
Abstract
Mammalian cells contain two related but unique isoforms of cyclooxygenase (COX-1 and COX-2). COX-1 is expressed constitutively in a majority of tissues and is involved in the production of prostaglandins (PGs) that modulate normal physiologic functions. COX-2 is inducible by various stimuli and is involved in the production of PGs that modulate physiologic events in development, cell growth, and inflammation. With the exception of peribronchial glands and chondrocytes of peribronchial cartilage, COX-2 is not detectable in the normal lung of nonhuman primates. We evaluated COX-2 expression by immunohistochemical methods in the inflammatory lesions of two cynomolgus monkeys (Macaca fascicularis) with acute severe pneumonia. Both monkeys exhibited acute severe bronchopneumonia; histologically, lung lesions were characterized by infiltration of large numbers of neutrophils and fewer macrophages, mild bronchial epithelial hyperplasia, and slight type-2 pneumocyte hyperplasia. In both monkeys, mild to marked COX-2 immunoreactivity was detected within the cytoplasm of macrophages, bronchial epithelial cells, type-2 pneumocytes, and endothelial cells of blood vessels. No COX-2 immunoreactivity was detectable in the neutrophils that constituted >90% of the inflammatory cells. These observations suggest that in acute inflammatory lung lesions in nonhuman primates 1) COX-2 is induced in the bronchial and alveolar epithelial cells, 2) macrophages are the primary inflammatory cells that exhibit COX-2, and 3) neutrophils do not express COX-2.
Prostaglandins (PGs) play an integral part in the physiologic and pathophysiologic processes in the lung, such as perfusion-ventilation, surfactant homeostasis, macrophage-related inflammation, bronchial mucus secretion and transport, and regulation of bronchial tone. 2 8 15 Two isoforms of cyclooxygenase (COX-1 and COX-2) can catalyze the metabolism of arachidonic acid to PGs. These isoforms are expressed differentially; COX-1 is constitutively present in a majority of tissues, whereas COX-2, absent under basal conditions, is induced during development, cell growth, inflammation, and stress. 5 9 11 13
Information regarding function and in situ distribution of these isoforms in the respiratory tract is not extensive. In a study of the differential expression of COX-1 and COX-2 in healthy ovine lungs, the presence of COX-1 protein but the complete absence of the COX-2 protein was demonstrated in the developing ovine lung. 4 Because COX-1 was prevalent throughout ovine lung development, this isoform was considered responsible for PG-mediated vasodilation, bronchodilation, and surfactant synthesis under normal circumstances. Prostanoids are present in high concentrations under various respiratory disease conditions, such as sepsis and adult respiratory distress syndrome (ARDS). 7 16 The PGs observed in stressed lungs could be generated by either COX isoform originating from resident lung parenchymal cells or from one or more types of infiltrating inflammatory cells. Macrophages are postulated to be the primary cell type expressing COX-2 and producing PGs at inflammatory sites. 13 Although neutrophils are the most common cell type found in acute inflammatory conditions, information about COX expression and PG production by this cell type remains controversial. Here, we present data on the distribution of both COX isoforms in normal primate lungs and in primate lungs with acute neutrophilic inflammatory lesions.
Normal lungs were obtained from 3 adult cynomolgus monkeys (Macaca fascicularis, laboratory accession number OL8075). Lungs with inflammatory lesions were obtained from two adult monkeys with acute pneumonia (OL8078 and OL8102). All monkeys were maintained at the Searle Research and Development facility (Skokie, IL) and were part of investigational studies. The animals were housed in individual stainless steel primate cages and were cared for according to the Animal Welfare Act (CFR, Title 9) and the NIH Guidelines for the Care and Use of Laboratory Animals. All experimental procedures received prior approval from the Searle Institutional Animal Care and Use Committee.
