
Abstract
Mycobacterium avium subsp. paratuberculosis is the causative agent of Johne's disease, a chronic granulomatous enteritis of wild and domestic ruminants. During a long subclinical period, the organism persists in the intestine despite systemic cellular and humoral immune responses. To explore the mucosal immune response in Johne's disease, we isolated mononuclear leukocytes from the ileum of cows naturally infected with M. avium subsp. paratuberculosis and from cows that were not infected. We evaluated the immunophenotype of these cells and the proliferative responses after the addition of M. avium subsp. paratuberculosis sonicate or B-cell or T-cell mitogens. Although the percentage of T cells was increased in infected cows, these cells consisted mostly of memory (CD2+CD62L−) and regulatory (CD4+CD25+) T cells. Further evidence of immune hyporesponsiveness included a decrease in the percentage of T cells with an activated phenotype and a decrease in cells expressing major histocompatibility factor class II (MHC class II). Unlike the spleen, ileal lymphocytes from infected cows failed to proliferate in response to M. avium subsp. paratuberculosis sonicate. Additionally, ileal lymphocytes from infected cows proliferated poorly in response to concanavalin A and pokeweed mitogen, suggesting generalized T cell and B cell hyporesponsiveness. These results indicate that a state of tolerance may exist in the intestine of cows subclinically infected with M. avium subsp. paratuberculosis organisms in subclinically infected cows. This effect may be induced, at least in part, by proliferation of regulatory T cells that nonspecifically suppress mucosal immune responsiveness.
Mycobacterium avium subsp. paratuberculosis is the causative agent of Johne's disease, a chronic granulomatous enteritis of wild and domestic ruminants. 9, 13, 27 Mycobacterium avium subsp. paratuberculosis is a slow-growing facultative intracellular bacterium that persists within macrophages in the intestinal tract for several years before the onset of clinical disease. 13, 27 Clinical disease is associated with granulomatous enteritis that is usually restricted to the ileum. 9
The pathogenesis of Johne's disease is poorly understood. The organism is endocytosed by M cells of the ileal Peyer's patches and is subsequently phagocytized by macrophages. 25 Lesions, in both naturally occurring and experimental disease, develop in the intestinal wall 1–3 months after infection and are characterized by aggregates of macrophages, epithelioid cells, and giant cells. 5, 21, 28 Systemic humoral and cellular immunity develop over 1–4 months after experimental infection. 31
Persistence of the organism in the intestinal lamina propria in the presence of a systemic immune response suggests that the immune response in the intestine may be fundamentally different from the systemic response. Results of recent studies in mice suggest that Peyer's patches may provide a permissive microenvironment for induction of tolerance to commensal flora and food antigens. 4, 16, 23 Tolerance appears to be mediated by several types of regulatory T cells including CD4+CD25+ regulatory T cells, type 1 regulatory T cells (Tr1), and type 2 regulatory T cells (Tr2, also called Th3 T cells). 14, 16 It is presently uncertain if Tr1 and Tr2 cells are CD25+. 14, 16
In the present study, we hypothesized that persistence of M. avium subsp. paratuberculosis in the intestinal mucosa was the result of proliferation of regulatory T cells that suppress Th1-type immune responses. To test this hypothesis, we isolated mononuclear leukocytes from the intestinal lamina propria of cows with subclinical Johne's disease and of uninfected cows and determined the immunophenotype of the cells and proliferative responses after addition of M. avium subsp. paratuberculosis sonicate or B-cell and T-cell mitogens.
Materials and Methods
Bacteria
M. avium subsp. paratuberculosis strain 19698 was obtained from the American Type Culture Collection. This strain was isolated from a naturally infected cow. The organisms were grown in 7H9 broth containing oleic acid–albumin–dextrose–catalase (OADC; Difco Laboratories, Detroit, MI), Tween 80, and mycobactin J (Allied Laboratories, Ames, IA). M. avium subsp. paratuberculosis was grown to an optical density reading at 540 nm of 0.2–0.4. The organisms were pelleted (10,000 × g for 20 minutes) and washed twice in cold Dulbecco's phosphate buffered saline (DPBS). The pellet was resuspended in phosphate-buffered serum (PBS) and sonicated with a probe sonicator. 30 Sonication consisted of three 10-minute cycles at 18 W on ice. Sonicated samples were centrifuged at 12,000 × g to remove debris, and the supernatant was harvested and stored at −20°C. Protein concentration was determined by use of a microprotein assay (Bio-Rad Laboratories, Richmond, CA).
Cows
Eight adult Holstein cows were used in the study. Four cows had at least three positive fecal cultures for M. avium subsp. paratuberculosis and were classified as high fecal shedders (i.e., >100 colonies/g of feces). Four cows were found to have negative fecal cultures on at least four occasions. The average age of infected cows was 3.7 years, and the average age of the uninfected cows was 4.4 years. None of the cows were clinically ill at the time of purchase. All cows were tested for M. avium subsp. paratuberculosis infection by use of fecal culture, serum ELISA test (IDEXX Laboratories, Inc. Westbrook, Maine), and cell-mediated immune responses (Bovigam Kit, CSL Limited, Victoria, Australia). After euthanasia, multiple pieces if ileal tissue were submitted to the Minnesota Veterinary Diagnostic Laboratory for mycobacterial culture and for M. avium subsp. paratuberculosis PCR testing. 11 All tests were positive for infected cows and all tests were negative for uninfected cows.
