
Abstract

Introduction
The concept of epigenetics was pioneered in 1939 by the Edinburgh-based developmental biologist, Conrad H Waddington 1 to encompass all aspects of embryological development, but today the term is more specifically applied to the study of hereditable change in the expression of genes without change in DNA sequences. In effect this amounts to a range of non-DNA-based mechanisms that control gene transcription. 2 These epigenetic mechanisms may be influenced by environmental factors, and this has potential therapeutic implications. When epigenetic mechanisms are inherited, this has evolutionary significance. 3, 4 Genomic duplication, particularly the whole genomic duplications that result from hybridogenesis and polyploidy, also has evolutionary and medical implications. In this final review, we shall examine the evolutionary implications of epigenetics and genomic duplications for human evolution, and the potential such understanding offers to medicine.
Epigenetics
Astrocytes, renal tubular cells and macrophages look and behave very differently from each other, yet they all contain the same genetic information. These differences in cellular and tissue differentiation are brought about by systems of epigenetic control rather than through genetic differences between cells. We are familiar with how genes, made up of DNA, are intrinsically bound up within RNA, proteins and other molecules, to form a tightly coiled package known as chromatin. The chromatin may be chemically marked, and this mark will help determine whether or not the gene is available to be translated to protein, or to fulfil a developmental or regulatory function. A common way of doing this is to attach a methyl group to the nucleotide molecule, cytosine, in the promoter sequence of a gene. Extensive methylation of cytosine in the promoter will switch off the promoted gene during development or post-developmental life. Cytosine methylation is also a powerful mechanism for suppressing unwanted viral genes and sequences within the genome. Other epigenetic mechanisms include a variety of covalent chemical modifications of ‘histone tails’ (regions of the histone protein that protrude from the chromosomes and are thus made available for chemical interaction with methyl, acetyl and other chemical groups), and the role of ‘regulatory RNAs’, for example through RNA interference, or RNAi, which targets specific messenger RNAs, preventing their translation to proteins. Two additional, and clinically important, mechanisms are ‘paramutation’ and ‘genomic imprinting’. Paramutation implies interaction between the maternal and paternal alleles of the same gene, so that expression of one allele is influenced by the presence