Standard immunohistochemical procedures and commercially available reagents were used for the immunolocalization of COX-1 and COX-2 as previously described, with minor modifications. 9 Formalin-fixed tissues were embedded in paraffin (Paraplast X-tra, Oxford Labware, St. Louis, MO), cut in 4–5-mm sections, and mounted on positively charged slides (Superfrost Plus, Erie Scientific Co., Portsmouth, NH). Sections were then dewaxed, rehydrated in xylenes and descending alcohols, and blocked for endogenous avidin/biotin. All tissues were permeabilized in 0.3% triton/0.2% saponin/1% bovine serum albumin (BSA, Sigma, St. Louis, MO) in phosphate-buffered saline (PBS), preblocked in PBS containing 10% normal goat serum (Vector Laboratories, Burlingame, CA), and incubated in primary antibody overnight at 4 C. The COX-1 and COX-2 primary antibodies were PG H synthase (PGHS)-1 polyclonal rabbit anti-human (160108, Cayman Chemical Co., Ann Arbor, MI) and PGHS-2 polyclonal rabbit anti-human (PG27, Oxford Biomedical Research, Oxford, MI) antibodies, respectively. These antibodies were diluted 1:100 in 1% BSA-PBS. Immunoreactive complexes were detected via an enhanced streptavidin–biotin affinity system (Omnitags Plus, Shandon-Lipshaw, Pittsburgh, PA) and visualized with diaminobenzidine (Dako, Carpinteria, CA), which reacts with peroxidase to give a brown reaction