Experimental design
Cows were euthanatized with an overdose of barbiturate, and a 20-cm piece of terminal ileum and an approximately 20-g piece of spleen were removed within 15 minutes. The piece of ileum was turned inside out and flushed with ice-cold, sterile, calcium- and magnesium-free DPBS. Distinct Peyer's patches could not be visualized in either infected or uninfected cows probably because ileal Peyer's patches normally involute at a young age in cattle. 20 Therefore, the mucosa was scraped off the entire piece of ileum as described. 6, 26 The tissue was washed three times in ice cold DPBS. 6 Spleen was cut into small pieces. Mucosal and splenic samples were incubated with 2-mM ethylenediaminetetraacetic acid (EDTA) at 37°C for 1.5 hours with constant stirring, washed, and incubated with collagenase (80 IU/ml; Sigma Chemical Co., St. Louis, MO) and deoxyribonuclease type V (0.1 mg/ml; Sigma Chemical Co, St. Louis, MO) at 37°C for 1 hour. Thereafter, cells were pelleted, washed 3 times, and filtered through glass wool to remove particulate matter and dead cells. Mononuclear cells were isolated by centrifugation over a 50% Percoll density gradient at 500 × g for 30 minutes. Cell viability was determined by Trypan blue exclusion. All cell suspensions had 85% or greater live cells. Smears were prepared from the ileal and splenic cell suspensions and were stained with Wright's-Giemsa stain. Five hundred cell-differential cells counts were performed by use of light microscopy. The immunophenotype of the cell populations and proliferative responses to M. avium subsp. paratuberculosis sonicate and B-cell and T-cell mitogens were determined.
Immunophenotyping mononuclear leukocytes
The immunophenotype of mucosal and splenic mononuclear cells was determined by one- or two-color flow cytometry. 35 Five hundred thousand viable cells were placed in each of 12 tubes. Cells were incubated with fluorescein isothiocyanate (FITC)-labeled CD2, anti-CD8, FITC-labeled anti-CD4, anti-CD11b, phycoerythrin (RPE)-Cy5-labeled anti-CD14, anti-CD21, FITC-labeled anti-CD2 and anti-CD62L, FITC-labeled anti-CD4 and anti-CD25, anti–major histocompatibility factor class II (MHC class II), anti–B cell and activated T cell, or anti–γδT cell receptor (Table 1). Protein concentration of each antibody was adjusted to approximately 10 µg/ml. Cells were displayed as forward-angle versus side-angle light scatter plots (Fig. 1). In preliminary studies, gates were set to identify lymphocytes, macrophages, and neutrophils (Table 1, 2). The macrophage population was identified as cells that labeled with anti-CD11b and anti-CD14. The lymphocyte population was identified as cells that labeled with anti-CD2 or anti-CD21. The neutrophil population was identified as cells that labeled with anti-CD11b but not with anti-CD14.
Monoclonal antibodies used for immunophenotyping bovine mononuclear leukocytes.
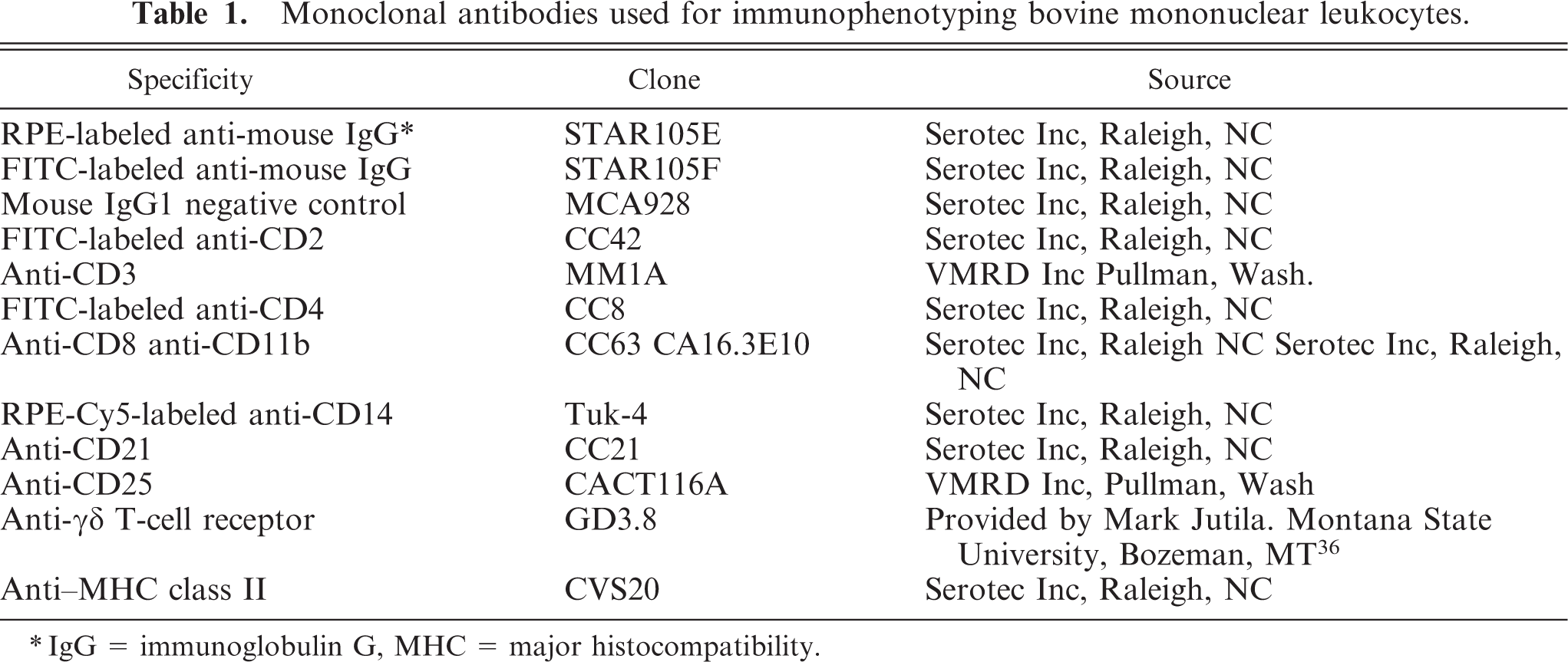
∗IgG = immunoglobulin G, MHC = major histocompatibility.
Evaluation of the purity of isolated ileal leukocyte subpopulations within flow cytometric gates (see Fig. 1). Data expressed as the percentage of cells with increased fluorescence intensity when compared with cells labeled with isotype-matched negative control serum. Data listed as mean ± SD.

