP and found that IL-6 deficiency enhanced ALP activity on the surface of the trabecular bone. This finding is consistent with previous reports that IL-6 depletion can enhance osteoblast proliferation and osteogenic capability (Yang et al. 2007), and that genetically induced overexpression of IL-6 in two transgenic mouse models led to a significant reduction in osteoblast number (Kitamura et al. 1995; De Benedetti et al. 2006). Meanwhile, we found an increase in osteoclast number in IL-6-deficient mice, which is inconsistent with the generally accepted theory that IL-6 has a positive effect on osteoclasts (Ishimi et al. 1990; Binkley et al. 1994; Yokota et al. 2014). However, subsequent studies have revealed that IL-6 primarily influences osteoclasts indirectly through RANKL production by osteoblasts, and this indirect IL-6 effect is substantially larger than the effect IL-6 exerts directly on osteoclast precursors (Udagawa et al. 1995, Johnson et al. In Press). As demonstrated in the schema (Fig. 5), we speculate that IL-6 deletion causes an increase in osteoblast numbers and thus an increase in RANKL secretion by these cells to promote osteoclast differentiation. More importantly, this indirect effect of IL-6 deletion far exceeds the direct inhibitory function on osteoclasts. In addition, the role IL-6 plays in osteoclastogenesis can be compensated for by other factors, such as TNF-α, monocyte chemotactic protein-1, and IL-1 (Poli et al. 1994), which may provide another possible mechanism to explain the increase in osteoclast number. However, the precise mechanism for this requires further study.
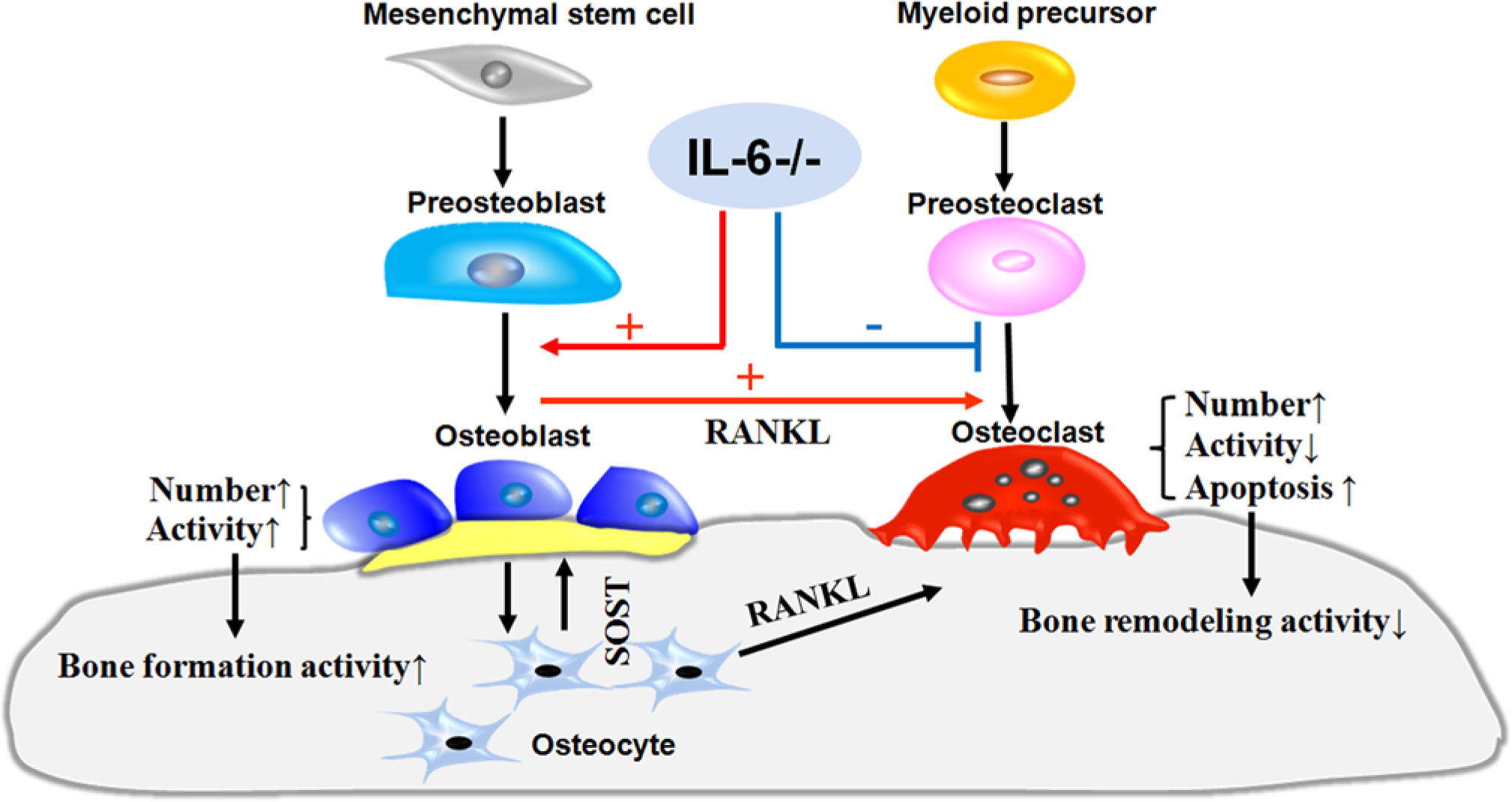
A hypothetical scheme illustrating the influence of IL-6 deficiency on bone remodeling. IL-6 deficiency likely regulates the differentiation of osteoclasts by direct inhibition (blue line) and indirectly by promoting (red line) osteoclastogenesis.
The glycosylated metalloenzyme TRAP is highly and specifically expressed in osteoclasts and is widely used as a specific bone osteoclast marker (Minkin 1982). Contrastingly, the cysteine proteinase cathepsin K is selectively expressed in osteoclasts (Drake et al. 1996). Although several studies indicate both TRAP and cathepsin K are involved in bone degradation, TRAP has been shown to be present in osteoclasts in an inactive pro-form that can be activated by certain proteinases, particularly cathepsin K (Ljusberg et al. 1999a; Ljusberg et al. 1999b). We therefore used TRAP