
Abstract
The 129 mice are well recognized for their low fertility and it is speculated that this lack of fertility may be due to the oocyte condition. In this study we investigated superovulation regimens for the 129S1/SvImJ mouse strain to improve the oocyte quality and fertility rate of in vitro fertilization (IVF). Female mice were divided into four groups based on hormone and timing of injection. Group 1 received pregnant mare serum gonadotropin (PMSG) and 48 h later human chorionic gonadotropin (hCG); using the same dose, group 2 received hCG 52 h post-PMSG and group 3, 55 h post-PMSG. Group 4 received buserelin (gonadotropin-releasing hormone agonist [GnRH]) followed 24 h later by PMSG and then hCG 55 h post-PMSG. IVF was performed using 129S1/SvImJ oocytes and sperm; C57BL/6J sperm with 129S1/SvImJ oocytes was used as fertility control. The IVF fertility rate was 1% (Groups 1 and 2), 17% (Group 3) and 55% (Group 4) for 129 oocytes fertilized with 129 sperm. For 129 oocytes fertilized with C57BL/6J sperm, the fertility rate was 5% (Group 1), 10% (Group 2), 40% (Group 3) and 59% (Group 4). These results suggest that extending the interval time between PMSG and hCG and giving GnRH in addition to the standard PMSG and hCG treatments can improve IVF fertility rate of 129S1/SvImJ mouse strains significantly.
Assisted reproductive technology (ART) – which is a conjunction of techniques that includes embryo and gamete manipulation – is a powerful and essential tool for facilitating maintenance of laboratory animals for biomedical research and for reproductive applications in humans and domestic animals. Today, researchers studying reproductive processes, laboratories creating genetically-modified animals and repositories archiving cryopreserved embryos routinely use and often combine several ARTs to meet their needs. In mice, in vitro fertilization (IVF) is one of the most important techniques among ARTs. Because of the enormous number of mutant, transgenic and knockout mice archived by sperm and embryo cryopreservation, IVF has become the most popular technique for producing sufficient numbers of embryos for colony expansion, re-derivation and cryopreservation. Although successful IVF is important to subsequent embryo manipulation such as cryopreservation or re-derivation by embryo transfer, knowing the optimal IVF conditions becomes essential.
It is well established that in the mouse the genetic background defines the reproductive performance of strain used and influences the success of IVF. 1–3 Reproductive traits such as litter size, sperm production and hormone-induced ovulation rate differ between mouse strains. 4 Furthermore, inbred mouse strains have defined reproductive parameters so distinctive as to be considered a characteristic of the strain. 2,5 Inbred strains also differ in their capacity for embryo development in culture 6 and in survival after embryo cryopreservation. 7 The 129 mouse strain family has many sublines; 8 among them, the 129S1/SvImJ strain shows high incidence of spontaneously testicular teratomas 9,10 and is popular for being the first line from which derivation of embryonic stem (ES) cells was established. 11,12 Since then, many ES cell lines with high frequency of germline transmission have been established using 129 mice. Consequently, a large number of gene-targeted mice have been created using the 129-derived ES cells, making this mouse strain one of the most popular strains in genetic research; however, 129 mice are also known for their poor reproductive performance compared with other popular inbred strain such as C57BL/6, having smaller litter size 13,14 and low IVF rate. 2,3,15,16 It was reported that the IVF using 129 oocytes and sperm (inbred cross) is lower than using sperm or eggs fertilized with other strains. 3 Hino et al. 15 found that the fertility rate was less than 50% even when 129 females were mated with proven fertile C57BL/6J males; however, when C57BL/6J females were mated with 129 males, all females demonstrated 100% fertility. They concluded that low fertility occurred whenever 129 females were used. These findings suggest that the low fertilization rate of 129 mice may be due to an oocyte problem. In our hands, although the 129 females, especially the 129S1/SvImJ strain respond very well to superovulation treatment, the production of large numbers of oocytes does not correlate with acceptable quality and fertility rate (data not shown).
Since Fowler and Edwards in 1957, exogenous gonadotropins are widely used for mouse superovulation treatment. In most regimens for humans, chorionic gonadotropin (eCG) is given to preempt the endogenous luteinizing hormone (LH) surge and to induce the final stages of oocyte maturation that precede ovulation. 17 Unlike the normal spontaneous ovarian cycle in which ovarian–hypothalamic–pituitary feedback systems ensure that the LH mid-cycle surge is released only when the preovulatory follicles are sufficiently mature in mice, the human chorionic gonadotropin (hCG) is given following a standard regimen of 46–48 h post-eCG (pregnant mare serum gonadotropin [PMSG]) irrespective of strain. Hillier et al. 18 found that prolonging the time between PMSG and hCG improved the IVF fertility rate in mice. Clinically, the gonadotropin-releasing hormone (GnRH) is commonly used as an adjunctive therapy during ovarian stimulation. In mice, Kanter et al. 19 found that an intramuscular injection of GnRH 24 h before superovulation treatment with PMSG–hCG increased oocyte number and maturation. The primary objective of this study was to investigate different parameters of superovulation regimen in a 129 mouse strain to improve the quality of oocytes and fertility rate of IVF.
