
Abstract
Positron emission tomography (PET) provides a means of studying physiological and pharmacological processes as they occur in the living brain. Mice, rats, dogs, cats, pigs and non-human primates are often used in studies using PET. They are commonly anaesthetized with ketamine, propofol or isoflurane in order to prevent them from moving during the imaging procedure. The use of anaesthesia in PET studies suffers, however, from the drawback of possibly altering central neuromolecular mechanisms. As a result, PET findings obtained in anaesthetized animals may fail to correctly represent normal properties of the awake brain. Here, we review findings of PET studies carried out either in both awake and anaesthetized animals or in animals given at least two different anaesthetics. Such studies provide a means of estimating the extent to which anaesthesia affects the outcome of PET neuroimaging in animals. While no final conclusion can be drawn concerning the ‘best’ general anaesthetic for PET neuroimaging in laboratory animals, such studies provide findings that can enhance an understanding of neurobiological mechanisms in the living brain.
Keywords
A new era of neuroscience began with the invention of positron emission tomography (PET) for studying processes as they occur in the living brain.1–4 PET makes use of the radioactive decay of positron-emitting nuclides to derive an image of physiological and pharmacological events in a living organ such as the brain. PET is currently the primary procedure for studying molecular events in realtime in intact animals and humans. PET scanning can be used in all branches of pharmacology, molecular biology and medicine, including neuroscience, cancer research and cardiovascular biology.5–7
Three types of PET studies characterize brain research, namely blood flow, metabolism and neurotransmission.8,9 PET neuroimaging in animals is typically carried out to model aspects of the living human brain. In principle, any animal could be used for PET neuroimaging, but most studies are carried out in mice, rats, dogs, cats, pigs and non-human primates. 8 Some animals can be PET-scanned while awake, which avoids possible side-effects of anaesthesia in the central nervous system (CNS). They are sometimes trained for PET scanning in a head-restraining devise.10–20 The authors of such studies typically claim that awake animals are not stressed by the procedures. However, acute and chronic physical restraint are well-established stressors of laboratory animals.21–24 Most importantly, stress affects the welfare of animals and can be expected to affect the outcome of PET brain imaging.25,26 Patel et al. 26 demonstrated, for example, that restraint of awake rats markedly reduced binding potentials of the dopamine D2/3 receptor antagonist [11C]raclopride. PET neuroimaging in animals is, therefore, ordinarily carried out under general anaesthesia.
There is currently no gold standard for evaluating effects of anaesthetics on PET brain imaging in laboratory animals. In general, three approaches are in use. One approach involves comparing effects of two or more anaesthetics on PET findings in animals.27,28 This procedure can investigate differences between anaesthetic drugs, but it is unknown which drug condition most closely represents the awake state. A second approach requires that PET brain imaging is carried out in both awake and anaesthetized animals. 12 However, if awake animals are restrained for PET, then the findings may reflect a stressed condition. A third approach uses freely-moving animals that are injected with a PET radiotracer just prior to PET scanning under anaesthesia. 29 This procedure can provide information on awake brain function, but it typically requires many animals and does not supply dynamic PET images of underlying molecular events.
Generally speaking, anaesthesia can alter physiological parameters such as cerebral blood flow (CBF), which may then affect the kinetics of PET tracers. 30 Most anaesthetics decrease cerebral metabolism 31 and very often they also influence neurotransmission 32 (see also Table 1). Dynamic PET scanning usually requires 1–2 h per carbon-11-tracer, so anaesthesia for animal studies is very often applied for hours, including the time of preparatory surgical procedures. Because anaesthesia can inhibit an animal's homeostasis, which indirectly can influence PET results, 33 careful monitoring is important. 34 Hypothermia can develop, for example, even during a few minutes of anaesthesia, 35 and reduce brain metabolism with effects on pathological central processes. 36 Thus, body temperature and other physiological parameters, such as heart rate, blood pressure, respiration rate and oxygen saturation, should be monitored during anaesthesia. Because kinetics of several PET tracers are also affected by CBF, 37 parameters effecting CBF, such as carbon dioxide tension in arterial blood samples (as PaCO2) or in expired air (as ETCO2), should also be carefully monitored during anaesthesia. 38 In particular, we focus on the three anaesthetic agents that have been used most often for PET neuroimaging in laboratory animals, namely ketamine, propofol and isoflurane/halothane.