product. Slides were counterstained briefly in hematoxylin-1 (Richard-Allan Scientific, Kalamazoo, MI). Control slides were not treated with primary antibody but were incubated with either biotinylated goat anti-rabbit IgG (Vector) or biotinylated horse anti-rabbit IgG (Vector) at the appropriate dilution to accompany the positive slides.
Both monkeys with inflammatory lung lesions (Nos. OL8078 and OL8102) had a clinical history of fever (104 F) and labored breathing and euthanatized. At necropsy, lungs in both animals were diffusely firm, dark brown, and edematous. Histologically, both exhibited severe bronchopneumonia characterized by infiltration of large numbers of neutrophils (>90% of inflammatory cells) and fewer macrophages, mild bronchial epithelial hyperplasia, and slight type-2 pneumocyte hyperplasia (Figs. 1, 2). In monkey No. OL8078, there were large numbers of gram-positive cocci and inflammatory cells within airways and alveoli consistent with acute bacterial pneumonia. In the lungs of monkey No. OL8102, there was a large amount of foreign material (plant material) within bronchioles and alveoli, consistent with aspiration pneumonia. There were no remarkable morphologic changes in the lungs of normal monkeys at gross and histologic examinations.
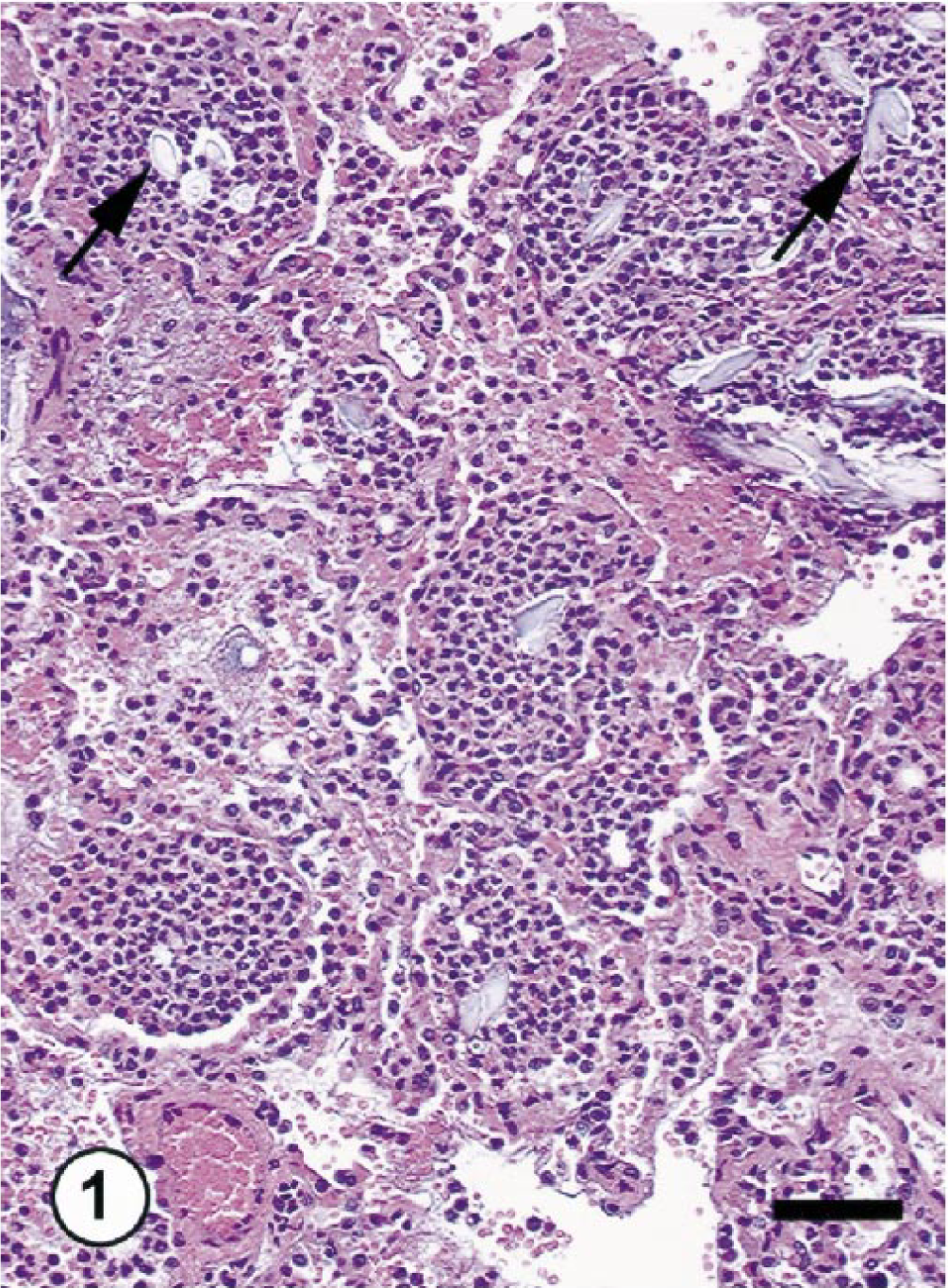