Materials and methods
Animals
Inbred wild-type mice 129S1/SvImJ and C57BL/6J were purchased from Jackson Laboratories (Bar Harbor, ME, USA). The mice were maintained in facilities accredited by the Association for the Assessment and Accreditation of Laboratory Animal Care International (AAALAC). Animal housing included a controlled light and dark cycle (14–10 h, lights on at 06:00 h and off at 20:00 h), ad libitum food, Purina 5058 irradiated high-fat rodent diet, ultra-filtered water, ventilated caging systems and standardized environmental enrichment system. Male mice were housed individually for at least five days prior to sperm collection. All experiments were conducted under the approval of the National Institute of Allergy and Infectious Diseases (National Institutes of Health, Bethesda, Maryland, USA) Institutional Animal Care and User Committee.
In vitro fertilization
IVF was performed using the method described by Vasudevan et al. 1 Research Vitro Fert (K-RVFE-50; Cook Medical, Inc, Bloomington, IN, USA) was used for sperm incubation, IVF and culture of zygotes. Sperm were collected from the caudae epididymides of three- to five-month-old male mice as described by Sztein et al. 3 and capacitated for 60 min at 37°C under 5% CO2 in the incubator. Sperm motility and concentration were confirmed by visual observation at the edge of the drop under a stereo dissecting microscope. Three to five two-month-old 129S1/SvImJ female mice were assigned to four groups: superovulated (group 1), superovulated with different timings of hCG (groups 2 and 3) and superovulated with GnRH pretreatment (group 4). Oocytes from 129S1/SvImJ females were fertilized by fresh sperm from 129S1/SvImJ and C57BL/6J males. Female mice were superovulated with 0.1 cm3 (5 IU in saline) of PMSG (Sigma, St Louis, MO, USA) intraperitoneally between 11:00 h and 15:00 h. At 48, 52 and 55 h post-PMSG, the mice were treated with 0.1 cm3 (5 IU) of hCG (Sigma) intraperitoneally between 15:00 h and 18:00 h Donor females were housed in the animal facility for 7–10 days before starting the hormone treatment. For the GnRH pretreated group, females were treated with 0.1 cm3 of GnRH (20 mg/kg buserelin, intraperitoneally) 24 h before superovulation ampullae and subsequently used for IVF. After sperm incubation (45 min), 10 μL of the fresh sperm suspension was added to a 250 μL drop of IVF medium. Thirteen to 14 h post-hCG treatment the females were euthanized and the isolated oocyte complexes were transferred to the sperm drop and incubated at 37°C for 5 h. The eggs were then washed and cultured overnight in IVF medium under the same conditions. The following day, the two-cell embryos were scored to evaluate the fertility and eventually prepared for either transfer to recipient females or for cryopreservation. Oocytes from the superovulated and GnRH treated superovulated females were pooled for each IVF; the experiment was done on three separate days.
Embryo transfer
To assess whether embryos produced by IVF using buserelin-treated females support development to term, 14 embryos from buserelin treated and control group were transferred to pseudopregnant mothers. Female CD1 mice (Charles River Laboratories International, Inc, Wilmington, MA, USA), 9–13 weeks of age were mated with vasectomized CD1 males (Harlan Laboratories, Inc, Frederick, MD, USA) to induce pseudopregnancy. The following morning, females with vaginal plugs were used as embryo transfer recipients. Embryos (14) were surgically transferred into the left oviduct of each female, as described by Nagy et al., 20 and the number of pups born was counted.
Statistical analysis
Fertility was considered as the percentage of two-cell stage embryos scored 24 h after insemination. For statistical analysis, percentages were transformed into arcsine values and then evaluated by a paired Student's t-test (one tailed). Differences were considered significant when a P value of <0.05 was obtained. Error bars on figures are SEM. Data were analysed using the GraphPad Prism version 5.0 computer program (GraphPad Software, San Diego, CA, USA).
Results
Results of the IVF procedures are summarized in the following tables and figure. Fertility was scored at the two-cell stage and calculated by dividing the number of two-cell embryos by the total number of oocytes exposed in the IVF dish. The fertility rate was high for group 3 (hCG given 55 h post-PMSG) when compared with groups 1 and 2 as shown in Table 1. The fertility rate was significantly higher for females treated with buserelin compared with females not treated with buserelin (shown in Table 2). The fertility rate for groups 1 and 2 versus group 3 was 1 ± 1% versus 17 ± 12% for 129 × 129 and 5 ± 4%, 10% versus 40 ± 22% for 129 × B6 (Table 1). The percentage of eggs fertilized for buserelin-treated and buserelin-untreated groups was 55 ± 16% versus 28 ± 10% for 129 × 129 cross and 59 ± 20% versus 32 ± 8% for 129 × B6 cross as shown in Table 2 and in Figure 1.