of the other. This is capable of silencing one of the alleles, and the silenced state may be inherited. Genomic imprinting results in the expression of alleles according to a specific parent-of-origin determination, as if the gene somehow remembers whether it was inherited from the father or mother and only the ‘imprinted’ version from a specific parent will be expressed. If the imprinted gene is defective, this can result in disease despite the fact the gene is not a Mendelian dominant and the other parental allele is structurally normal.
It is important to grasp that, like mitochondrial inheritance, epigenetic inheritance is
independent of the classical Mendelian rules.
5
Moreover, multiple epigenetic mechanisms can be operational at any one time and
the different mechanisms may influence Operative mechanisms for hereditary change in evolution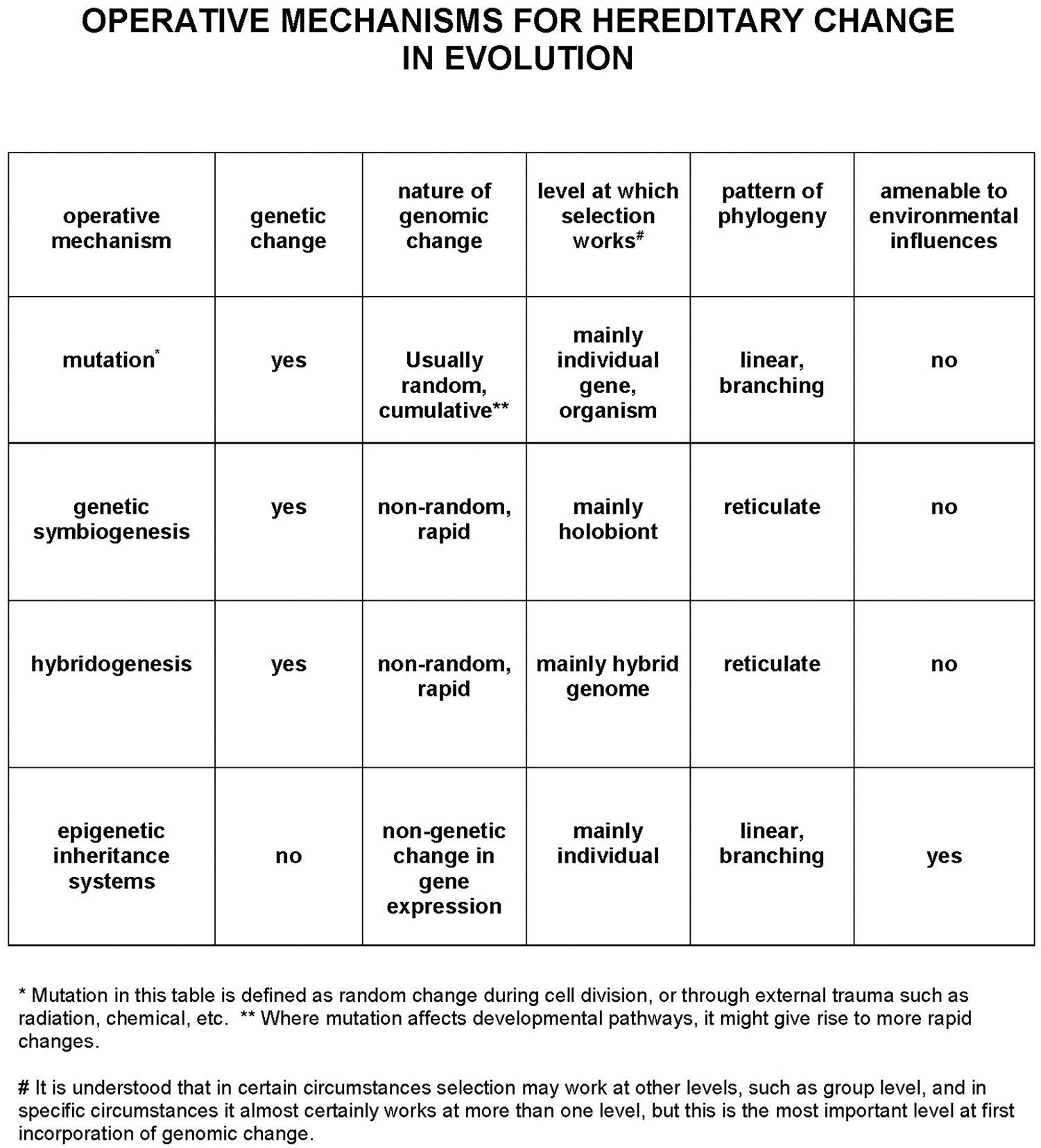
For each cell to know its fate in a predictable and reliable fashion, the epigenetic mechanisms must be remembered by the cell when it divides, so that the daughter cell follows the same histological patterns of the parent. Within the individual cell the epigenetic patterns, or epigenotype, which decide which genes or genetic control sequences are available for transcription or activation, will have dramatic effects on the cellular chemistry. A key implication is the fact that epigenetic control mechanisms are capable of change during the lifetime of the cell – indeed they have evolved the capability of being affected by environmental factors. For example, starvation during embryogenesis, or prolonged inflammation affecting an organ or tissue, and even lifestyle factors, perhaps in association with genetic predisposition, can change the epigenotype of cells, and this in turn can lead to age-related human diseases including neoplasia. 6 An earlier generation of evolutionary geneticists had assumed that acquired epigenetic change could not be inherited across generations. However, as long ago as 1975, the pioneering ideas of Holliday and Pugh, 7 and Riggs 8 suggested ways in which this might happen, and today there is growing evidence for the transmission of acquired epigenetic information across generations through a variety of mechanisms that Jablonka and Lamb have termed ‘epigenetic inheritance systems’, or EIS. 9, 10 Although understanding of such mechanisms is still limited, it means that, conceptually and dynamically, EIS constitute a fourth evolutionary mechanism for hereditary change (Figure 1). A new generation of scientists has begun to explore the applications of epigenetics to evolution and development, meanwhile the extrapolation of epigenetics to medicine is creating a new field of applied research.
