
Abstract
Summary
F1 hybrid pups from crosses between the strains 129SvEv/Crl and C57BL/6/Crl were subjected to an analysis of the development of adult swimming pattern (from the day of birth until 21 days old) to study the potential gender difference in neuromuscular development of neonatal and juvenile mice and the cognitive component in the development of swimming skills. Swimming as a parameter of scrutiny was chosen because it requires total coordination of the body's muscles, and we have previously demonstrated that the gradual change from a neonatal to an adult swimming pattern follows a fixed pattern, that can be scored objectively. Five different parameters were scored: the position of the head in the water, the use of front legs, the use of hind legs, the use of the tail as a rudder and whether or not the animals are able to maintain a straight course in the water. Each parameter could be objectively scored as 0 (neonatal), 1 (juvenile) or 2 (adult) level of development. There was no significant difference between development of locomotor skills in female and male pups. The maximum score obtained at any given day of development was not altered by learning from the previous daily swimming experiences. However, in individual swimming sessions, the time span between exposure to the water and display of maximum swimming score for the age was significantly shortened by daily exposure to water and swimming, indicating habituation to submersion in water. Startle reactions to water exposure could be minimized and finally eliminated by daily swimming sessions. This suggests a cognitive component limited, however, by the physical maturation of the nervous system and muscles, thus not resulting in acceleration of the development of swimming skills.
One of the most important challenges for functional genomics in the 21st century will be the systematic determination of the function of all genes in the mammalian genome and their role in disease. The mouse will play a pivotal role in achieving this endeavour. Thus, there is intense activity in the development of tools for improving phenotyping of genetically modified mice (see e.g. www.eumodic.org and www.eumorphia.org).
Swimming requires the smooth integrated organization of a coordinated series of reflex responses including the righting reflex, vestibular reflexes and extensor reflexes (Pavlovska-Teglia et al. 1994). This makes swimming suitable for assessing the functional and behavioural development of the sensory–muscular system in neonatal and juvenile rodents (Young et al. 1996). In young inbred mice, the development of adult mode of swimming is accomplished by Day 20 (Day 15 in F1 hybrids) at which time the juvenile mouse has a fully developed adult swimming pattern (Adencreutz & Hau 2004). It is quite easy to objectively characterize the normal development of the adult pattern from Days 1 to 20 by assessing five parameters: head position in water, use of front limbs, use of hindlimbs, use of tail and line of swimming.
In mice, life expectancy of many gene-modified strains is shorter than that of the parent strains (Morton & Hau 2003), but it is often difficult to detect phenotypic changes from normal in terms of normal development and performance in various test systems. Phenotypic characterization methods for gene-modified mice including motor function analyses are of importance, perhaps, in particular for mutants developed to study ataxias and other neuromuscular diseases. We have recently demonstrated that the pattern of how juvenile mice develop an adult mode of swimming varies between strains, but is very stable within a strain, and also very susceptible to changes in neuromuscular function and development (Adencreutz & Hau 2004).
In the production of mutant mice via microinjection of gene-targeted embryonic stem (ES) cells C57BL/6 (C57) mice are often used as blastocyst donors and 129Sv (129) mice routinely used for ES cells (Rubin et al. 2003). In the present study, we mated 129 males with C57 females to obtain F1 hybrid pups, which develop swimming skills more quickly than the parental strains (Adencreutz & Hau 2004). C57 mice can learn by observation (Carlier & Jamon 2006), and differences in learning abilities for different tasks between C57 and 129 mice have been described in water maze studies (Wolff et al. 2002). Gender-related differences in the performance in water mazes of young adult mice have been described (Arters et al. 1998).
Consequently, in order to assess the robustness and general usefulness of the mouse swim test to reveal deficiencies in neuromuscular development, the present study was designed to determine if there was a gender difference in the development of the adult swimming pattern. In addition, whether there was a cognitive component in the development of swimming skills, such that neonatal and juvenile mice that have been subjected to swimming every day may develop an adult swimming pattern quicker than mice of the same age that have never been exposed to water previously.
Materials and Methods
Animals and husbandry
F1 hybrid offspring from crosses between the strains 129S2/SvPasCrl males and C57BL/6NCrl (Charles River Laboratories, Wiga, Germany) were studied. The mice were of SPF quality and maintained in a conventional animal facility. On arrival, the parents (7 weeks old nulliparous) were caged in polycarbonate type III cages (Scanbur, Koge, Denmark), three females together with one male. A total of 25 cages were used, three for the experiment group, i.e. the pups subjected to swimming every day and the rest for control groups. Pregnant females were separated and housed individually in type III cages (Scanbur). A total of 27 pups (12 males and 15 females) from three litters were subjected to swimming every day throughout the study period (experiment group) and 220 pups (22 × 10 pups, 5 males and 5 females) from the remaining 22 litters were chosen to serve as controls and were thus subjected to water and swimming only once. They were chosen so that there would be 10 naïve pups on each day throughout the study period.