Lungs; cynomolgus monkey OL8102. Severe aspiration pneumonia. Note marked neutrophilic infiltrates in the lung parenchyma and foreign material (arrows). HE. Bar = 25 μm.
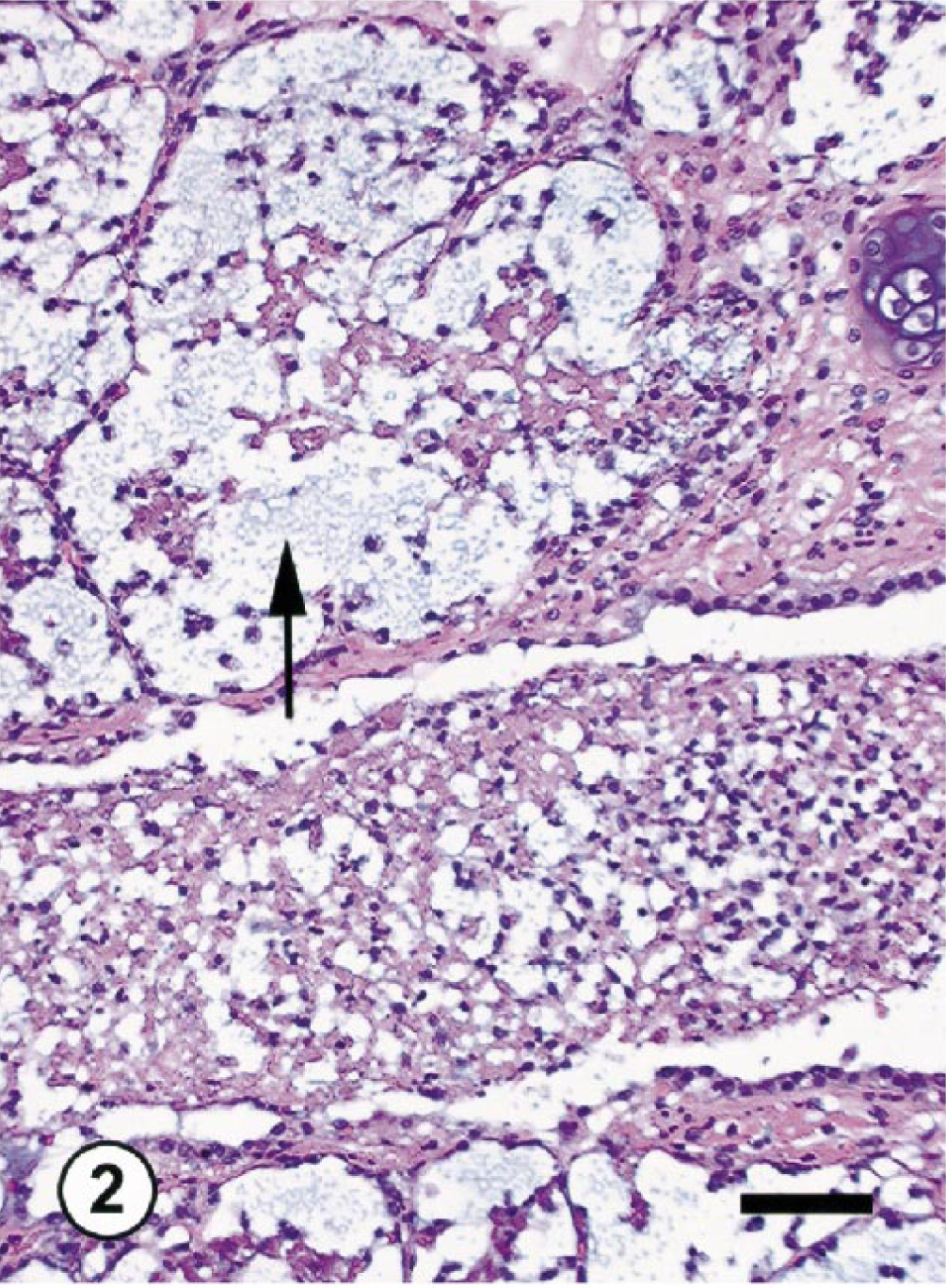
Lungs; cynomolgus monkey OL8078. Acute bacterial pneumonia. Note marked neutrophilic infiltrates in the lung parenchyma and bacteria (arrow). HE. Bar = 25 μm.
The immunohistochemical evaluations of lungs from the three normal monkeys revealed COX-1 expression in the vascular endothelium and alveolar epithelial cells throughout the lung and COX-2 expression in peribronchial glands and chondrocytes of peribronchial cartilage (Fig. 3). In the two monkeys with inflammatory lung lesions, there was no change in COX-1 expression; however, mild to marked COX-2 immunoreactivity was detected within the cytoplasm of macrophages, bronchial epithelial cells, type-2 pneumocytes, and endothelial cells of blood vessels (Fig. 4). With the exception of rare cells (<1%), no COX-2 immunoreactivity was detectable in the neutrophils that constituted >90% of the inflammatory cells. The COX-2 immunoreactivity in those few neutrophils was slight and punctate and was considered secondary to engulfment of degenerated and necrotic COX-2–positive cells. The intensity of COX-2 staining in peribronchial glands and chondrocytes of peribronchial cartilage did not change in these two monkeys in comparison with normal monkeys.