Epigenetics in hereditary disease
In 2002 Cohen and colleagues reported the diagnosis of schizophrenia in a 12-year-old
boy who also suffered from a receptive language disorder.
11
On genetic analysis, the child had a point mutation in the X-chromosome-based
gene MECP2, which codes for the protein MeCP2. MeCP2 plays an
important role in two major epigenetic mechanisms of gene regulation, the methylation
of cytosine within cytosine-guanine couplets, and histone deacetylation, which alters
the structure of chromatin. Disturbance in this same gene, through a variety of
mechanisms, has also been linked to Rett syndrome, an X-linked disorder limited to
girls that is usually included within the umbrella of autism. The The epigenetic steps in carcinogenesis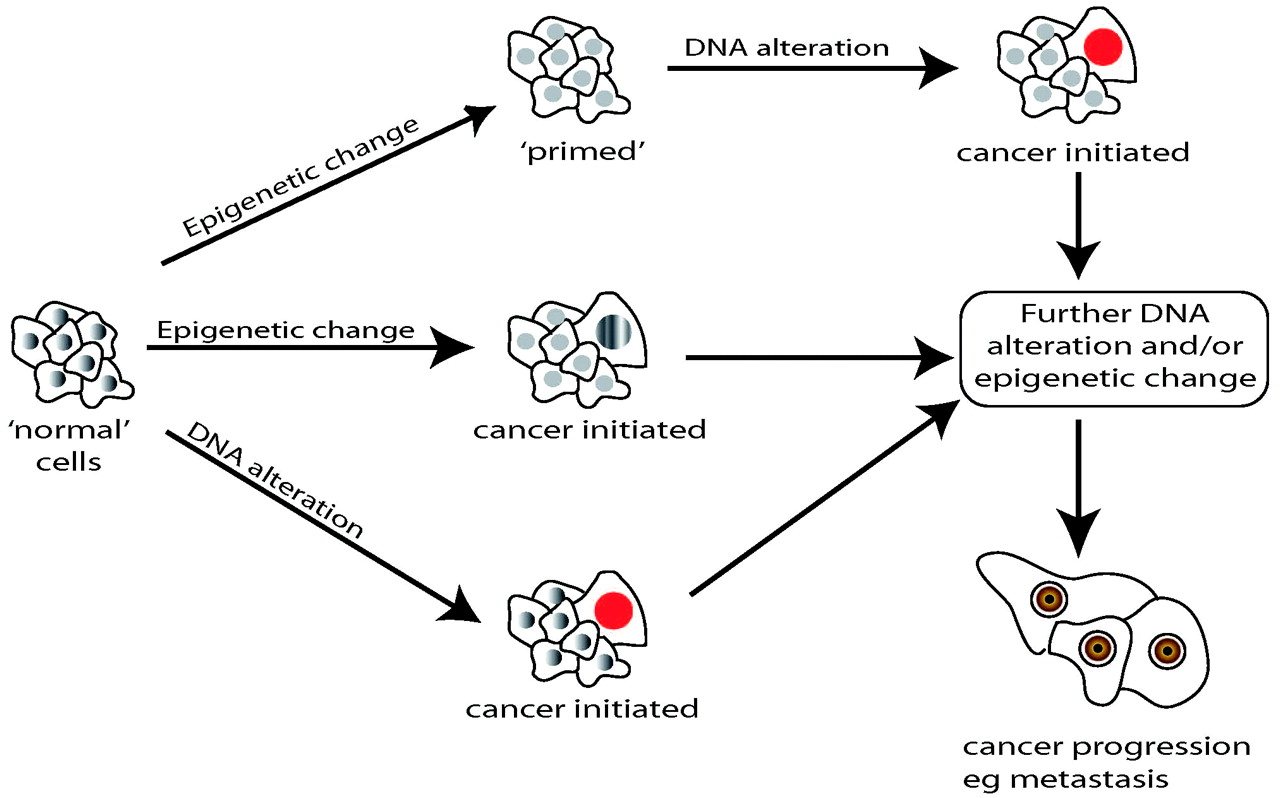
Global imprinting errors can result in more bizarre developmental anomalies, such as embryos that express a single parental genome, as seen in ovarian teratomas and hydatidiform moles. More specific genomic imprinting errors may also play a role in some cases of eclampsia, a review of which has been published by Rodenhiser and Mann. 2 The same authors highlight the fact that epigenetic disorders are significantly increased in embryos that have been manipulated during assisted reproduction and these may also be responsible for most embryonic losses in animal cloning experiments.