The mice were kept in standard animal rooms and subjected to standard animal house conditions: the light regime was 12/12 h dark/artificial light cycle, temperature was maintained at 21 ± 1°C, the relative humidity was between 30 and 60%, and the cages were cleaned twice a week. Aspen chips (Tapvei, Oy, Kortteinen, Finland) were used as bedding and aspen hides used as environmental enrichment. Water and standard rodent pelleted diet (Brogaarden, Gentofte, Denmark) were available ad libitum.
Experimental procedures
Swim tests were carried out daily, in a room adjacent to the animal holding room, from Days 1 to 21 (Day 0 being the day when the pups had been born during the night) in the dark phase between 20:00 and 22:00 h in an aquarium approximately 40 × 40 × 50 cm containing water at a depth of 20 cm maintained at 30–34°C. The observations were done using a torch and the light beam of this was not visible to mice not subjected to swimming. Mice younger than seven days were left in the water for approximately 10 s and older mice for 20–60 s, depending on their stamina. After each swimming session the pups were dried with paper tissue, before being returned to their mothers. The behaviour of the young and the mothers seemed unaffected by the procedures. When a pup subjected to swimming was returned to the mother and the other pups in the litter, the mother immediately concentrated her attention on this pup and licked it. Swimming ability was evaluated by five rating systems as described by Adencreutz and Hau (2004) (Table 1).
Score assessment scale for scoring swimming ability in mice

During the swimming analysis, the general behaviour in water and the overall swimming capability were also assessed. This included how the mice reacted when placed in the water (i.e. startle reactions, swimming or immobility) and how the final adult swimming behaviour differed between the groups (i.e. body angle in the water, fatigue and overall behaviour).
Ethics committee approval
The relevant Danish authorities did not consider this study subject to licensing. Throughout the study, the mice were never subjected to longer periods of swimming, which would have resulted in fatigue and potential stress and suffering.