In vitro fertilization (IVF) of 129S1/SvImJ female mice treated with buserelin before superovulation. 129S1/SvImJ oocytes were fertilized by 129S1/SvImJ and C57BL/6 sperm
In vitro fertilization of mouse oocytes: effect of giving human chorionic gonadotropin (hCG) at different timings to induce ovulation Group 1: hCG was given 48 h post-PMSG; Group 2: hCG was given 52 h post-PMSG; Group 3: hCG was given 55 h post-PMSG *129 oocytes were fertilized by 129 sperm
†129 oocytes were fertilized by B6 sperm Effect of buserelin treatment on in vitro fertilization (IVF) The data shown in the table represent cumulative results of all IVF experiments done on three different days. Superscript letter represents significant difference in fertility rate between no buserelin and buserelin-treated groups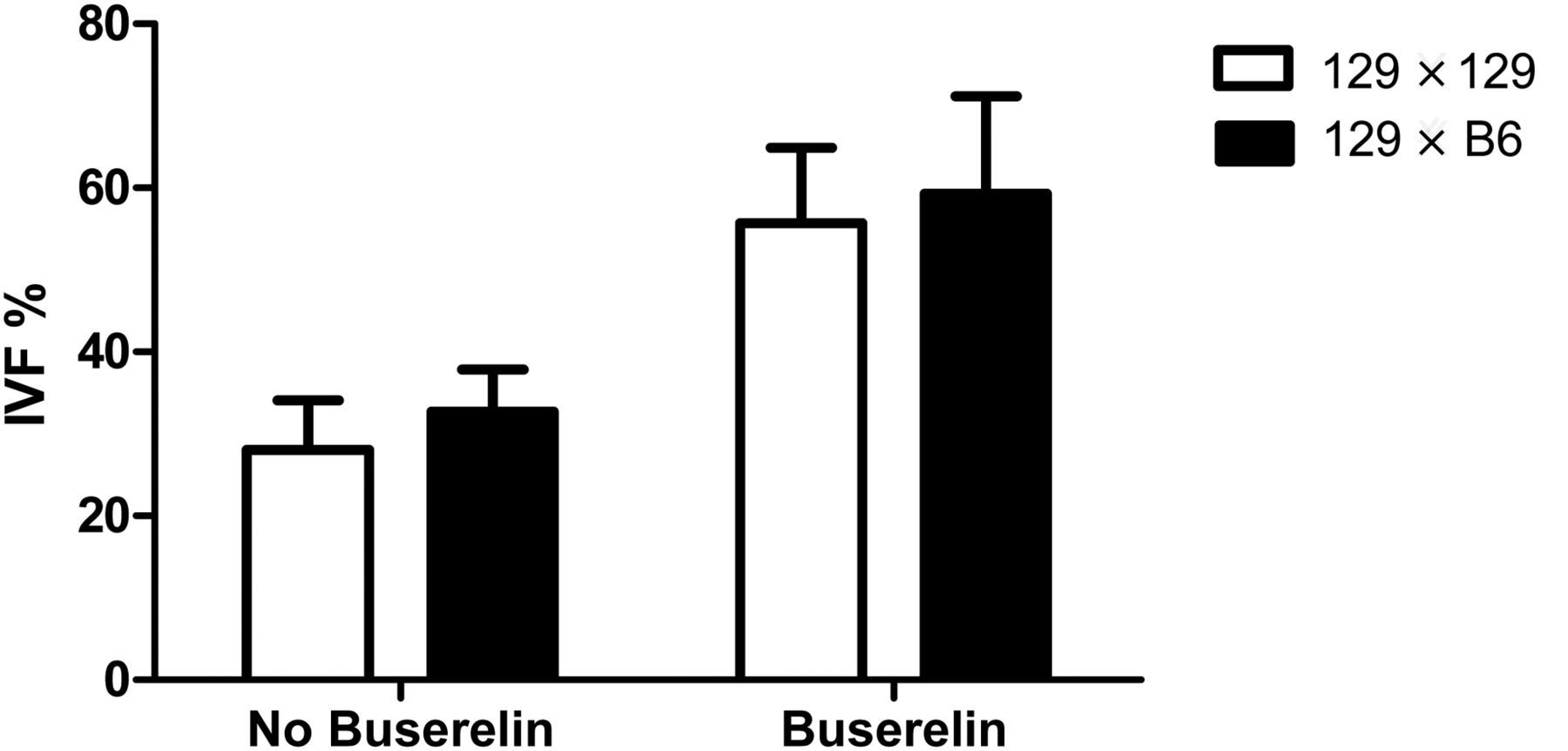
Newborn mice derived from buserelin-treated and buserelin-untreated groups
Since the rationale of the study was to improve the oocyte quality for IVF, it was important to assess that buserelin treatment does not affect the embryo's development to term. Fourteen embryos from each group were transferred to pseudopregnant CD1 females, and the number of pups born was counted. There was no marked difference between the two groups. The number of offspring born for the control group was 10 out of 14 and for buserelin-treated females was 18 out of 28.