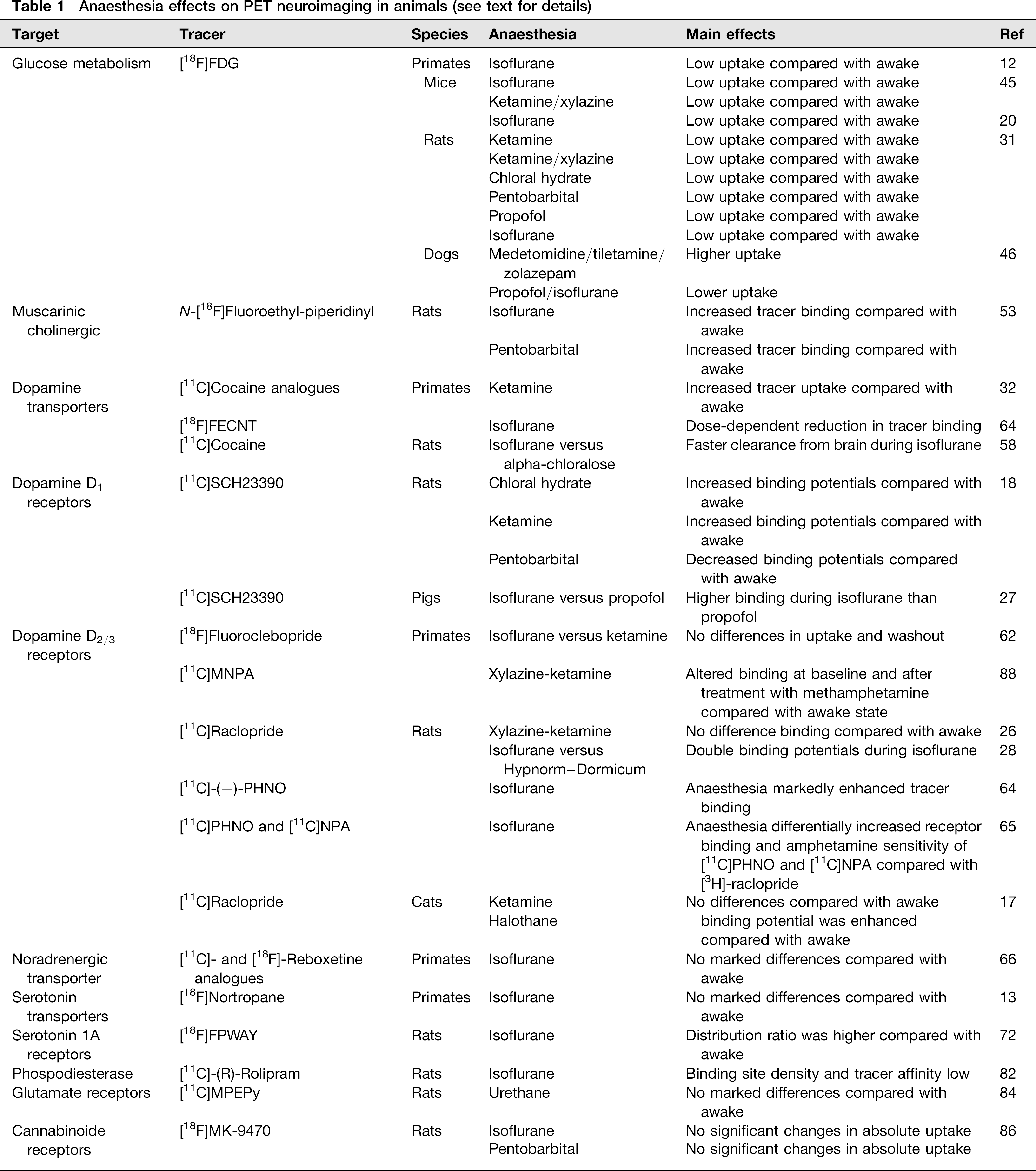
Anaesthesia effects on PET neuroimaging in animals (see text for details)
CBF and cerebral blood volume
Anaesthetic agents typically affect CBF, which can markedly affect the kinetics of PET radiotracers. 37 Ketamine is often used in PET studies carried out in laboratory animals, because it leaves respiration and cardiac output unaffected. 30 However, ketamine typically increases CBF and brain metabolism.39–42 Ketamine has indirect sympathomimetic effects, increases plasma noradrenaline concentration and can influence PET neuroimaging. 33 Propofol is a shortacting anaesthetic that rapidly crosses the blood–brain barrier; it has an onset of action of one minute. It is rapidly cleared from plasma, and therefore consciousness returns more rapidly with propofol than with most other anaesthetics. Propofol permits better cerebral autoregulation than most other anaesthetic agents. 43 However, propofol decreases CBF and may thereby also affect the kinetics of some PET tracers. 27 Isoflurane and halothane are used increasingly in PET neuroimaging, mainly because they allow good control of the length and depth of anaesthesia. 33 However, both compounds tend to increase CBF and to disrupt cerebral autoregulation in animals. 34
In summary, CBF and cerebral blood volume (CBV) are typically decreased by anaesthesia produced by propofol and increased by ketamine and isoflurane/halothane in animals used for PET neuroimaging.30,44 Hassoun et al. 17 noted, for example, that binding of [11C]raclopride in cat brain was enhanced, along with CBF, under halothane as well as ketamine anaesthesia. A similar enhancement of [11C]raclopride binding was noted in brains of rats anaesthetized with isoflurane. 28
Brain metabolism
Effects of anaesthesia on cerebral glucose metabolism have been studied by PET in several species using 2-deoxy-2-[18F]fluoro-
Cholinergic neurotransmission
General comments
Cholinergic neurotransmission takes place both in the brain and in the parasympathetic branch of the autonomic nervous system. Cholinergic neurotransmission is governed by acetylcholine, which acts on two types of receptors, known as muscarinic and nicotinic. Central muscarinic cholinergic receptors are involved in appetitive behaviours and cognition,48,49 whereas peripheral muscarinic cholinergic mechanisms affect cardiac, respiratory and digestive functions. 50 Questions remain, however, concerning whether anaesthetics have direct effects on peripheral muscarinic cholinergic neurotransmission or whether their actions are primarily the result of central pharmacological effects.51,52 Thus, anaesthetics have been found, in general, to lessen peripheral sympathetic neuroactivity, 50 but whether those effects are caused by direct local inhibition of muscarinic cholinergic neurotransmission or are the consequence of central baroreceptor mechanisms remains unclear.51,52