Statistics
The data were analysed using Friedman two-way analysis of variance and the MINITAB statistics program (CLE COM Ltd, Birmingham, UK). The scores in each swimming test were tested on each day of observation for significant differences between the four groups of pups. P values <0.05 were considered significant.
Results
Head position
The males in the experiment group (subjected to swimming every day throughout the study period) started to raise their heads on Day 7 and all had reached the adult head position on Day 13. The females in the experiment group first showed score 1 signs on Day 8 and were somewhat less mature but caught up and reached the adult head position already on Day 12. The first male pups of the control group subjected to swimming only once raised the head to water level on Day 7 and on Day 11 all were displaying the adult head position. The female pups in the control group also began to raise their heads above water level on Day 7 and were all displaying adult head position on Day 11. There were no significant differences between the four groups (Figure 1).
Male and female experiment are pups subjected to swimming every day throughout the study period. Male and female controls are pups subjected to swimming on one day only. Average of cumulative score (see Table 1 for details on the scoring system applied) for the pups of the groups tested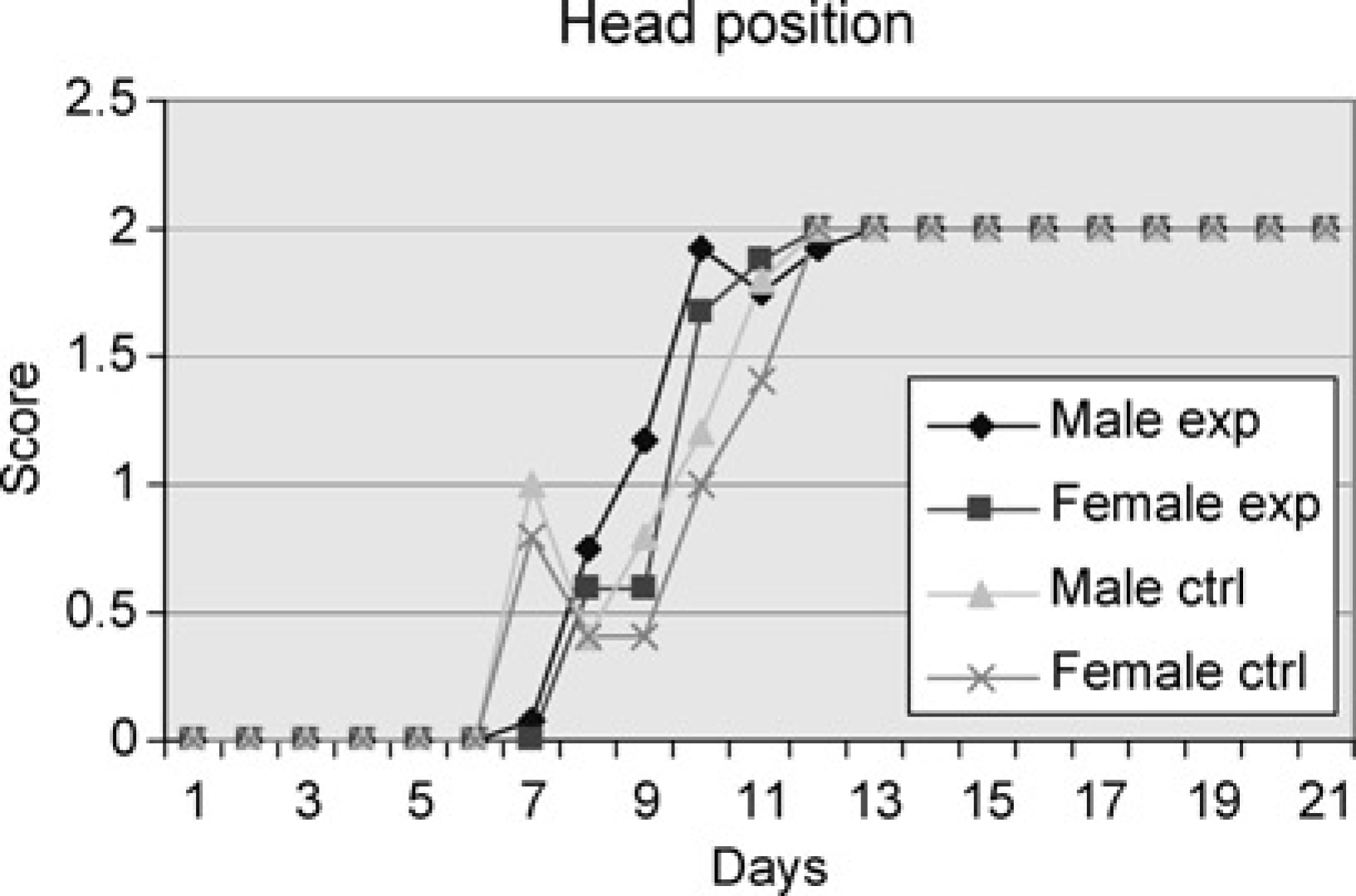
Front leg movement
During swimming tests partial, and then total, cessation of the use of forelimbs as in the adult mode of swimming developed as follows. The males in the experimental group began to show score 1 signs of a mixed front leg swimming pattern on Day 9 and all displayed adult front leg movement by Day 15. The females of the experimental group first showed score 1 signs on Day 9 and all had reached score 2 on Day 15. The male pups of the control group subjected to swimming only once began to show development on Day 9 and achieved the adult front leg position on Day 14. The female pups in the control group showed development already on Day 8 but had not completed development until Day 15, which was similar to the experimental group. There were no significant differences between the four groups (Figure 2).
