Mice were infected experimentally and subclinically with Corynebacterium kutscheri to recover the organism from mice faeces. The faeces were then cultured using selective furazolidone-nalidixic acid-colimycin agar. The number of C. kutscheri per gram of fresh faeces varied from mouse to mouse, but once established in the intestine, the organism was excreted in the faeces for at least five months. Viable bacteria were detected in most of the faecal samples, including those stored in the animal room for five days. The number of organisms in the stored faeces decreased gradually but did not differ significantly from those in the fresh faeces until they had been stored for more than three days. Many infected mice excreted between 104.77 and 105.37 colony forming units (CFU) of C. kutscheri per day in their faeces, and one mouse even excreted 103.74 CFU at eight weeks postinfection. These values showed little daily variation. Our present study showed that subclinically infected mice discharged the organism continuously and persistently in their faeces. Therefore, faecal samples would be useful for monitoring infection with C. kutscheri in living mice in a manner that is not stressful for the animals.
Natural infection with Corynebacterium kutscheri is usually subclinical and occurs in conventionally reared mice and rats. Infection with C. kutscheri results in pseudotuberculosis only after severe immunosuppression of the host (Fauve et al. 1964, Fox et al. 1987). We have established simple procedures for the detection of subclinical infections with C. kutscheri in mice, rats and Syrian hamsters using a selective medium (Amao et al. 1990, 1991, 1995a,b). We have also shown that subclinically infected mice harboured C. kutscheri in the oral cavity, caecum, colon and rectum. These locations suggested that the organism is likely transmitted via a faecal–oral route. However, the modes of excretion and spreading of the agent are not fully known, and a detailed bacterial study of faeces in relation to organism colonization has not been carried out. If bacterial isolation from faeces is possible and reliable, faecal samples could be useful as a non-invasive method for detecting subclinical infections with C. kutscheri. A quantitative study of faeces may be an indicator of stress, because the number of C. kutscheri in animal intestines increases in immunosuppressed hosts. Hence, as a primary study, we attempted to isolate and count C. kutscheri in faeces collected from experimentally and subclinically infected mice.
Materials and methods
Animals
Three-week-old male Slc:ICR-specific pathogen-free mice (Japan SLC, Inc, Shizuoka, Japan) were purchased and given one week for acclimatization to the study conditions. The mice did not possess serum antibodies to Clostridium piliforme, Mycoplasma pulmonis, haemagglutinating virus of Japan (HVJ) or mouse hepatitis virus (MHV). They were also negative for Pseudomonas aeruginosa, Salmonella spp., Pasteurella pneumotropica, Citrobacter rodentium, Mycoplasma spp., dermatophytes, Giardia spp., Spironucleus muris, Syphacia spp. and C. kutscheri. All the mice were housed in polycarbonate cages (W22 × D33 × H13 cm), with five animals per cage until 14 days postinfection, at which time they were placed in individual wire cages (W13 × D20 × H13 cm), so that C. kutscheri could be recovered from their faeces. All the mice were housed in an absorbed air isolation rack (Tokiwa Kagaku Kikai Co, Tokyo, Japan) in an animal room under controlled conditions of 20–25°C, 40–70% relative humidity, air ventilation (12 times/h) and a 14 h (05:00–19:00 h) lighting cycle. Commercial diet (MB-1; Funabashi Farm Co Ltd, Chiba, Japan) and water were provided ad libitum.
Preparation and inoculation of C. kutscheri
The strain S5L (Amao et al. 1990) of C. kutscheri, originally isolated from a lung abscess in a rat, was used. A frozen stock culture of the strain was thawed and cultured on heart infusion agar (HIA; Eiken Chemical Co Ltd, Tokyo, Japan) for 48 h at 37°C in air. Serial 10-fold dilutions of the bacterial suspension were then prepared in heart infusion broth (HIB; Eiken Chemical Co Ltd) for the enumeration of colony forming units (CFU) using HIA. Ten mice were given 0.2 mL of bacterial suspension containing 1.52 × 108 CFU of C. kutscheri, as measured using a viable colony count. The suspension was inoculated via an intrastomach injection through a metal stomach tube. The animals used in this study were treated in accordance with the provisions for animal welfare of the Nippon Veterinary and Life Science University, which has adopted the Guidelines for Animal Experimentation issued by the Japanese Association for Laboratory Animal Science (Japanese Association for Laboratory Animal Science 1987).