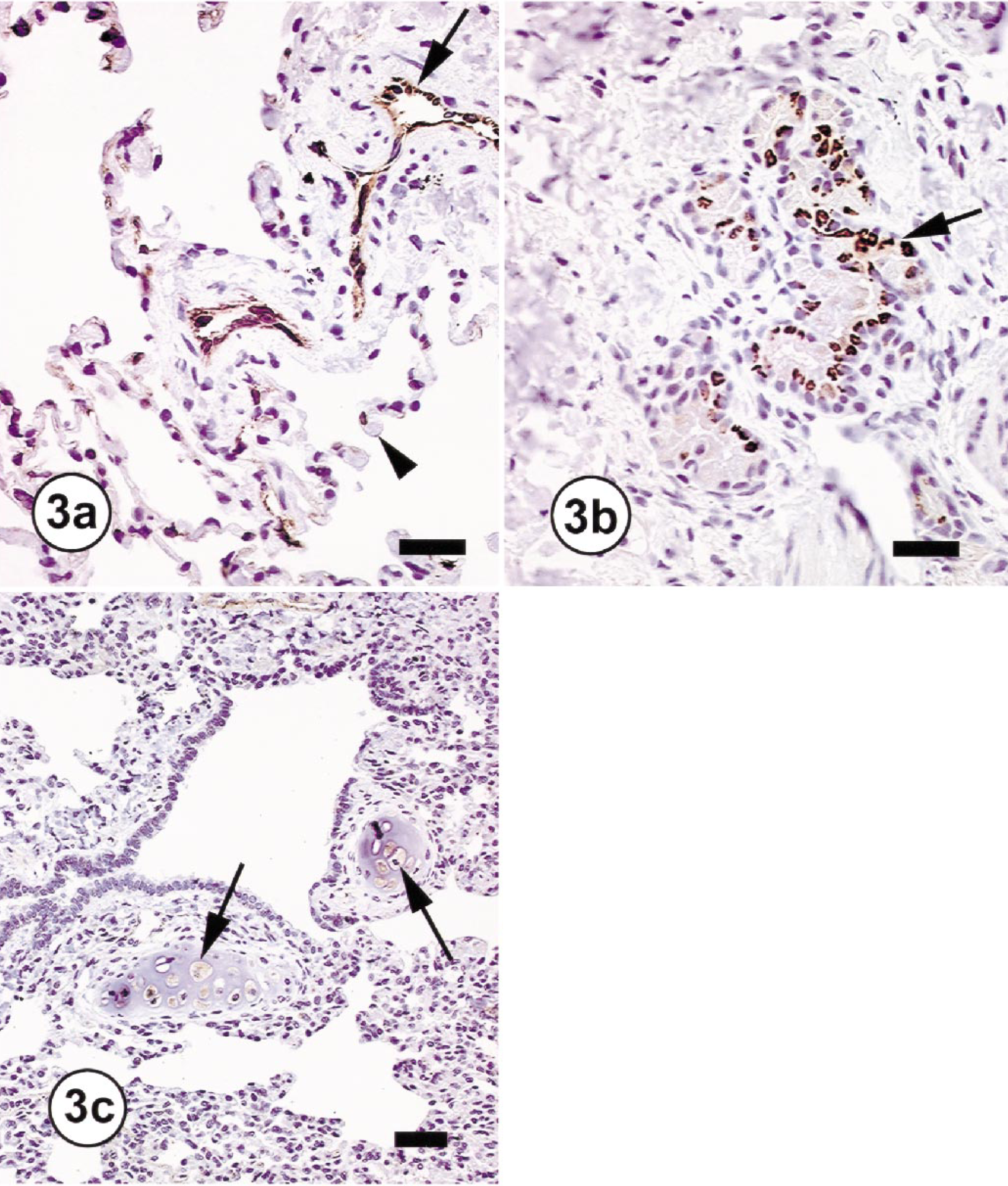
Lungs; cynomolgus monkeys (OL8075). Immunohistochemical staining for COX-1 and COX-2 in the normal lung.