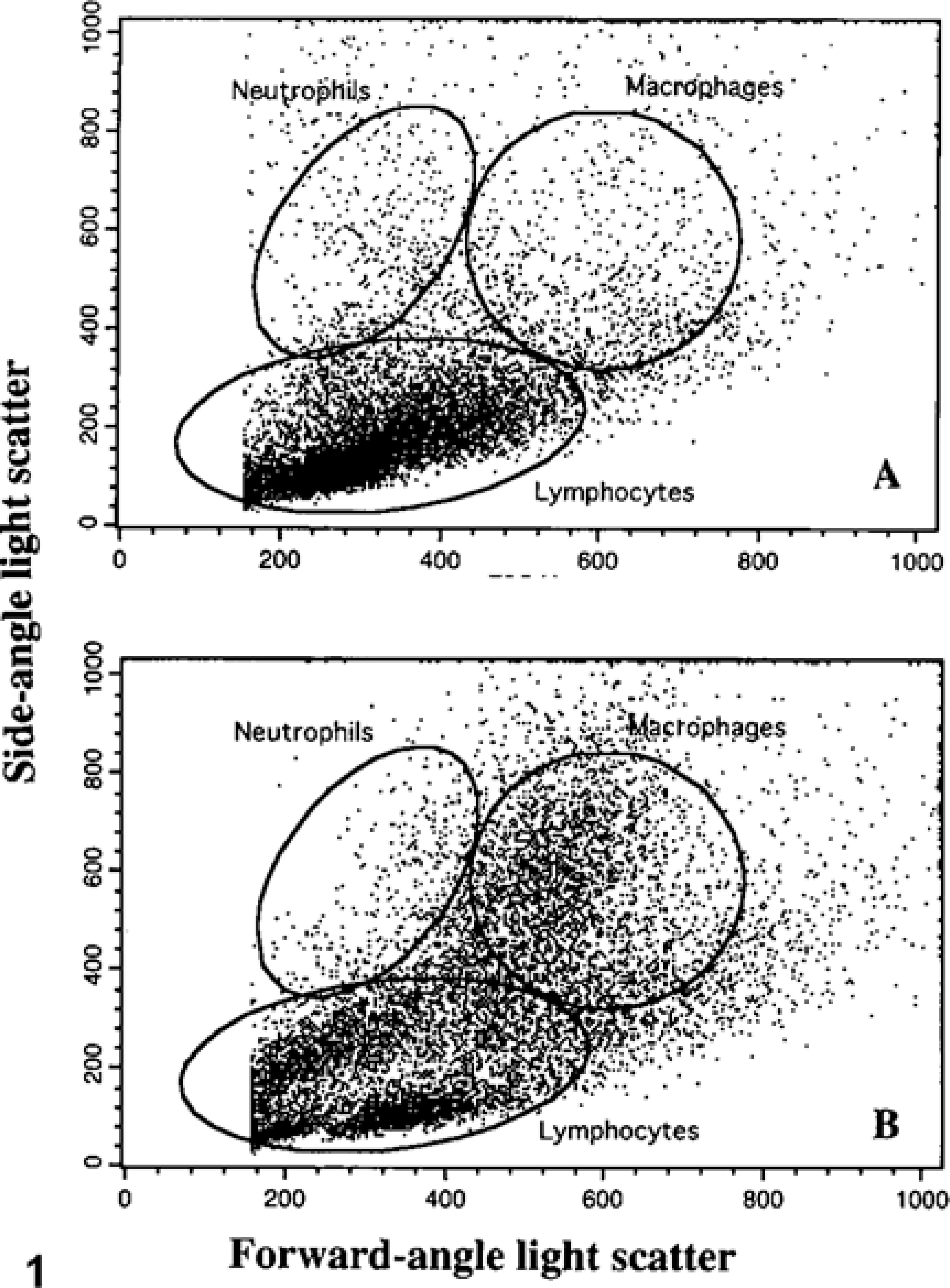
Flow-cytometric scatter plot of isolated ileal leukocytes from
The method for one-color flow cytometry consisted of incubation of cell suspensions with primary antibody for 30 minutes, washing, and incubation of cell suspensions with FITC-labeled anti-mouse immunoglobulin G (IgG) for 30 minutes. The method for two-color flow cytometry consisted of incubation with the primary antibody for 30 minutes, washing, incubation with RPE-labeled anti-mouse IgG for 30 minutes, washing, blocking with 5% mouse serum for 10 minutes, and incubation with FITC-labeled anti-bovine CD2 or CD4 for 30 minutes. 32, 36 Macrophages were identified by incubation with RPE-Cy5-labeled anti-CD14. Labeling with anti-CD3 identified T cells and labeling with anti-CD21 identified B cells. Naive (CD2+CD62L+), memory (CD2+CD62L−), activated/suppressor (CD4+CD25+), activated T cells, and γδT cells and were identified by their labeling patterns. 12, 32 Mouse IgG1 was added to negative control cells. A commercial software program (CELLQUEST, Becton Dickenson Co, Pleasanton, CA) was used to evaluate flow-cytometric data.