Bacteriology
A selective medium, furazolidone-nalidixic acid-colimycin agar (FNC agar) (Amao et al. 1990), was used to isolate C. kutscheri. This medium inhibited the growth of Gram-negative rods, P. aeruginosa, Proteus sp. and Gram-positive cocci but did not affect the growth of C. kutscheri. Samples of faeces were homogenized using a glass mortar in 10 or 100 volumes of sterile HIB. Serial 10-fold ranges of the homogenate dilutions were prepared in HIB, and a 0.05 mL aliquot of each dilution was placed on FNC agar. After incubating the agar plates at 37°C for 72 h in air, corynebacterium-like colonies were isolated and subjected to slide agglutination tests using rabbit antiserum against strain S5L of C. kutscheri (Amao et al. 1990). Strain S5L of C. kutscheri was used as a positive control in the test. Briefly, one drop of the C. kutscheri antiserum and one drop of saline were placed on each side of a partition on a slide. A small amount of bacterium was then added and mixed with each drop using a loop, and the slide was rotated by hand for 30 s. A coarse aggregation of bacterial cells with a clear background was considered a positive agglutination result. A homogenous emulsion of bacterial cells was interpreted as a negative result. Isolates that tested positive were further identified by Gram staining and their biochemical properties (Amao et al. 1991). C. kutscheri colonies were counted, and the averages from duplicate experiments were used to calculate the number of bacteria (CFU/g).
Long-term excretion of C. kutscheri in faeces
A survey of the long-term excretion of C. kutscheri in the faeces of 8–10 infected mice was conducted by examining faecal samples collected two weeks, two months or five months postinfection. Ten fresh faecal pellets per mouse were collected, homogenized, and tested using a quantitative culture method.
Number of C. kutscheri per gram of individual fresh faeces
To quantify the excreted C. kutscheri, faecal samples from two mice were examined at five months postinfection. Fresh faecal pellets were collected at between 10:00–12:00 h, homogenized, and the number of C. kutscheri was quantified. Twenty-four faecal pellets per mouse were examined for this experiment.
Variations in the number of organisms in the faecal samples after storage
To examine the variation in the number of C. kutscheri isolated from stored faecal samples, fresh faecal samples were divided into two halves. One-half was tested immediately, and the other half was tested after 0, 1, 2, 3 or 5 days of storage at room temperature. The faecal samples were stored in an open container in an absorbed air isolation rack. On each of the examination days, six faecal samples from three infected mice were examined using a quantitative culture.
Numbers of C. kutscheri per gram of total faeces collected during a 24 h period
To examine the number of C. kutscheri per gram of total faeces during a 24 h period, faecal sampling was performed on five consecutive days in five infected mice, starting at eight weeks postinfection. All the faeces were collected from the animals' bedding at 12:00 h every day and were then homogenized and cultured immediately.
Impression culture of C. kutscheri from faeces
Comparison of a direct preparation culture with a quantitative culture using the same faecal sample was attempted. This test was conducted using faecal samples obtained from 10 infected mice at two months postinfection. A fresh faecal pellet was cut in half, and the cut surface was firmly pressed onto FNC agar in one place and then further spread using a loop. The other half of the pellet was homogenized in 100 volumes of sterile HIB using a glass mortar and processed using the quantitative culture method described above.
Statistical analysis
A student's t-test was used to compare the mean numbers of organisms per gram of homogenated faeces.
Results
None of the mice manifested clinical signs of murine corynebacteriosis.
Long-term excretion of C. kutscheri in faeces
The organisms were detected in faeces from all 10 mice at two weeks and two months postinfection and in seven of the eight mice at five months postinfection (data not shown). C. kutscheri was not detected in 10 faecal samples from five non-infected mice that were cared for and managed in the same absorbed air isolation rack as that used during the experiment.