Male and female experiment are pups subjected to swimming every day throughout the study period. Male and female controls are pups subjected to swimming on one day only. Average of cumulative score (Table 1) for the pups of the groups tested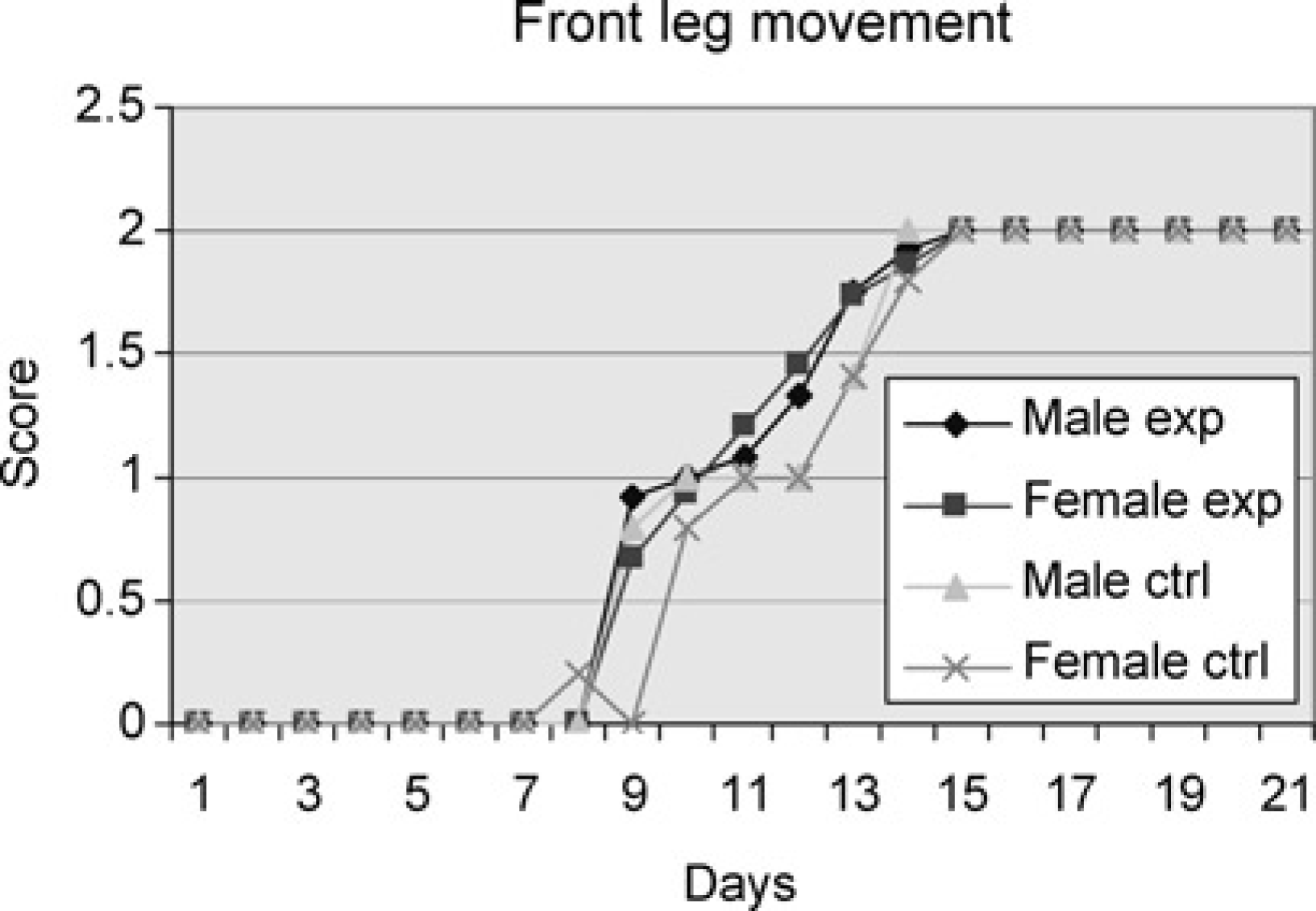
Hind leg movement
The males in the experimental group began to show score signs of a mixed hind leg swimming pattern on Day 9 and all had achieved adult hind leg use by Day 12. The females of the same group first showed score signs on Day 9 and all quickly reached the adult use of the hind legs by Day 12. In the control group, male pups began development on Day 8 and achieved the adult hind leg use on Day 11. The females also commenced development on Day 8 but were not finished developing until Day 12. There were no significant differences between the four groups (Figure 3).
Male and female experiment are pups subjected to swimming every day throughout the study period. Male and female controls are pups subjected to swimming on one day only. Average of cumulative score (Table 1) for the pups of the groups tested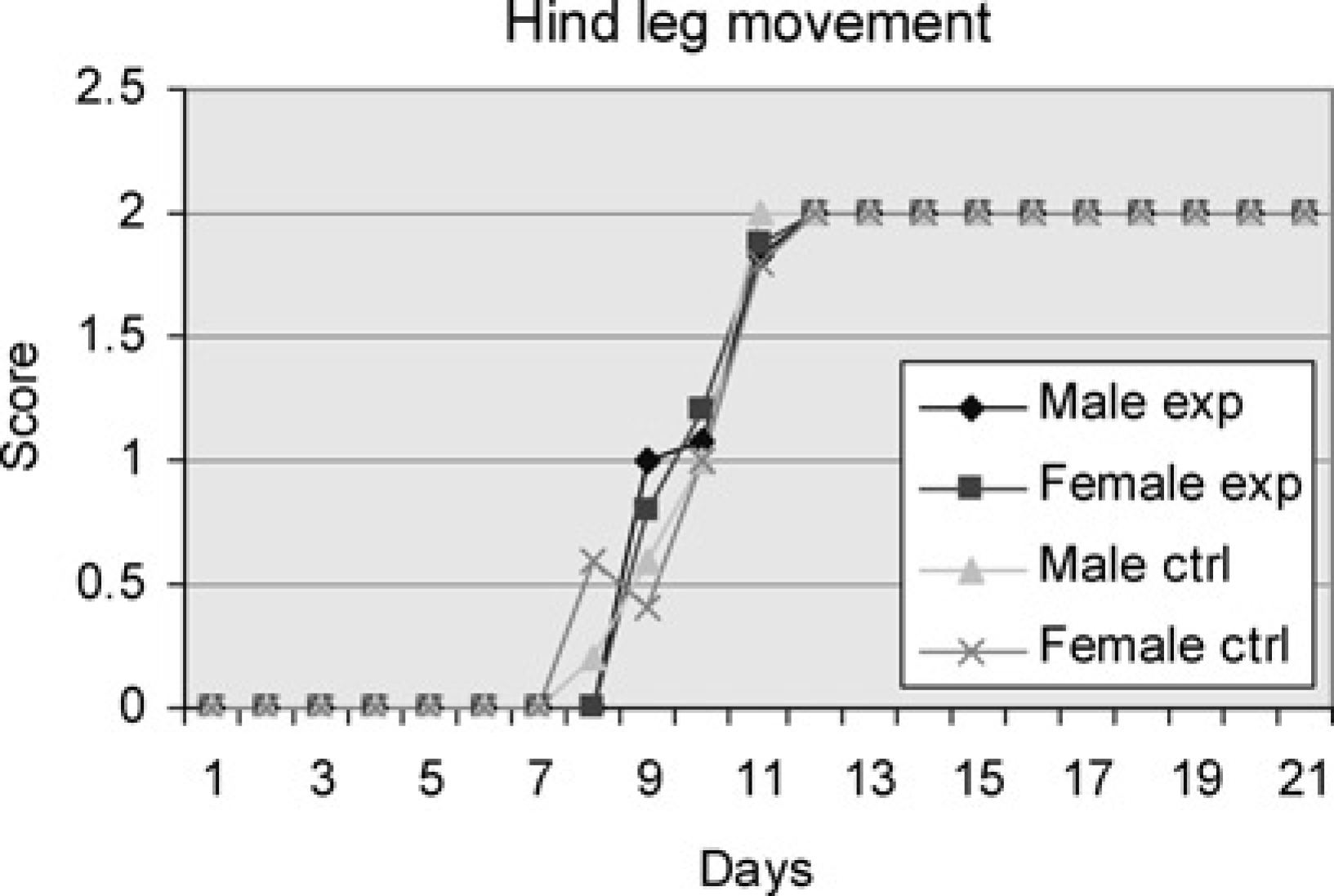
Tail movement
In the beginning, the pups did not use their tails at all, after which they began to move their tails like a propeller with circular movements in the water. They then progressed to lateral tail movements as in the juvenile and adult swimming patterns. The males of the experimental group started to show score signs on propeller tail movement on Day 3 and all displayed adult tail movement on Day 14. The females of the same group also first showed score signs on Day 3 and all reached score 2 on Day 14. One of the male pups in the control group began development on Day 2, some more on day 3 and all achieved the adult movement pattern of the tail by Day 11. Also in the female group one pup began development already on Day 2, some more on Day 3 but not all were exhibiting the adult pattern until Day 12. There were no significant differences between the four groups (Figure 4).