as a specific marker of osteoclasts whereas cathepsin K was used to indicate osteoclastic activity. To explain delayed bone remodeling, as signified by abundant residual cartilage cores and an abnormal growth plate in the metaphysis of IL-6-deficient mice, we detected osteoclast activity using TRAP and cathepsin K assays. Our results found an obvious discrepancy between the number and bone resorption ability of osteoclasts in the IL-6-/- group. That is, more TRAP-positive and less cathepsin K-positive osteoclasts were seen in the same visual field, as shown in Figure 3. This phenomenon can be partially explained by previous studies, whereby the combination of IL-6 and its soluble receptor are reported to stimulate bone resorption activity of mature osteoclasts in vitro (Adebanjo et al. 1998; Palmqvist et al. 2002) and in vivo (Binkley et al. 1994). However, further examination of the osteoclast ruffled border ultrastructure and enzymatic immunocompetence are necessary in future studies. In addition, our results showed many TRAP-positive osteoclasts were apoptotic in the IL-6-/- group, indicating that IL-6 deficiency had a negative effect on osteoclast survival. In view of the capacity of IL-6 to delay osteoclast apoptosis, as reported in previous studies (Yonish-Rouach et al. 1991; Schwarze et al. 1995), our findings are consistent with the well-recognized, anti-apoptotic properties of IL-6.
In the current study, IL-6 depletion increased TRAP-positive osteoclast numbers, many of which showed faint bone resorption activity and characteristics of apoptosis, resulting in reduced bone resorption ability and delayed bone remodeling. However, the presence of these TRAP-positive cells is enough to couple bone formation (Soriano et al. 1991). Considering the significant osteogenic capability and low bone resorption activity, it is not surprising that IL-6-/- mice displayed thicker trabeculae and a more immature bone structure, characterized by abundant residual cartilage cores and a wider growth plate, than their WT controls. In summary, our study suggests that IL-6 deficiency increases TRAP-positive osteoclast numbers, but delays bone remodeling by significantly inhibiting the bone resorption ability and promoting apoptosis of osteoclasts.
Footnotes
Declaration of Conflicting Interests
The authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The authors disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This study was partially supported by the