Number of C. kutscheri per gram of individual fresh faeces
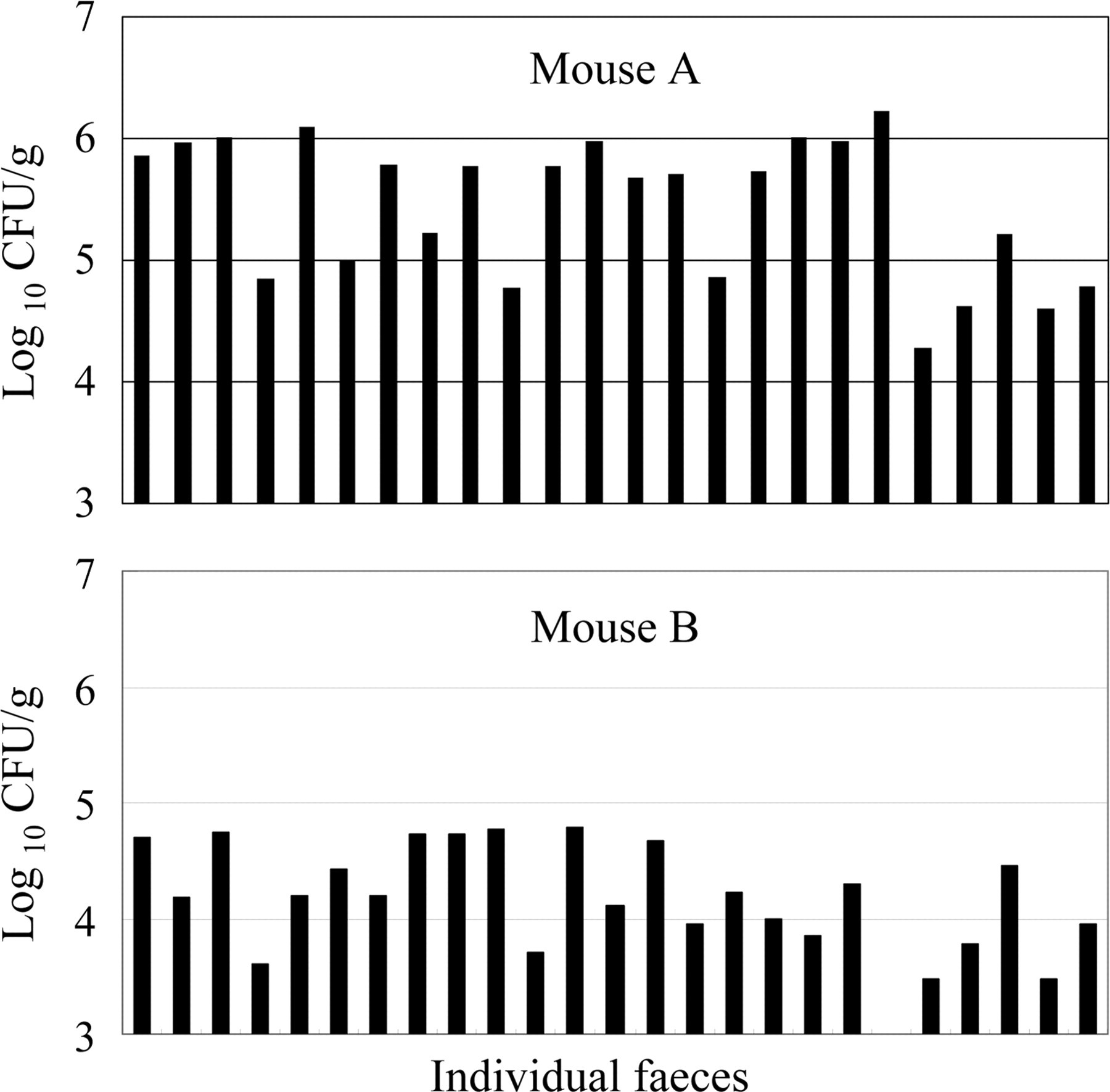
In one mouse whose faecal samples showed a high number of organisms, C. kutscheri was detected in every sample (104.28 –106.23 CFU/g, Figure 1A). In another mouse whose faecal samples showed a low bacterial number, the organism was detected in 23 of 24 samples (95.8%, 103.50–104.79 CFU/g, Figure 1B).