Muscarinic cholinergic receptors
Kilbourn et al. 53 studied the in vivo binding of N-[18F] fluoroethyl-piperidinyl benzilate to the muscarinic cholinergic receptor in rats. Compared with awake rats, anaesthesia with either isoflurane or pentobarbital markedly increased tracer binding in striatum, cortex and hippocampus. Pretreatment with the acetylcholinesterase inhibitor phenserine increased tracer binding in awake rats, but not in anaesthetized animals.
Monoaminergic neurotransmission
General comments
Central monoaminergic mechanisms have received much interest in neuropsychopharmacology.54,55 Dopamine, noradrenaline and serotonin are the major monoaminergic neurotransmitters. They are instrumental for behaviors such as motivated activities and neuropsychiatric disorders. For example, disturbances in dopaminergic neurotransmission are central to Parkinson's disease and psychosis,56,57 whereas noradrenergic and serotonergic neurotransmission are involved in mood disorders. 55 PET studies of monoaminergic pathways in laboratory animals are typically carried out in order to improve our understanding of molecular mechanisms in brain disease.
Dopamine transporters
Anaesthesia effects on dopamine transporters (DATs) have been studied in non-human primates and rats.32,58–60 Generally speaking, isofluorane has major, dose-dependent effects on DATs. Also ketamine may influence the results of PET studies on DATs in animals. Tsukada et al. 32 studied the effect of ketamine anaesthesia on the binding of two [11C]-labelled cocaine analogues to DATs in rhesus monkeys. The uptake of both radiotracers was higher during ketamine anaesthesia than in awake animals. Both tracers entered the brain in a dose-dependent manner, and kinetic analysis indicated that binding potentials were elevated by ketamine. Later, Votaw et al. 58 studied the effect of isoflurane on DATs in rhesus monkeys. They carried out PET scanning with the DAT tracer [18F]FECNT during isoflurane anaesthesia. The binding potential decreased when isoflurane was increased from 1% to 2%. These studies show that anaesthetics can markedly affect radioligand binding by DATs. Du et al. 59 investigated the effects of anaesthesia on clearance of [11C]cocaine in rat brains. They noted that [11C]cocaine clearance was faster from brains during isoflurane anaesthesia than during anaesthesia with alpha-chloralose. They also found that acute injection of cocaine (1 mg/kg) decreased CBF and CBV during isoflurane anaesthesia, but increased CBF and CBV during anaesthesia with alpha-chloralose. The mechanisms responsible for opposite effects of isoflurane and alpha-chloralose on CBF, CBV and drug clearance are unknown.