Male and female experiment are pups subjected to swimming every day throughout the study period. Male and female controls are pups subjected to swimming on one day only. Average of cumulative score (Table 1) for the pups of the groups tested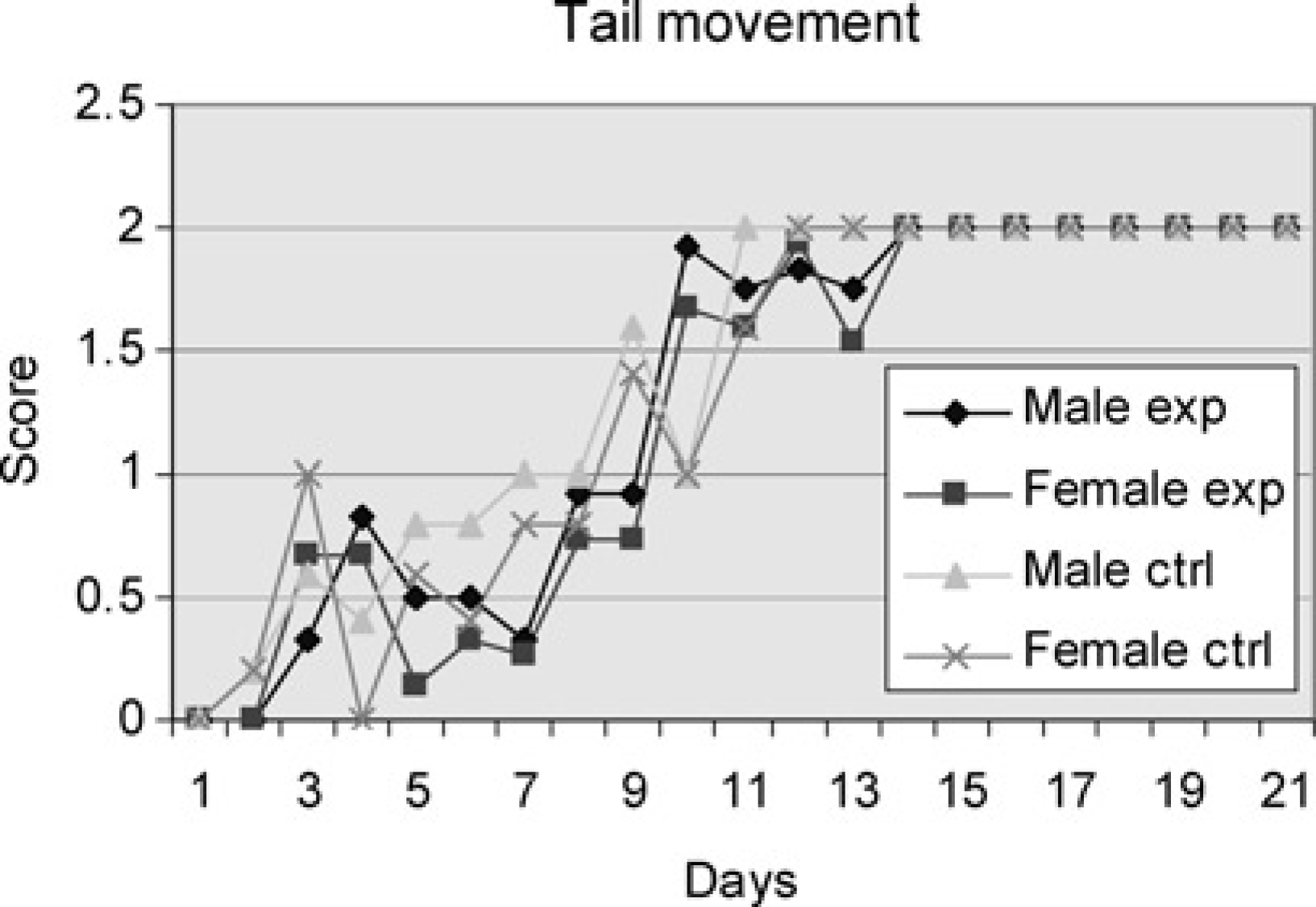
Line of movement
Orientation in space, posture and coordination of the locomotor activities are the components that determine the line of swimming. In the beginning of the study period, the pups swam in small circles. The diameter of the circles progressively grew larger as they began to maintain the nose above the water and with their strength developing. By the time the animals attempted to swim in straight lines, they sometimes changed direction and began to swim in circles in the opposite direction. The males in the experimental group began to show score signs on breaking the circular swimming on Day 3 and all had an adult straight purposeful mode of swimming on Day 12. The females of this group also first showed development on Day 3 and all quickly finished development by Day 11. In the control group, the males began development on Day 3 and were finished developing on day 10, whereas the first females of this group started breaking the circular swimming already on Day 2, but not all achieved the adult swimming pattern until Day 12. There were no significant differences between the four groups (Figure 5).