Numbers of Corynebacterium kutscheri in fresh faeces. Fresh faeces from individually housed mice were obtained between 10:00–12:00 h and were homogenized and tested for the number of C. kutscheri. The numbers of organisms recovered from mouse A were relatively high, while those from mouse B were relatively low
Variations in the number of organisms in the faecal samples after storage
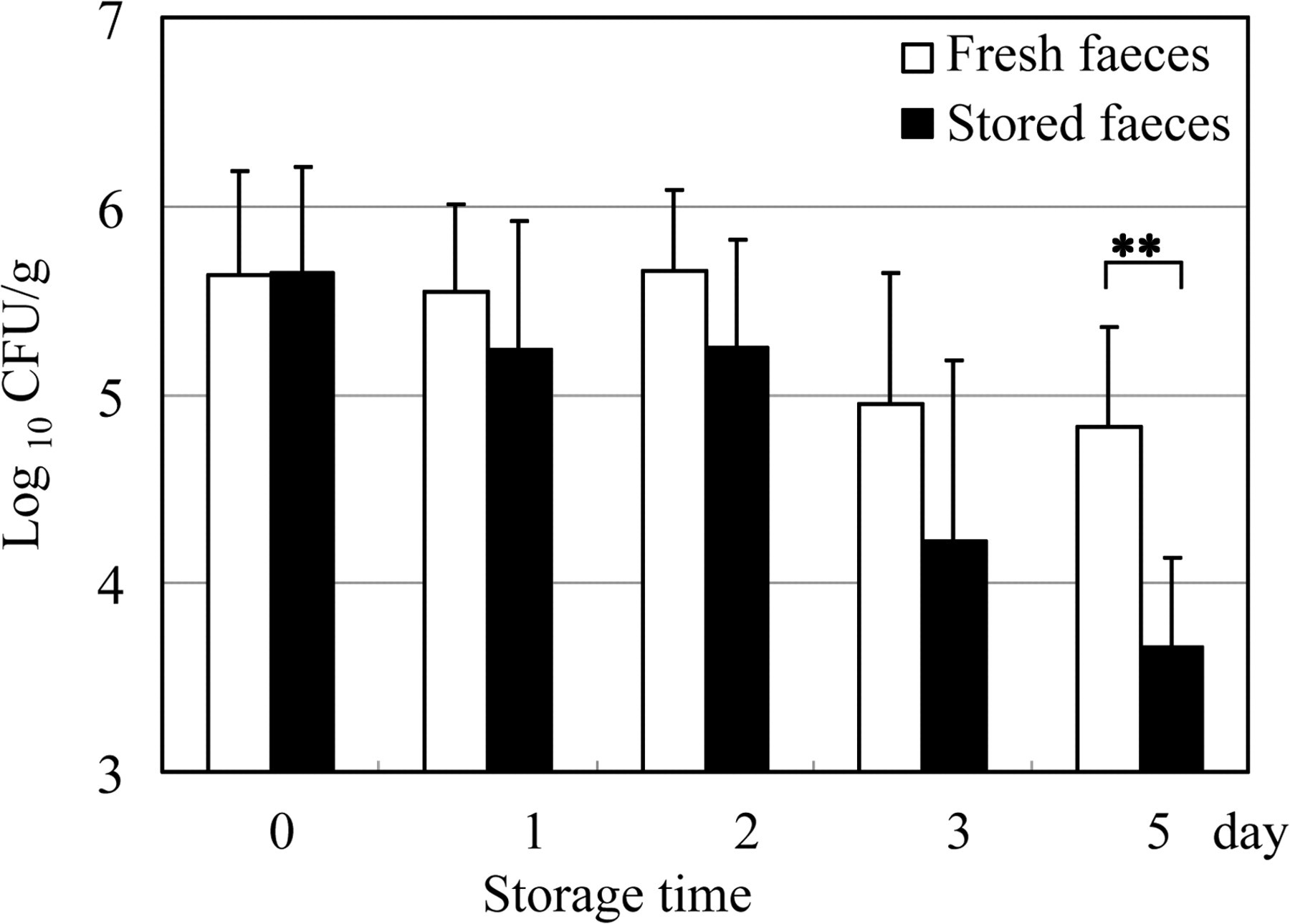
The mean numbers of C. kutscheri per gram of faeces are shown in Figure 2. The organisms were detected in 29 of 30 samples, including those that had been stored for five days (96.7%). The number of organisms decreased gradually with the length of storage, but no significant difference between the fresh and stored faecal samples was observed until after three days. The number of organisms in the samples that had been stored for five days was significantly lower than that in the fresh faecal samples (P < 0.01).