Dopamine D1 receptor
Anaesthesia effects on PET scanning of dopamine D1 receptors have been studied in rats and pigs.18,27 Momosaki et al. 18 evaluated the binding potential of [11C]SCH23390 during three different anaesthesia procedures in rats. Binding potentials in the striatum were increased during chloral hydrate and ketamine anaesthesia and decreased during pentobarbital anaesthesia compared with awake rats. The authors postulated that the anaesthetics directly affected either CBF, physiological variables or dopamine D1 receptors. Alstrup et al. 27 compared the binding potentials of [11C]SCH23390 in Göttingen minipigs anaesthetized with either isoflurane or propofol. They found a higher binding of the tracer in most brain areas during isoflurane than during propofol anaesthesia, and postulated that the anaesthesia-dependent differences in tracer binding were most likely caused by different effects on CBF. Thus, major effects of anaesthesia were observed in both rats and minipigs; dopamine D1 receptor binding increased with ketamine and isoflurane, whereas it decreased with pentobarbital. Clearly, the dopamine D1 receptor is affected by anaesthesia, with the direction of change depending on the selected compound.
Dopamine D2/3 receptor
Many PET studies of anaesthesia effects on dopamine D2/3 have been published. Some of the studies compared anaesthetized and awake animals, while others compared different anaesthetics. Generally speaking, ketamine has been found to have minor effects on the dopamine D2/3 system, while inhalation anaesthetics tend to increase PET radioligand binding by dopamine D2/3 receptors. Nader et al. 61 determined whether the cerebral distribution of [18F]fluoroclebopride, a dopamine D2/3 receptor radioligand, was affected by anaesthesia in cynomolgus monkeys. They found no differences between isoflurane and ketamine on the uptake and washout of this tracer. Kobayashi et al. 62 used dopamine D2/3 antagonists [11C]-N-methylspiperone and [11C]raclopride to study anaesthetic effects using PET in rhesus monkeys; they found that binding of [11C]-N-methylspiperone was lower in striatum during isoflurane than during ketamine anaesthesia. Tsukada et al. 63 determined whether isoflurane affects the outcome of drug challenges on [11C]raclopride binding in the brain of anaesthetized and awake Macaca mulatta monkeys. They found that methamphetamine caused a marked release of dopamine in both anaesthetized and awake primates, whereas nicotine failed to affect dopamine release under either condition. Hassoun et al. 17 compared the binding of [11C]raclopride in awake and anaesthetized cats. They found that binding in striatum was unaffected by ketamine but was enhanced by halothane anaesthesia compared with the awake condition, and accounted for halothane's effect in terms of increased CBF. Patel et al. 26 also studied anaesthesia effects on the binding of [11C]raclopride. They found approximately the same striatal binding of [11C]raclopride in xylazine–ketamine anaesthetized rats as in freely-moving awake rats. Furthermore, drug challenges had similar effects on receptor binding during anaesthesia and in the awake condition. Alstrup et al. 28 also studied anaesthesia effects on [11C]raclopride binding in rats. They found that striatal [11C]raclopride binding was twice as high in rats anaesthetized with isoflurane than in rats given fentanyl–fluanisone–midazolam (i.e. the so-called Hypnorm–Dormicum rodent mixture). McCormick et al.64,65 also assessed effects of isoflurane on dopamine D2/3 receptor binding in rats. They noted increased binding of the dopamine D2/3 agonist [11C]-(+)-PHNO and enhanced inhibitory effects of amphetamine on [11C]-(+)-PHNO binding during isoflurane anaesthesia compared with awake rats.
Norepinephrine transporter
The norepinephrine transporter (NET) limits the duration of action of the neurotransmitter by removing it from the synapse. Blockade of NET has been viewed as an important mechanism in antidepressant properties of certain drugs. 55 One such drug, reboxetine, provided a starting point for a PET study exploring possible effects of anaesthesia on radioligand binding by NET. 66 Using rhesus monkeys, PET studies of radiolabelled analogues of reboxetine were carried out in animals that were either awake or anaesthetized with isoflurane. The awake monkeys were trained in a custom-made restraining device in order to minimize their potential stress and discomfort during PET imaging. 11 Accumulation of PET radioligands in monkey brain regions agreed with the anatomical location of NET, with relatively high levels in the thalamus, hypothalamus and raphe, and relatively low levels in the striatum.67,68 However, no marked differences were noted in PET findings between anaesthetized and awake animals.
Serotonin transporter
The serotonin transporter (SERT) is responsible for removing serotonin from the synaptic cleft, thereby limiting the actions of the neurotransmitter. SERT has been the focus of antidepressant drug development, and serotonin reuptake has been the target of numerous PET studies in laboratory animals and humans.9,69 Little is known, however, concerning whether PET findings on serotonin reuptake depend on anaesthetics. In fact, we are aware of only one PET study that addressed this question directly. 13 In that study, Stehouwer et al. found no reliable effect of isofluorane on PET radioligand binding to SERTs in cynomolgus monkeys.