Cell proliferation assay
The proliferative responses of cell populations derived from the ileum and spleen were evaluated. Five hundred thousand viable lymphocytes, suspended in RPMI medium supplemented with 10% fetal bovine serum, 1 µg/ml gentamycin, 2.5 mg/ml chloramphenicol, and 10 µg/ml amphotericin B were added to 96-well microtiter plates and incubated for 3 days at 37°C, and in the presence of 4% CO2 with or without addition of M. avium subsp. paratuberculosis sonicate (10 µg /well), the T-cell mitogen concanavalin A (10 µg/ml; Sigma Chemical Co. St. Louis, MO), or the B-cell mitogen pokeweed mitogen (1 µg/ml, Sigma Chemical Co). Thereafter, the cells were frozen at −70°C until assayed. After thawing, DNA was fluorescently labeled by adding 200 µl of a green fluorescent dye/cell lysis buffer (CyQUANT GR dye, Molecular Probes Inc, Eugene, OR), and fluorescence was measured at 480-nm excitation and 520-nm emission using a fluorescence microplate reader. Negative controls consisted of mononuclear cells incubated with vehicle and dye. Results for proliferation assays were expressed as stimulation indices (SI)
6
:

Necropsy procedures and histopathologic studies
After tissues were collected for in vitro studies, a gross and histopathologic examination of the intestinal tract was performed. The ileum, jejunum, and mesenteric lymph nodes were examined for gross lesions. Tissue samples were taken from all gross lesions. Tissue samples were also be taken every 10 cm throughout the ileum and from several sites in the jejunum for histopathologic examination. All tissue sections were stained with hematoxylin and eosin (HE) and Ziehl-Neelzen acid-fast stains. 10 Thickness of the mucosa was determined in histopathologic sections by light microscopy using an ocular micrometer. Selected tissue samples were submitted for mycobacterial culture, and remaining tissue was homogenized and submitted for detection of Mycobacterium avium subsp. paratuberculosis by use of a specific PCR test. 11
Analysis of data
Data derived from four infected and four uninfected cows was analyzed. Means of duplicate or triplicate samples were analyzed by analysis of variance (ANOVA). Means of interest were compared by use of the Bonferroni-Dunn F-test. P < 0.05 was considered to be statistically significant.
Results
Necropsy and histopathology
When compared with uninfected cows, infected cows had a thicker ileal mucosa (P < 0.05). Ileal thickness in uninfected cows was 2.9 ± 0.4 mm, and ileal thickness in infected cows 4.8 ± 0.8 mm. One infected cow had enlarged mesenteric lymph nodes. Histopathologic sections from infected cows contained multifocal infiltrates of macrophages and epithelioid cells and more diffuse infiltrates of lymphocytes and plasma cells within the lamina propria. Peyer's patches were not apparent in these sections. Large numbers of acid-fast organisms were present within macrophages and epithelioid cells in all four infected cows. Uninfected cows had a sparse infiltrate of lymphocytes and plasma cells in the lamina propria, but few macrophages were observed. Peyer's patches were not observed in any sections. Acid-fast organisms were not identified.
Cell distribution in ileum and spleen
Leukocyte percentages in ileal and splenic cell isolates were evaluated by light microscopy and by flow cytometry (Fig. 1). Leukocytes from both ileum and spleen consisted mostly of lymphocytes. Ileal leukocytes from infected cows had 3.8-fold greater numbers of macrophages when compared with ileal leukocytes from uninfected cows (Table 3). Splenic cell distribution was similar between infected and uninfected cows.
Cell distribution in ileum and spleen of uninfected cows and cows subclinically infected with Mycobacterium avium subsp. paratuberculosis.
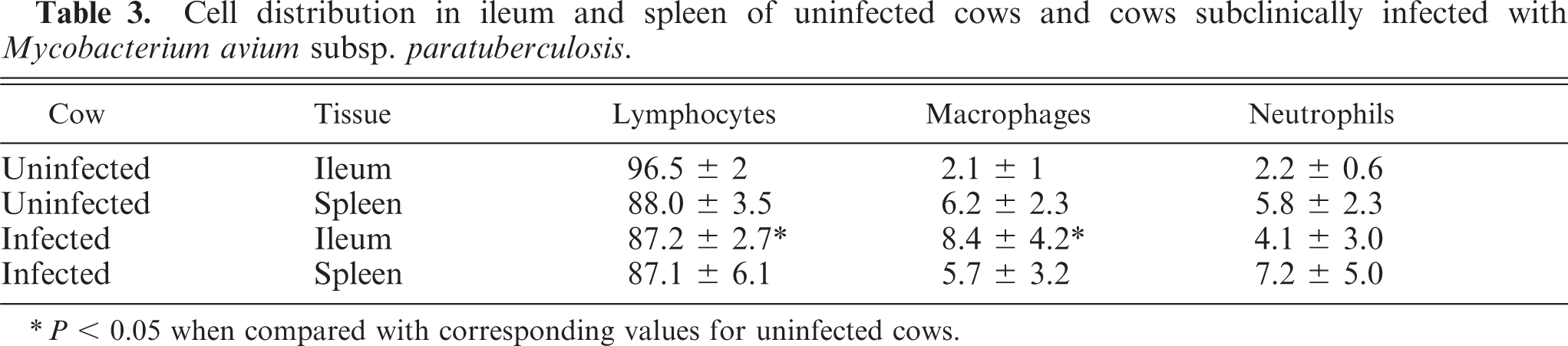
∗P < 0.05 when compared with corresponding values for uninfected cows.
Immunophenotype of the ileum
Ileal mononuclear leukocytes were labeled with a battery of monoclonal antibodies to detect functional subtypes. The immunophenotype of M. avium subsp. paratuberculosis-infected cows differed from that of uninfected cows (Table 4). The ratio of T cells to B cells for infected cows was 5.8 : 1, whereas the ratio for uninfected cows was 0.7 : 1 (P < 0.01). Infected cows had a lower percentage of MHC class II+ cells, and CD2+CD62L+ (i.e., naive) T cells (P < 0.01; Fig. 2). Infected cows had a greater percentage of CD4+CD25+ T cells (Fig. 2). Few cells in either infected or uninfected cows labeled for the γδ T cells receptor, indicating that most T cells were of the αβ type. Infected cows had a greater percentage of macrophages in the ileum compared with uninfected cows (P < 0.05).
Immunophenotype of mononuclear leukocytes from the ileum and spleen of uninfected cows and cows subclinically infected with Mycobacterium avium subsp. paratuberculosis. Results are listed as a percentage of total lymphocytes in the ileal or splenic cell populations.