Variations in the number of Corynebacterium kutscheri in stored faeces. Fresh faecal samples were divided in half; one-half was tested immediately, and the other half was tested after storage for 0, 1, 2, 3 or 5 days. Each value indicates the mean ± standard deviation of six faecal pellets from three infected mice. **P < 0.01
Numbers of C. kutscheri per gram of total faeces collected during a 24 h period
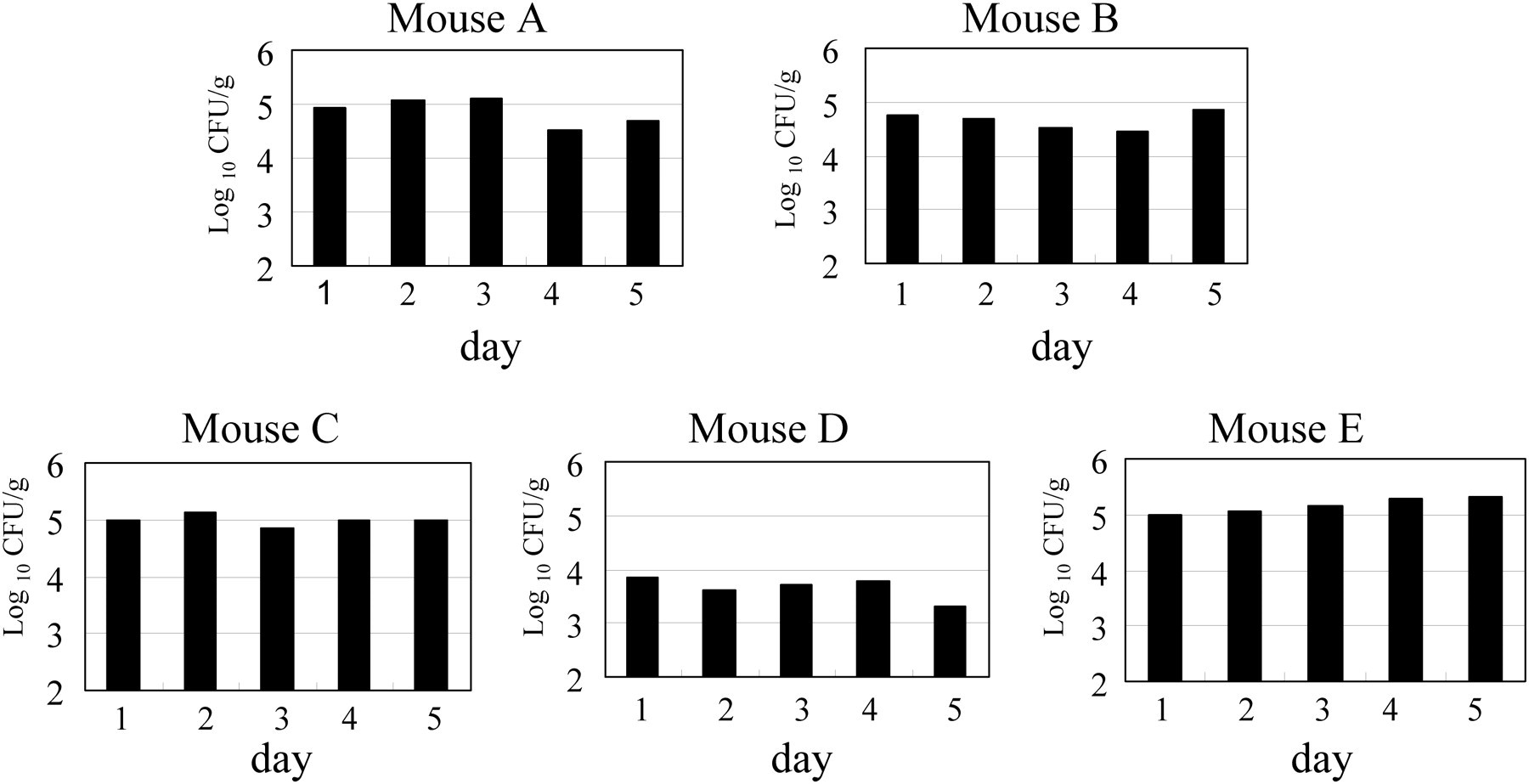
The numbers of C. kutscheri per gram of total faeces collected during a 24 h period are shown in Figure 3. Between 104.46 and 105.33 CFU/g of C. kutscheri were observed in the faecal samples from mice A, B, C and E, but the numbers in mouse D were lower (103.30–103.85 CFU/g). No remarkable daily variation was observed in any animal. In these mice, the mean faecal weights were between 1256 and 1635 mg per day. Thus, mice A, B, C and E excreted about 104.77–105.37 CFU per day and mouse D excreted about 103.74 CFU per day (data not shown).