Discussion
The results of this study shows that the IVF fertility rate of the 129S1/SvImJ strain can be improved significantly by extending the time of hCG injection after PMSG and by treating the females with GnRH before superovulation. The quality of the oocyte was clearly improved by the action of buserelin demonstrated by the increase in the fertility rate and as expected, the embryo development and birth rate were not altered by the buserelin.
Archiving mouse strains by embryo cryopreservation has great potential, because it preserves the whole genome and it is simple, rapid and inexpensive if we are able to generate embryos in vitro. The use of IVF to produce embryos for embryo banking and/or re-derivation can shorten the time required to complete the process; however, success in producing those embryos needed for cryopreservation is influenced by the genetic background and reproductive performance of the mouse strain used. 1,21 Although the 129 mouse strain has become popular because its embryos were used to derive many ES cell lines, all of them resulting in high-frequency germline transmission, they are also popular for their poor reproductive performance, which also affects IVF fertility rate. 15 Hino et al. found that the fertility rate was less than 50% even when 129 females were mated with C57BL/6J males with normal fertility. These findings suggest that the low fertilization rate of 129 mice may be due to an oocyte problem. In general, 129 females from all substrains respond well to standard superovulation treatment and large numbers of oocytes are obtained, but this occurrence does not correlate with acceptable quality and fertility of the oocytes produced. The rationale of this study was to improve the quality of oocytes of 129S1/SvImJ mice, which will be reflected in the IVF fertility rate. For that, we investigated different hormonal treatment parameters and combinations to optimize the quality of oocytes obtained from superovulated 129 mice.
The results of the study highlights one of the options for improving the IVF fertility rate of the 129 strain by extending the interval time between PMSG and hCG injection up to 55 h and by treating the females with 20 mg/kg of buserelin 24 h prior to the standard superovulation treatment. The increase in IVF fertility rate showed by the greater number of two-cell embryos obtained from the treated females may be due to the effect on oocyte maturation induced by the GnRH with its activity especially on the ovary. 22 Recent studies have shown that at least in mice, rats and rabbits – GnRH and its analogues exert a direct effect on gonadal function, affecting or influencing oocyte maturation both in vivo and in vitro. 22,23 It has been suggested that GnRH induces oocyte maturation via activation of specific GnRH receptors on granulosa cells 24,25 and that exposure to GnRH stimulates prostaglandin (PG) E2 and PGF2α synthesis in preovulatory follicles. 26 Also, the increasing concentration of PGs plays an important role in oocyte maturation. 27
The other possible reason for the increase in fertility rate could be the increase in the interval time between PMSG and hCG. It has been shown that moderate decreases in follicular maturation time before inducing ovulation with hCG caused consistent and marked reduction in oocyte fertility. 18 Hillier et al. in the same study showed that by increasing the interval time in superovulation treatment between the PMSG–hCG injections up to 50 h resulted in an increase in the oocyte fertility rate in vitro. A similar observation was made in hamsters, where the extension of the hCG time interval from 52 to 76 h resulted in an increased fertilization rate. 28
In conclusion, the supplement of a GnRH treatment 24 h before superovulation and extending the interval to 55 h between injections of PMSG and hCG result in an improvement of IVF fertility rate in the 129S1/SvImJ mouse strain, and as expected in this study, embryo development was not affected by the treatment, demonstrated by the normal number of pups born after embryo transfer. The results of this study show that, improving the fertility rate of this ‘difficult strain’ makes it possible to obtain a larger number of viable embryos for embryo banking or experimental studies.
Footnotes
ACKNOWLEDGEMENTS
This work was supported by the Intramural Research Program of the Division of Intramural Research, National Institute of Allergy and Infectious Diseases, National Institutes of Health. We thank Brenda Rae Marshall, DPSS, NIAID, for editing. Because the authors are government employees and this is a government work, the work is in the public domain in the USA. Notwithstanding any other agreements, the NIH reserves the right to provide the work to PubMedCentral for display and use by the public, and PubMedCentral may tag or modify the work consistent with its customary practices. Rights outside the US subject to a government use licence can be established.