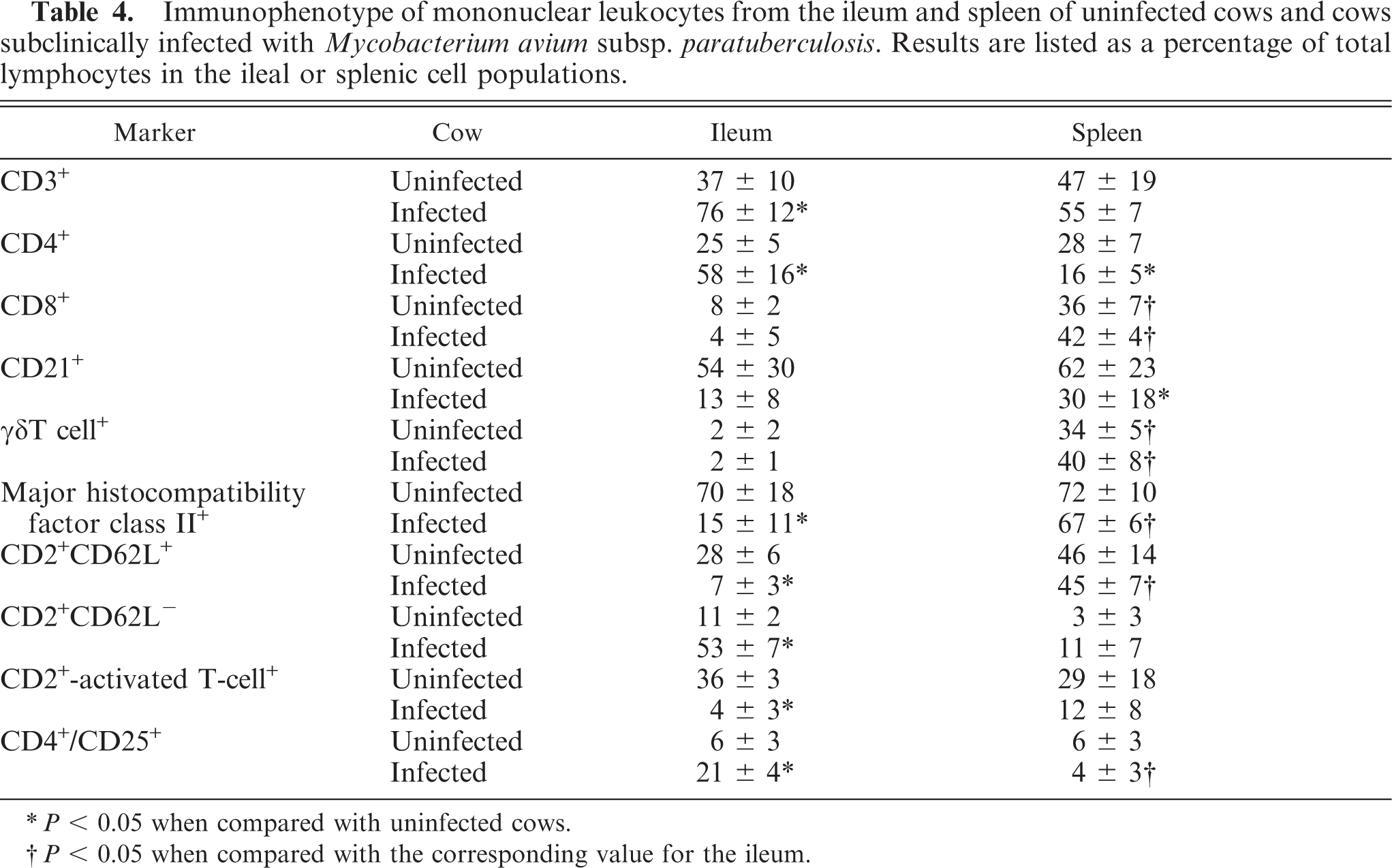
∗ P < 0.05 when compared with uninfected cows.
† P < 0.05 when compared with the corresponding value for the ileum.
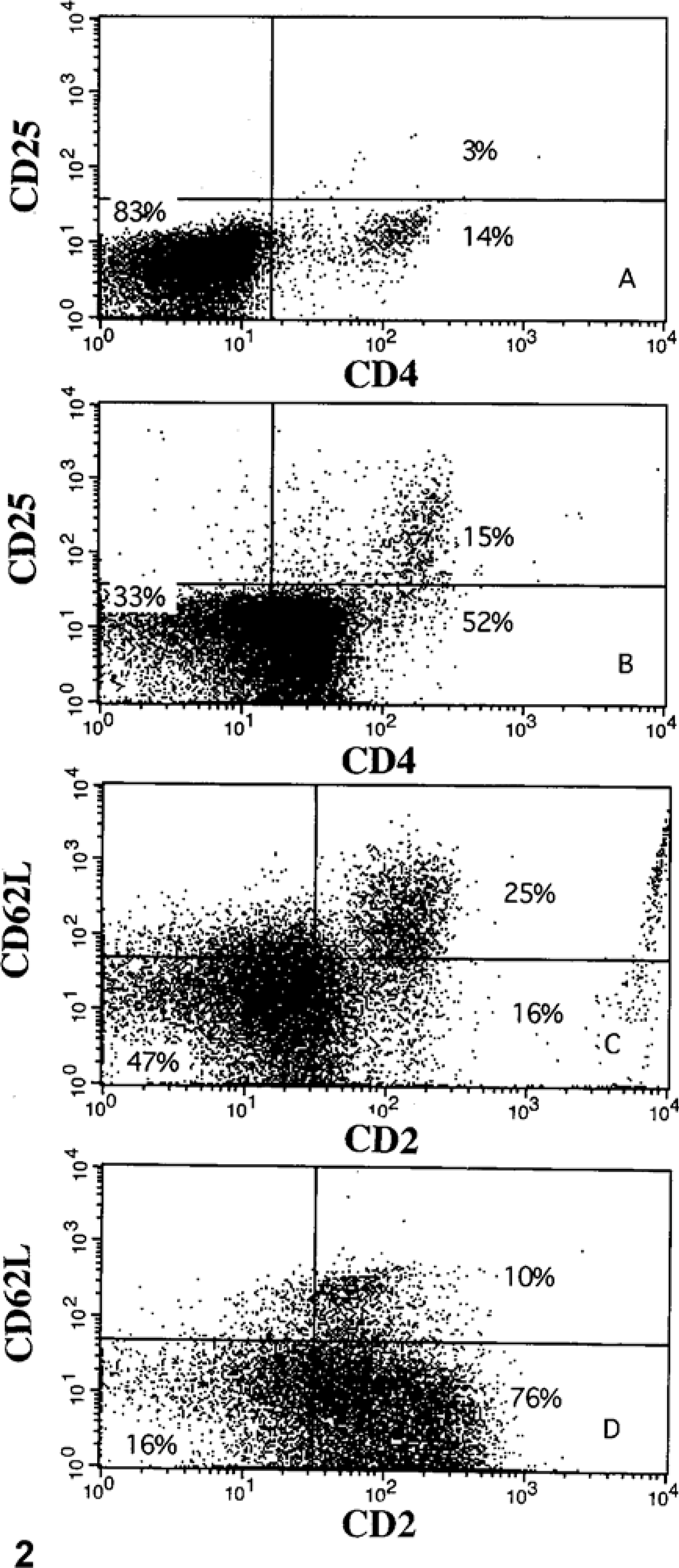
Flow-cytometric scatter plot of isolated ileal leukocytes from an uninfected cow and a cow with subclinical Johne's disease.