Numbers of Corynebacterium kutscheri per gram of total faeces collected during 24 h periods
Impression culture of C. kutscheri from faeces
The results of the impression cultures are shown in Table 1. C. kutscheri was detected from all the faecal samples examined using quantitative cultures (103.30–105.11 CFU/g) but was not detected in direct impression cultures of two out of 10 faecal pellets with low numbers of organisms (below 103.80 CFU/g).
Isolation of Corynebacterium kutscheri from faeces: comparison of direct impression cultures and quantitative cultures
No. of animal
Quantitative culture (CFU/g)
Impression culture*
1
105.11
++
2
104.00
+
3
104.59
+
4
103.78
–
5
104.46
+
6
103.30
–
7
104.54
++
8
104.40
++
9
104.99
++
10
103.90
+
*Number of C. kutscheri colonies
–: negative; +: 1–10; ++: 11–50
CFU: colony forming units
Discussion
This is the first report describing the detailed isolation of C. kutscheri from faeces in subclinically infected mice. C. kutscheri was isolated from fresh faeces obtained from seven out of eight experimentally infected mice at two weeks to five months postinfection, suggesting that colonization was persistent whether the faecal counts were low (103.0–105.0 CFU/g) or high (104.0–106.0 CFU/g). Using a rifampicin-resistant C. kutscheri strain, Yokoiyama et al. (1975) found that mice infected perorally began to excrete the organisms in their faeces within 6 h and that most of the mice continued to carry the organisms in their intestines and faeces even after 90 days post-administration. However, although the present study did not examine early bacterial discharge in faeces, the finding that C. kutscheri was excreted in the animals' faeces for several months was consistent with these previous findings.
Even though faeces were stored for five days, viable C. kutscheri was present and could have become a source of infection for other animals. Therefore, faeces containing C. kutscheri must be managed with caution, even if they have been stored for a few days. Conversely, the persistent viability of C. kutscheri is useful for their detection because the organism can easily be cultured even from faecal samples that are a few days old.
The number of C. kutscheri in daily faecal samples remained stable in each animal. Any variation in this stability might be an indicator of stress, as latent infection with C. kutscheri can become a clinical problem when subjected to stressors, including shipment, overcrowding, nutritional deficiencies (Zucker & Zucker 1954), immnosuppressive drugs (Fauve et al. 1964, Miyamae & Shinoda 1982), irradiation (Schechmeister & Adler 1953) and experimental manipulation. We have found that mice which developed a clinical disease after oral inoculation with C. kutscheri accompanied by cortisone treatment exhibited ulcerated caeca that were positive for the organism (106.60–109.30 CFU/g), and most of their internal organs were also positive for C. kutscheri (Amao H, unpublished observations). Many papers have discussed the relationship between stress and mucosal immunity and have concluded that the host's colonic epithelial defence function is significantly affected by stress (Schleifer et al. 1983, Saunders et al. 1994, Kiliaan et al. 1998, Alverdy & Rocha 1999, Santos et al. 1999, Meddings & Swain 2000, Hart & Kamm 2002). In contrast, studying the immunological state of a host mouse with a small number of C. kutscheri in its faeces may help to identify factors involved in the organism/host balance. The number of C. kutscheri in faecal samples may become a measure of the host's condition, but further controlled studies are required. Furthermore, colonization resistance (Vollaard & Clasener 1994) may influence the bacterial population in the intestine, thereby affecting the C. kutscheri population in the faeces.
C. kutscheri was excreted in the faeces of infected mice for several months. Direct impression cultivation of faecal pellets seems to be a simple screening method for detecting C. kutscheri infection. However, the organism was not detected in direct impression cultures of faeces from two mice infected with low numbers of the organism (below 103.80 CFU/g). We recommend cultures of homogenized faecal pellets in HIB in combination with cultures of oral swabs for the detection of C. kutscheri based on the results of the present study and our previous study (Amao et al. 2002), in which oral cavity swabs from rats were cultured on FNC agar and were found to be useful for the detection of subclinical infections with C. kutscheri.
We have also found differences in susceptibility to infection with C. kutscheri among sexes and various mouse strains (Amao et al. 1993, Komukai et al. 1999). These differences may be important when screening for infection with C. kutscheri. Future investigation is necessary.
In summary, our present study showed that subclinically infected mice excrete C. kutscheri in their faeces consistently for at least several months. Therefore, faecal samples would be useful for the detection of C. kutscheri in living mice in a manner that is not stressful for the animal.