Male and female experiment are pups subjected to swimming every day throughout the study period. Male and female controls are pups subjected to swimming on one day only. Average of cumulative score (Table 1) for the pups of the groups tested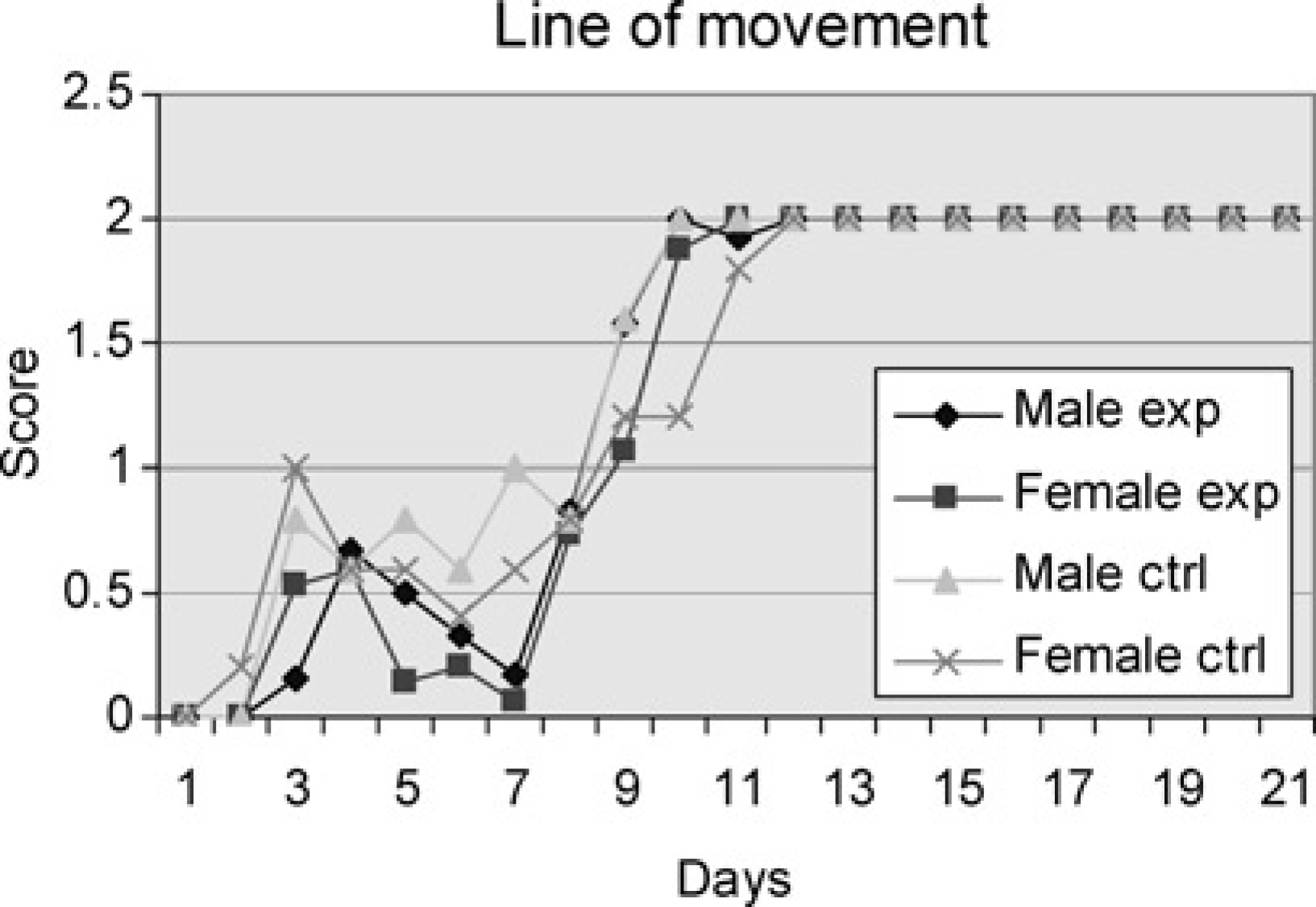
Total locomotor development
When the total sums of the five rating systems were added, the males of the experimental group began to show score signs of a juvenile swimming pattern on Day 3 and all had achieved adult swimming pattern by Day 15. The first females in the experimental group also showed score signs on Day 3 and all had also reached score 2 by Day 15. The male pups of the control group began development on Day 2 but not all achieved the adult pattern until Day 14. The first female pups of the control group also commenced development by Day 2 but did not finish development until Day 15. There were no significant differences between the four groups (Figure 6).