Immunophenotype of the spleen
Splenic leukocytes from uninfected cows contained a mixture of B cells and T cells (Table 4). The majority of cells were MHC class-II+ and many T cells were CD62L+ and activation marker positive. Only 6% cells were CD4+CD25+. When splenic leukocytes from infected cows were compared with those from uninfected cows, infected cows had a lower percentage of CD4+ T cells and a lower percentage of B cells.
Proliferative responses of ileal and splenic leukocytes
Lymphocyte proliferation in response to M. avium subsp. paratuberculosis sonicate, concanavalin A, and pokeweed mitogen was evaluated. Ileal lymphocytes from both uninfected and infected cows failed to proliferate in response to M. avium subsp. paratuberculosis sonicate (Fig. 3). Ileal lymphocytes from uninfected cows proliferated in response to both concanavalin A and pokeweed mitogen, however, ileal lymphocytes from infected cows proliferated poorly. The poor proliferative response did not appear to be due to cell death because Trypan blue staining indicated that greater than 75% of cells were viable after the 3 day incubation period. Splenic lymphocytes from uninfected cows failed to proliferate in response to M. avium subsp. paratuberculosis sonicate. Alternatively, splenic lymphocytes from infected cows proliferated in response to M. avium subsp. paratuberculosis sonicate. (P < 0.001; Fig. 4). Splenic lymphocytes from both infected and uninfected cows proliferated in response to concanavalin A and pokeweed mitogen.
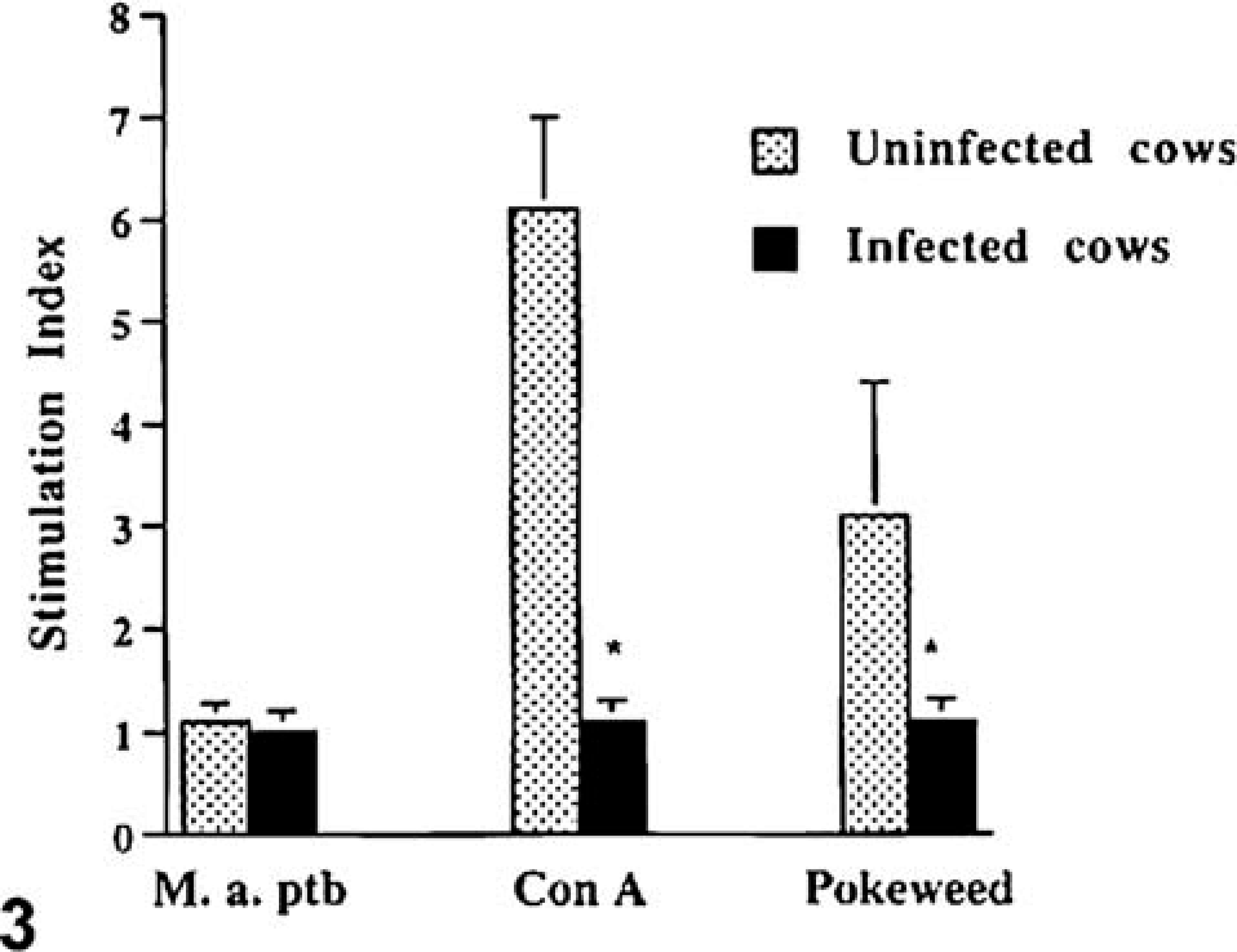
Proliferative responses of lymphocytes isolated from the ileum of cows subclinically infected with M. avium subsp. paratuberculosis and from uninfected cows. 5 × 105 lymphocytes were incubated with M. avium subsp. paratuberculosis sonicate (10 µg/ml), concanavalin A (10 µg/ml), or pokeweed mitogen (1 µg/ml), and cell proliferation was detected by binding of a green fluorescent dye to DNA. Asterisks indicate statistically significant differences between infected and uninfected cows (P < 0.05).