References
1.
AlverdyJC, RochaF (1999) Surgical stress, bacteria and mucosal immune function. European Journal of Pediatric Surgery9, 210–13
2.
AmaoH, SaitoM, TakahashiKW, NakagawaM (1990) Selective medium for Corynebacterium kutscheri and localization of the organism in mice and rats. Jikken Dobutsu39, 519–29
3.
AmaoH, AkimotoT, TakahashiKW, NakagawaM, SaitoM (1991) Isolation of Corynebacterium kutscheri from aged Syrian hamsters (Mesocricetus auratus). Laboratory Animal Science41, 265–8
4.
AmaoH, KomukaiY, SugiyamaM, SaitoTR, TakahashiKW, SaitoM (1993) Differences in susceptibility of mice among various strains to oral infection with Corynebacterium kutscheri. Jikken Dobutsu42, 539–45
5.
AmaoH, KomukaiY, AkimotoT, (1995a) Natural and subclinical Corynebacterium kutscheri infection in rats. Laboratory Animal Science45, 11–14
6.
AmaoH, KomukaiY, SugiyamaM, TakahashiKW, SawadaT, SaitoM (1995b) Natural habitats of Corynebacterium kutscheri in subclinically infected ICGN and DBA/2 strains of mice. Laboratory Animal Science45, 6–10
7.
AmaoH, AkimotoT, KomukaiY, SawadaT, SaitoM, TakahashiKW (2002) Detection of Corynebacterium kutscheri from the oral cavity of rats. Experimental Animals51, 99–102
8.
FauveRM, Pierce-ChaseCH, DubosR (1964) Corynebacterial pseudotuberculosis in mice. II. Activation of natural and experimental latent infections. Journal of Experimental Medicine120, 283–304
9.
FoxJG, NiemiSM, AckermanJ, MurphyJC (1987) Comparison of methods to diagnose an epizootic of Corynebacterium kutscheri pneumonia in rats. Laboratory Animal Science37, 72–5
10.
HartA, KammMA (2002) Review article: mechanisms of initiation and perpetuation of gut inflammation by stress. Alimentary Pharmacology Therapeutics16, 2017–28
11.
Japanese Association for Laboratory Animal Science (1987) Guidelines for animal experimentation. Experimental Animals36, 285–8
12.
KiliaanAJ, SaundersPR, BijlsmaPB, (1998) Stress stimulates transepithelial macromolecular uptake in rat jejunum. American Journal of Physiology275, G1037–44
13.
KomukaiY, AmaoH, GotoN, (1999) Sex differences in susceptibility of ICR mice to oral infection with Corynebacterium kutscheri. Experimental Animals48, 37–42
14.
MeddingsJB, SwainMG (2000) Environmental stress-induced gastrointestinal permeability is mediated by endogenous glucocorticoids in the rat. Gastroenterology119, 1019–28
15.
MiyamaeT, ShinodaM (1982) Corynebacterium kutscheri invasiveness of the gastrointestinal tract in young mice. Experimental Animals31, 189–94
16.
SantosJ, SaundersPR, HanssenNP, (1999) Corticotrophin-releasing hormone mimics stress-induced colonic epithelial pathophysiology in the rat. American Journal of Physiology277, G391–9
17.
SaundersPR, KoseckaU, McKayDM, PerdueMH (1994) Acute stressors stimulate ion secretion and increase epithelial permeability in rat intestine. American Journal of Physiology267, G794–9
18.
SchechmeisterIL, AdlerFL (1953) Activation of pseudotuberculosis in mice exposed to sublethal total body radiation. Journal of Infectious Diseases92, 228–39
19.
SchleiferSJ, KellerSE, CamerinoM, ThorntonJC, SteinM (1983) Suppression of lymphocyte stimulation following bereavement. Journal of the American Medical Association250, 374–7
20.
VollaardEJ, ClasenerHAL (1994) Colonization resistance. Antimicrobial Agents and Chemotherapy38, 409–14
21.
YokoiyamaS, AndoT, HayanoK, FujiwaraK (1975) Latent infection with Corynebacterium kutscheri in mice after peroral inoculation and its provocation by cortisone. Experimental Animals24, 103–10
22.
ZuckerTF, ZuckerLM (1954) Pantothenic acid deficiency and loss of natural resistance to a bacterial infection in the rats. Proceedings of the Society for Experimental Biology and Medicine85, 517–21