Serotonin type 1A (5-HT1A) receptor
The serotonin type 1A (5-HT1A) receptor is a presynaptic and postsynaptic macromolecule that modulates cognitive and behavioural functions.70,71 Tokugawa et al. 72 determined the brain-to-plasma equilibrium ratio of the 5-HT1A PET tracer, [18F]FPWAY, in brain regions of rats that were either awake, partially immobilized or anaesthetized with isoflurane. They found that the distribution ratio of the radiotracer was higher in anaesthetized rats than in awake rats, and postulated that the elevated values produced by isoflurane may have been due to reduced endogenous serotonin secretion. Alternatively, postoperative pain and immobilization may have enhanced central serotonin release in awake rats, thereby altering regional CBF as well as the brain-to-plasma equilibrium ratio.73–77
Non-monoaminergic neurotransmission
General comments
Interest in central non-monoaminergic mechanisms is gradually increasing in response to increased demands for new and better treatments of brain disorders.49,78,79 Enzymatic regulation of central processes, excitatory actions of central glutamate and pharmacological properties of a novel receptor have recently received attention in studies of anaesthetic effects in laboratory animals.
Phosphodiesterase
Inhibition of phosphodiesterase has been considered as a potential site for antidepressant drug actions.80,81 Itoh et al. 82 studied the binding site density and tracer affinity of phosphodiesterase subtype 4 toward the PET radioligand [11C]-(R)-rolipram in awake rats and rats under isoflurane anaesthesia. Both binding site density and tracer affinity were lower during isoflurane anaesthesia than in awake rats.
Glutamatergic neurotransmission
Glutamate is a major excitatory neurotransmitter in the mammalian brain that interacts with several types of receptors. 83 The metabotropic glutamate receptor (mGluR) modulates excitatory actions of glutamate and has been implicated in neurological, psychiatric and neurodegenerative diseases. 83 Anaesthesia effects on glutamatergic neurotransmission were described in a report by Severance et al. 84 They used rats to determine whether anaesthesia played a crucial role in the distribution of [11C]MPEPy, a PET radioligand for mGluR, and found that urethane anaesthesia failed to affect binding of the PET radioligand in brain regions.
Cannabinoid receptor
The endocannabinoid system is an endogenous pathway involved in normal brain homeostatic mechanisms as well as some neuropsychiatric conditions. 85 Casteels et al. 86 assessed possible effects of pentobarbital and isoflurane anaesthesia on brain uptake of the type 1 cannabinoid receptor PET radioligand [18F]MK-9470 in rats. They found that the uptake of [18F]MK-9470 in whole brain failed to be affected by either anaesthesia. However, cortical uptakes of [18F]MK-9470 tended to be lower and cerebellum uptakes tended to be higher during pentobarbital and isoflurane anaesthesia than in awake rats.
Discussion and concluding remarks
As expected, most anaesthetics affect certain molecular mechanisms in the brain of laboratory animals. We must, therefore, exercise caution in the interpretation and generalization of findings obtained by PET brain imaging in anaesthetized animals. Needless to say, no final conclusion can be drawn concerning the ‘best’ general anaesthetic for PET neuroimaging in laboratory animals, since there are pros and cons of each procedure that require careful consideration in the planning of each research project. Firstly, we must note that effects of anaesthetics on brain mechanisms can be species-specific.33,87 Secondly, we must be aware that effects of anaesthetics on brain mechanisms are dosedependent.32,59 As a result, careful monitoring of anaesthetic levels and of vital functions is often required. Thirdly, too few studies have been carried out on possible effects of anaesthetics on most aspects of CBF, brain metabolism and neurotransmission to provide a solid basis by which to judge the reliability of findings.17,58 Ideally, confirmatory PET studies could be carried out in unanaesthetized, freely-moving animals, but that goal is currently out of reach in most cases.
We are currently reluctant to make general recommendations for the use of specific anaesthetics for future PET studies of brain mechanisms in laboratory animals, because each species and each experiment may have special requirements. We can, however, summarize the information reviewed here along with our research experience on PET brain imaging in laboratory animals as follows: (i) a mixture of ketamine and midazolam is well-suited for both induction of anaesthesia and brief scanning procedures, at least in pigs, (ii) isoflurane and halothane are usually well-suited for PET scanning procedures of intermediate length (e.g. 2–6 h), provided that physiological functions are carefully monitored and controlled, and (iii) propofol is well-suited both for intermediate and prolonged PET scanning. While the use of anaesthetics for PET brain imaging may have drawbacks due to their impact on physiological and pharmacological functions, we are confident that properly monitored anaesthesia will continue to provide suitable conditions for assessing diverse neuromolecular processes in the living brain of laboratory animals.