Differences in swimming patterns
The study revealed no significant differences in swimming pattern between the two genders nor between the mice that had been exposed to swimming every day as opposed to the mice of the same age that had never been exposed to water before. It was notable, however, that the pups that were exposed to swimming every day, males as well as females, were more used to being handled and did not startle as much when introduced to the water as did the swimming-naïve pups. The experienced pups also reached their maximum score for the day much faster than the pups of the control group. In some cases within 5 s as opposed to up to 20–30 s for a first-time swimmer, during which time the pup would exhibit startle reactions and would be swimming in a more primitive and confused manner than after some time. This period is shortened notably if the pup is made to swim every day. The everyday swimming pup would start out with a minor startle reaction, initially exhibit a more primitive swimming behaviour similar to the day before, but would soon achieve a more adult way of swimming than the day before.
Male and female experiment are pups subjected to swimming every day throughout the study period. Male and female controls are pups subjected to swimming on one day only. Total sums of the five rating systems (Table 1) for the pups of the groups tested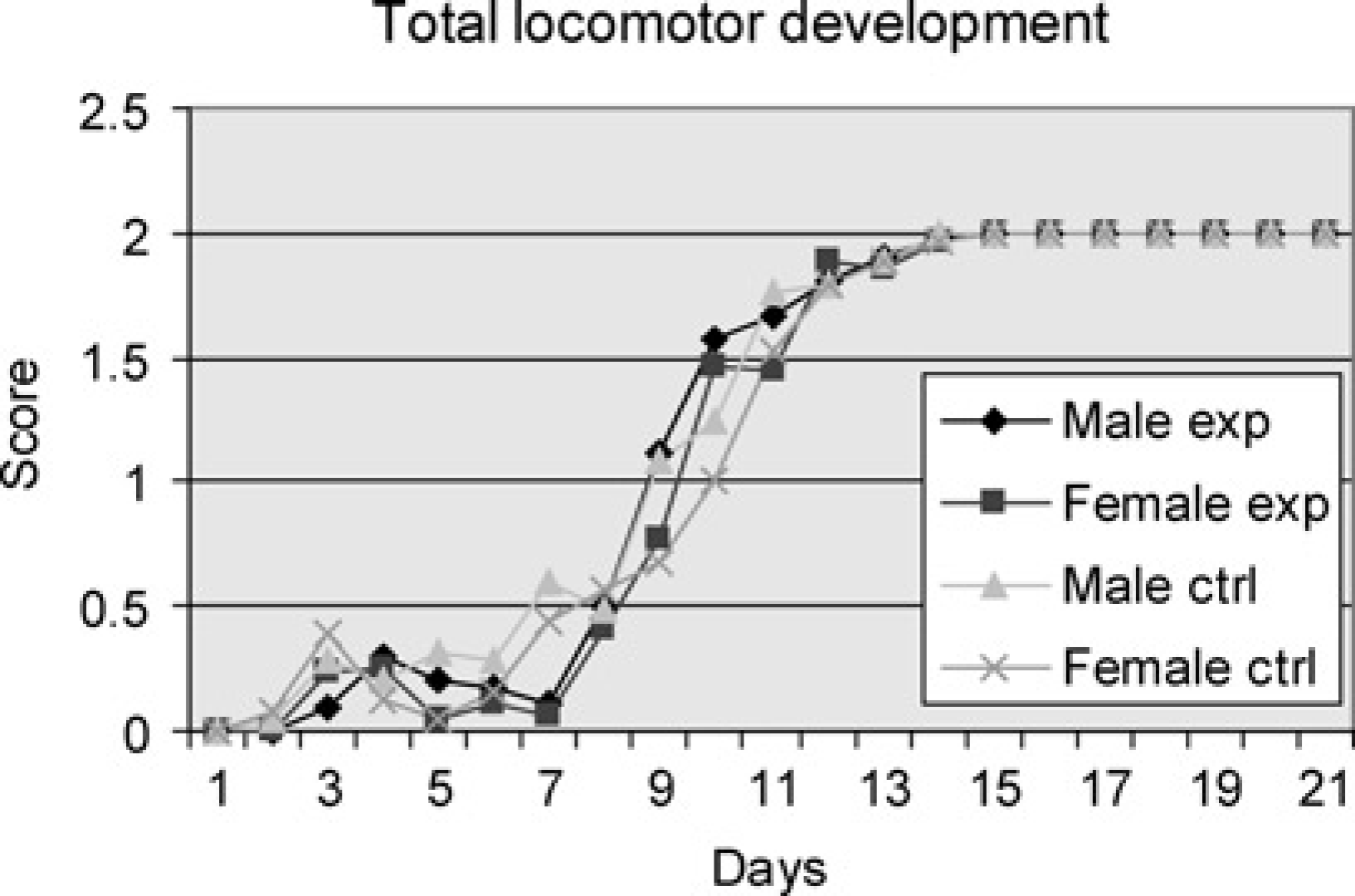
Discussion
Locomotor activity is a common parameter used to characterize the behavioural phenotype of mice. The C57 mouse strains differ markedly from 129 mouse strains with respect to locomotor behaviour variables (Paulus et al. 1999). There is evidence that C57 mice can learn by observation and in doing so there seems to be no sex difference (Carlier & Jamon 2006). Strain differences have been described for the behaviour of mice in Morris water maze (Wahlsten et al. 2005). Although C57 mice are faster swimmers than 129 mice they display no difference in the path length in Morris’ water maze tests and show a more pronounced startle reaction than do 129 mice (Contet et al. 2001). This agrees with the findings that C57 mice exhibit a profound startle reaction when put into water when compared with 129 mice (Adencreutz & Hau 2004). This is in accordance with the findings that the autonomic stress response is the highest in C57 mice as compared with 129 mice and Swiss Webster mice when heart rate and body temperature were measured (van Bogaert et al. 2006). Corticosteroids influence rodent behaviour and CNS development (Brain 1979), and stress may facilitate learning in mice as shown in another study (Peeters & Broekkamp 1994) and there is some indication that diet has an impact on the learning abilities of mice (Frances et al. 1995). A difference in learning abilities for different tasks between C57 and 129 mice has also been reported in a study of performance in three water maze protocols (Wolff et al. 2002). Previous studies have revealed a sex difference in the performance in the spatial water maze of young adult mice, and certain lesions in nucleus basalis magnocellularis, that provides the primary source of cholinergic input to the cortex, produced a spatial navigation deficit affecting only males in the water maze (Arters et al. 1998). It has also been demonstrated that 5-HT (serotonin) plays a vital part in the maturation of the neuromuscular system, as monoamine oxidase A-deficient mice had impaired hind leg use when swimming (Cazalets et al. 2000).
Adult mice of the strain 129SvEv/Bkl are strong and confident swimmers. By contrast, the C57BL/6/Bkl adults show difficulties with pronounced startle reactions when introduced into the water and tire quickly (Adencreutz & Hau 2004). F1 hybrid pups (offspring of C57 × 129 crosses) develop an adult mode of swimming significantly faster than pups of the two parental strains, clearly demonstrating a heterosis effect (Adencreutz & Hau 2004).