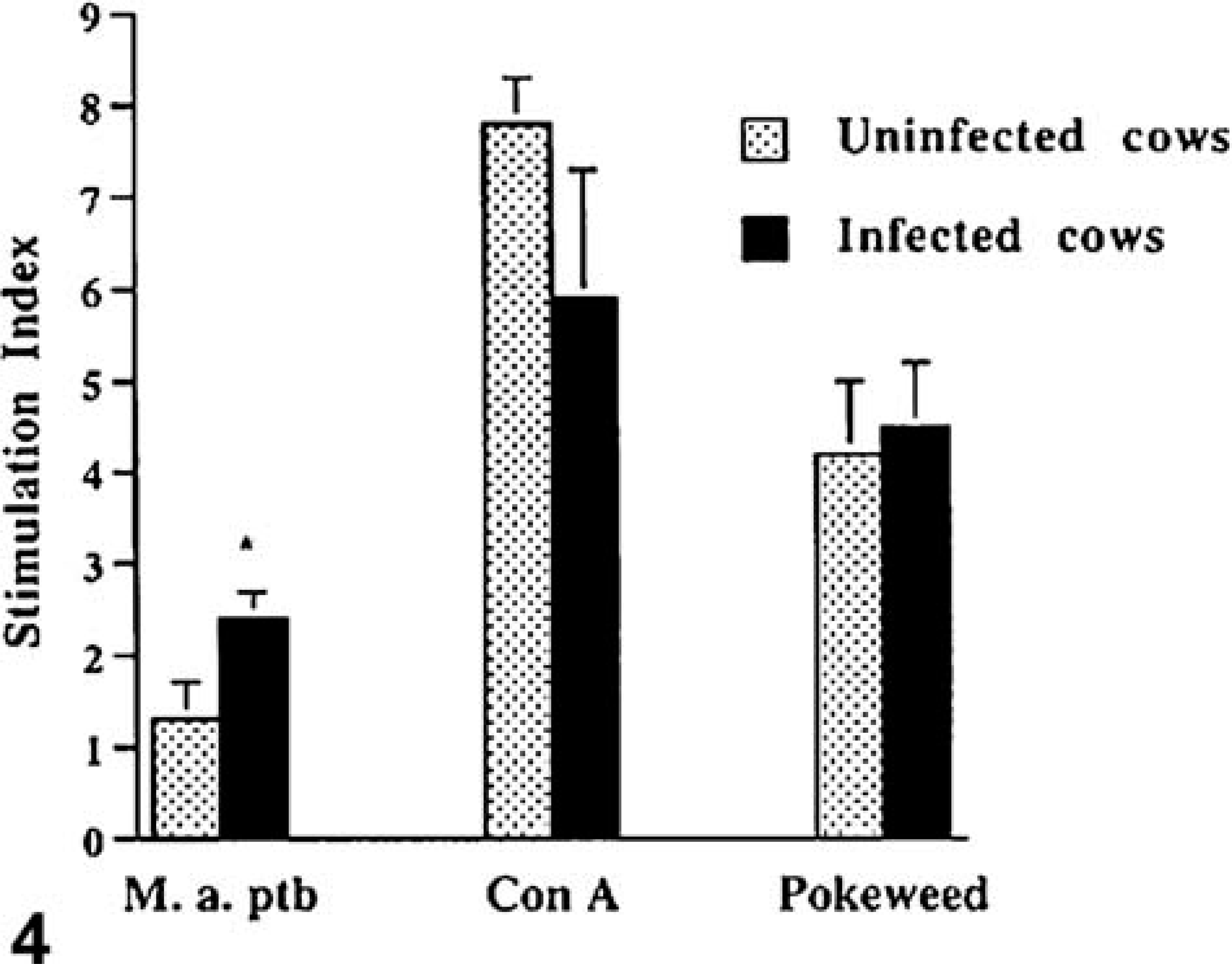
Proliferative responses of lymphocytes isolated from the spleen of cows subclinically infected with M. avium subsp. paratuberculosis and from uninfected cows. The 5 × 105 lymphocytes were incubated with M. avium subsp. paratuberculosis sonicate (10 µg/ml), concanavalin A (10 µg/ml), or pokeweed mitogen (1 µg/ml), and cell proliferation was detected by binding of a green fluorescent dye to DNA. Asterisk indicate statistically significant differences between infected and uninfected cows (P < 0.05).