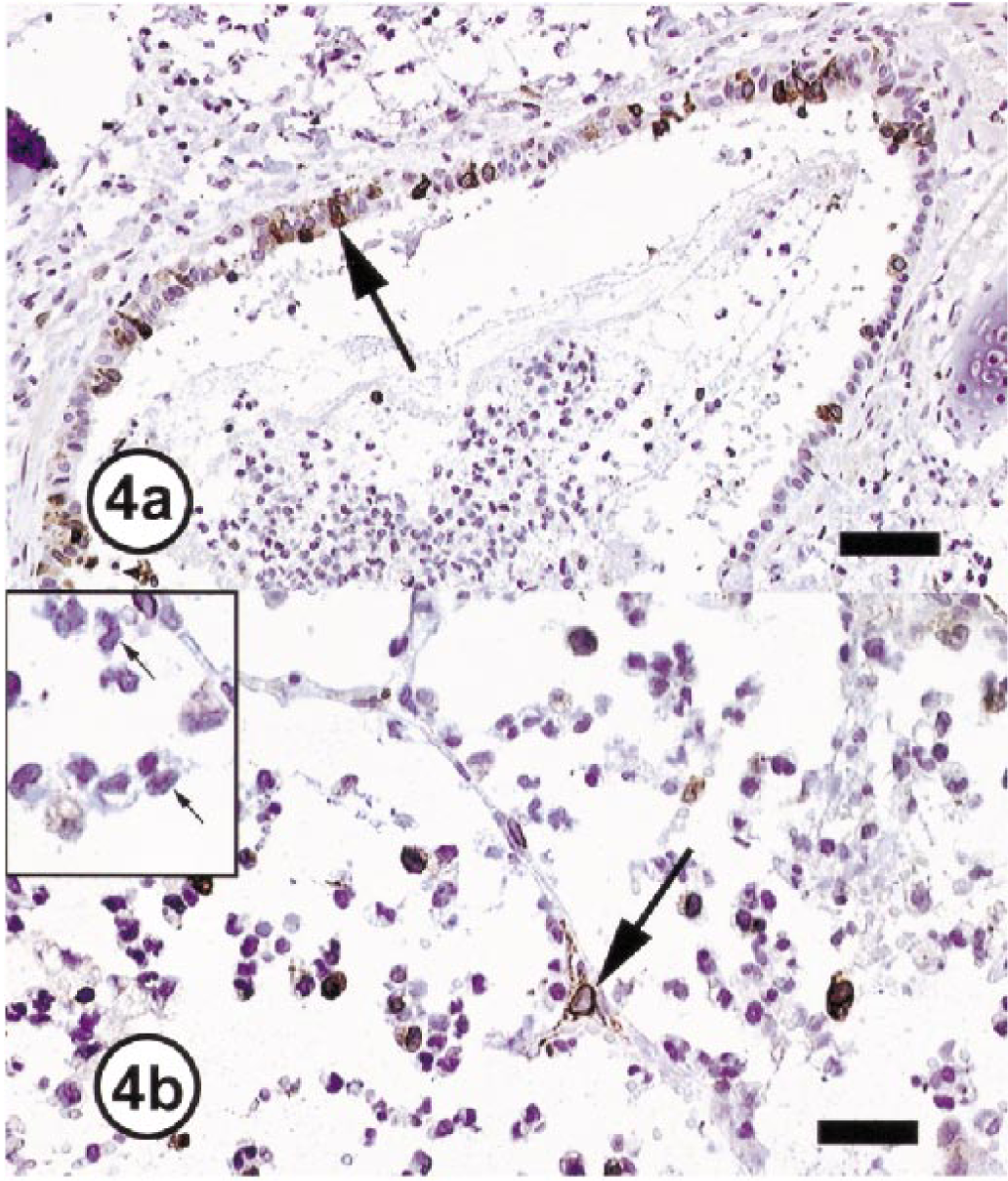
Lungs; cynomolgus monkeys. Immunohistochemical staining for COX-2 in inflammatory lung lesions.