The present study demonstrated that there were no significant differences between the four groups of mice tested, with respect to the temporal development of an adult mode of swimming. In behavioural tests, gender differences are common (Astur et al. 2004), but we found in this study that there was no gender difference in juvenile mice in the development of swim pattern, and that previous exposure to water environment does not accelerate the development of the adult swim pattern. Previous experiments on training in 2-month-old mice have demonstrated that training is beneficial for the performance in water mazes (Vicens et al. 1999), but in our study training had no significant effect on the developing nervous system in animals younger than three weeks.
There was, however, an indication of a cognitive component in the juvenile mice. The time span between the moment the pup was placed in the water until it displayed swimming to the best of its ability for its developmental stage was significantly shortened by daily exposure to water and swimming. Startle reactions were also minimized suggesting a cognitive component limited by the physical growth and maturation of the nervous system and the muscles in the body. The myelinization of the axons cannot be forced by swimming, but there was a clear memory of the behaviour from the day before and this obviously helped the pup to more quickly regain the behaviour from the day before. There is also likely a recognition of the water environment as the startle reaction was decreasing in experienced pups compared with pups exposed to water for the first time. To measure this, another type of scoring must be developed, that takes into account the time elapsed from exposure to water until the maximum score is obtained, and to which degree the pup exhibits a startle reaction.
The genotypes chosen for the present study are widely used in the production of genetically-modified mice. Considering that many of these are developed with the aim to create models to study disorders, it seems important to characterize potential gender differences and learning abilities in normal locomotor and neuromuscular development in the 129 and C57BL strains as well as crosses between them.
From a practical point of view, it is fortunate that there were no observed gender differences in locomotor development and no significant difference in the development stages of swimming ability between first-time swimmers and pups exposed to water daily throughout the juvenile period. This makes it feasible to use this method for an easy testing system for phenotyping genetically-modified animals. These can, for example, be subjected to the swim test on a specific day and if affected by a locomotor or a neurogical deficit this would be revealed by the test regardless of the gender of the mouse, ambient environment and previous experiences.
Studies are in progress on the efficacy of the present method to reveal genetic modifications affecting neuromuscular coordination and development.
In conclusion, the present study demonstrates for the first time that there was no gender difference in the temporal development of an adult swimming pattern in neonatal and juvenile mice. The developmental stages towards an adult swimming pattern in neonatal and juvenile mice subjected to swimming every day from birth and onwards did not differ from those of age-matched controls not subjected to swimming more than one single day during this period.
Footnotes
Acknowledgement
The authors are grateful for the financial support of FoU Sundsvall, Västernorrlands läns landsting, Sweden.