Discussion
The results of the present study support the hypothesis that the mucosal immune response to M. avium subsp. paratuberculosis is attenuated in the subclinical stage of Johne's disease in cattle. When compared with control cows, the spectrum of immune cells was altered in Johne's disease. Although the total number of lymphocytes and the percentage of T cells were greater in infected cows, the percentage of memory cells (CD2+CD62L−) and regulatory T cells (CD4+CD25+) was greater. Further evidence of immune hyporesponsiveness includes a marked decrease in T cells with an activated phenotype and in cells expressing MHC class II. Previous studies have documented that in vitro infection of monocyte-derived macrophages with M. avium subsp. paratuberculosis resulted in down-regulation of MHC class-I and class-II expression. 35
Additional evidence of immune suppression is provided by the lymphocyte proliferation data. Unlike the spleen, ileal lymphocytes from infected cows failed to proliferate in response to M. avium subsp. paratuberculosis sonicate. Additionally, poor proliferative responses of ileal lymphocytes in response to concanavalin A suggest a generalized T cell hyporesponsiveness. Therefore, the immune response in the ileum in Johne's disease appears to be biased toward a hyporesponsive memory and regulatory T-cell phenotype.
The distribution of lymphoid cells in the intestinal mucosa of cattle in this study is similar to that described for mice. Several studies conclude that mouse mucosal lymphocytes are enriched for memory and regulatory T cells. 16, 23 Three types of regulatory T cells have been described in mice, including CD4+CD25+, Tr1, and Tr2 cells. 23, 30 Tr1 and Tr2 cells appear to exert their suppressive effects primarily through production of the interleukin (IL)-10 and transforming growth factor (TGF)-β. 23 CD4+CD25+ T cells suppress IL-2 production and promote cell cycle arrest of CD4+CD25− and CD8+ T cells. 23, 24 Additionally, CD4+CD25+ T cells induce CD4+CD25− T cells to differentiate into Tr1 and Tr2 cells. 23, 30
CD4+CD25+ T cells appear to be important to disease susceptibility. In mice infected with Listeria monocytogenes, depletion of CD4+CD25+ T cells early in the infection enlarged the population size of antigen-specific CD8+ T cells. 19 In other studies, long-term persistence of leishmaniasis and tuberculosis in mice was dependent on the presence of CD4+CD25+ T cells. 4, 15 In skin chronically infected with Leishmania major, 40 to 50% of CD4+ cells were CD25+ indicating a suppressor phenotype. 4 These cells produced large amounts of IL-10 and little gamma interferon (IFN-γ) and suppressed proliferation of CD4+CD25− T cells. 4
Several previous studies have addressed the mucosal immune response in ruminants infected with M. avium subsp. paratuberculosis. 1, 3, 6, 7, 22 The distribution of lymphocytes in the ileal Peyer's patches of lambs experimentally infected with M. avium subsp. paratuberculosis was investigated. 2, 3 As in the present study, the percentage of T cells was greater and the percentage of B cells was lower 8 weeks after infection. Unlike the present study, γδT cells were reported to increase. 2, 37 Proliferative responses of mucosal leukocytes from sheep naturally infected with M. avium subsp. paratuberculosis also were evaluated. Three of six sheep with the tuberculoid form of Johne's disease had proliferative responses of lamina propria–derived lymphocytes in response to Johnin pure protein derivative. 6 Alternatively, only 1 of 11 sheep with the lepromatous form of Johne's disease proliferated. Lymphocytes from sheep with the tuberculoid form of Johne's disease had greater production of IFN-γ and IL-2 in response to Johnin pure protein derivative than did lymphocytes from sheep with lepromatous lesions. 7 This response appeared to be more effective because sheep with tuberculoid-type lesions had few acid-fast organisms in intestinal lesions, whereas sheep with lepromatous lesions had large numbers of organisms.
Cytokine production and gene expression by ileal leukocytes from cows naturally infected with M. avium subsp. paratuberculosis have been evaluated. 1, 8, 17, 18, 33 In one study, DNA microarray identified 13 genes that had >1.5-fold difference in expression. 1 These genes included greater expression of interleukin-1α (IL-1α), tumor necrosis factor receptor-associated protein, N-cadherin, β1-integrin, and macrophage chemotatic protein-2 in infected cows. Ileal tissue from cows with clinical Johne's disease had greater expression of IL-1α, IL-1β, interleukin-6, IL-10, and TGF-β when compared with uninfected cows. 17, 22
In the present study, splenic lymphocytes from infected cows proliferated in response to M. avium subsp. paratuberculosis sonicate, whereas, ileal lymphocytes did not. This data suggest that the mucosal immune response is attenuated. These findings are consistent with those of previous studies. Cattle infected with M. avium subsp. paratuberculosis mount a systemic cell-mediated immune response (Th1-type) and peripheral blood lymphocytes from subclinically infected cows were reported to have substantially greater proliferative responses to both concanavalin A and M. avium subsp. paratuberculosis sonicate than did uninfected cows. 29, 31 The cell-mediated immune response appears to wane near the onset of clinical signs of disease at which time the organism may disseminate beyond the intestine. 29 Taken together, these observations have lead to the hypothesis that the systemic, cell-mediated, immune response prevents M. avium subsp. paratuberculosis from spreading beyond the intestine during the subclinical stages of disease. 29
Failure to induce an effective cellular immune response in the mucosa in M. avium subsp. paratuberculosis-infected cattle may be due to the altered antigen-presenting and costimulatory functions of macrophages. The present as well as previous studies have documented a decrease in MHC class I and class II expression on the surface of bovine macrophages and lymphocytes associated with M. avium subsp. paratuberculosis infection. 35 Bovine macrophages also have reduced MHC class II mRNA expression when infected with M. avium subsp. paratuberculosis. 34 Greater expression of the immunoinhibitory cytokine IL-10 and lower expression of the immunostimulatory cytokine IL-12 may also contribute to failure to develop an effective immune response. 36
In conclusion, results of this study indicate that the immune response to M. avium subsp. paratuberculosis in the intestinal lamina propria may be attenuated during the subclinical stages of Johne's disease. This effect may be induced, at least in part, by the proliferation of regulatory T cells that nonspecifically suppress mucosal immune responsiveness.
Footnotes
Acknowledgements
This work was supported by a grant from the University of Minnesota, Academic Health Center.