Prostanoids generated by COX play an important role in normal pulmonary functions. Immunohistochemical evaluations demonstrate the constitutive but differential expression of both COX-1 and COX-2 in primates. The results obtained with these monkeys are similar to previous observations of the distribution of COX in the lungs of rats and sheep but with several differences. 4 6 In the ovine lung, COX-1 is the main isoform involved in PG-mediated vasodilation, bronchodilation, and surfactant synthesis at various stages of lung development, whereas COX-2 protein is not detectable. 4 In rats, both COX-1 and COX-2 are present at various sites in the lung and are implicated in regulatory processes under physiologic conditions. 6
In the monkeys with inflammatory lung lesions, the extent of COX-1 expression did not change, which suggests a lack of involvement of this isoform in acute inflammatory conditions. However, immunoreactive COX-2 was markedly induced in bronchiolar and alveolar epithelial cells and blood vessels. These results suggest that COX-2 is induced in lung parenchyma during inflammation, a conclusion supported by recent observations of COX-2 induction by inflammatory cytokines in human cultured airway epithelial cells and smooth muscle cells. 3 14 COX-2 induction in alveolar epithelial and interstitial cells has also been reported in a mouse model of oxygen-induced ARDS. 1
Of the inflammatory cells in the affected monkey lungs, macrophages but not neutrophils exhibited strong immunoreactivity to COX-2, suggesting that neutrophils may not take part in the production of inflammatory prostaglandins. These results are supported by observations of increased COX-2 expression in alveolar macrophages in the mouse model of oxygen-induced ARDS. 1 The lack of COX-2 in neutrophils, however, is in contrast to recently described COX-2 expression in isolated neutrophils following stimulation with lipopolysaccharide (LPS) or with inflammatory cytokines (interleukin [IL]-1, IL-8, and tumor necrosis factor α) in the isolated neutrophils. 10 12 However, conflicting reports warrant further study of the neutrophilic expression of COX-2. For example, constitutive expression of COX-2 in freshly isolated unstimulated neutrophils and inhibition of LPS-induced COX-2 expression in neutrophils with IL-4 and IL-10 were observed in one study 12 but not in another. 10 The discrepancies between these reports and our observations could be due to several variables, including differences in in vitro assay conditions, the in vitro versus in vivo microenvironment, the specificity of COX-2 antibodies used in immunohistochemical and western blot assays, and the effective blocking of endogenous peroxidase, which may result in false-positive staining reactions. The source of COX-2 antibodies used in all three investigations was different. The COX-2 antibodies used in the present study were obtained from a commercial source (Oxford); these antibodies have been validated previously for use in immunohistochemical studies for the detection of human and monkey COX-2. 9
COX-2 immunoreactivity in neutrophils in sections of lungs from individuals with ARDS and sepsis has been described; 10 however, the staining intensity and the number of neutrophils stained in a given field were variable, suggesting that this observation was possibly due to poor sensitivity of the immunohistochemical procedure, variation in the length of time an individual neutrophil was exposed to the agonist, or the differential expression of positive or negative signaling molecules that may modify COX-2 expression. The reason only a few neutrophils would exhibit COX-2 immunoreactivity might be engulfment of COX-2 protein or cell constituents from the neighboring dead and dying COX-2 positive macrophages and epithelial cells. In addition to the monkeys described here, we have evaluated over 20 cases of neutrophilic inflammation in various tissues from dogs, mice, and rats and were unable to unequivocally demonstrate COX-2 expression in neutrophils in any instances (unpublished data).
The available information does not provide definitive support for the expression of COX-2 in neutrophils. COX-2 may be transiently expressed in neutrophils under the regulatory influence of certain agonists. 10 12 Additional studies are needed to confirm and fully understand the role of COX-2 neutrophilic inflammatory conditions. These immunohistochemical evaluations of inflammatory lung lesions in cynomolgus monkeys demonstrate that 1) COX-2 is induced in bronchial and alveolar epithelial cells and 2) macrophages and not neutrophils are the primary inflammatory cells expressing COX-2.