Epigenetics in cancer
As long ago as 1979 Holliday suggested that methylation defects might cause cancer, 19 and this was subsequently confirmed by Feinberg and Vogelstein. 20 Since then a great many studies have probed the association between epigenetic disturbance and carcinogenesis. For example, the disordered genomic imprinting associated with Beckwith Wiedemann syndrome also leads to a thousand-fold increase in the risk of childhood tumours, such as Wilm's tumour of the kidneys. 21 Loss of normal imprinting can lead to activation of oncogenes or inactivation of tumour suppressor genes, such as IGF2, and p5 kip2. 22, 23 Figure 2 illustrates the general principles of how epigenetic alterations, coupled with predisposing genetic change, may play a variety of roles in carcinogenesis.
Over the last decade there has been an expansion of epigenetic studies in cancers of stomach, colon and rectum, 24, 25, 26, 27 lymphoid and haematological malignancy, 28 breast, oesophagus and lung cancers associated with inflammation, 29 and cancer of the prostate, 30 in which the epigenetic changes have been spotted throughout all stages of tumour development, including the early stages. 31, 32 Findings such as these suggest that epigenetics may be helpful in diagnostic screening for cancer, in early diagnosis, and chemo prevention of carcinogenesis, 33 in addition to a growing potential for understanding and treatment of all varieties of malignancy. Such research will ultimately depend on improved mass screening analysis, which will need to integrate epigenetic dysregulation and the more classical genetic aspects of disease. 34, 35, 36
Epigenetic therapy of cancer
Although several epigenetic-based therapies are already available, this approach is still rapidly evolving. 37 Many therapies aim to reactivate genetically silenced tumour-suppressor genes, for example the nucleoside analogue 5-aza-2'-deoxycytidine, or ‘decitabine’, which while initially proving toxic in high doses, has shown useful therapeutic application in elderly patients with high-risk myelodyspastic syndrome. 38 Small molecule non-nucleoside analogues are undergoing early trials. Other approaches include histone deacetylase inhibitors, which comprise a large, diverse class of drugs that fall into four classes (short-chain fatty acids, hydroxamic acids, benzamides and cyclic tetrapeptides), many of which have already found their way into therapeutic use for a variety of conditions. They include sodium valproate, established in the treatment of epilepsy and mood stabilization, which has proved useful as an adjuvant therapy in myeloid leukaemia, and sodium butyrate, which induces breast cancer cell apoptosis. 39 A number of hydroxamates have entered early clinical trials and show promise in cancer therapy. These include trichlorostatin A and suberoylanilide hydroxamic acid, meanwhile newer compounds are also undergoing Phase I cancer trials. Two benzamindes are also undergoing extensive trials in cancer therapy as are several cyclic tetrapeptides. Anti-methylation treatments are somewhat limited by their lack of target specificity, so that many of the therapies tend to be used as adjuvants to more specific therapy. Nevertheless, the understanding derived from the different mechanisms of epigenetics is likely to increase the range of therapies, particularly for cancers, and newer approaches, for example the targeting of paramutations, and the growing understanding of the importance of zinc-finger proteins, may allow targeting of specific DNA sequences, and thus a more precisely honed efficacy. 40
Epigenetics in non-cancerous conditions
Given the universal nature of epigenetic regulation and the responsiveness of such regulation to environmental stimuli, it is inevitable that epigenetic factors will play an important role in a wide range of diseases. 41 Folic acid plays an important role in DNA methylation, so that dietary deficiency in early pregnancy results in epigenetic hypomethylation of the fetus, which plays an important role in the associated risk of neural tube defects. 42 Disturbed methylation has been linked with many of the so-called ‘complex disease’ predispositions,43 including asthma, diabetes and ageing, 44, 45, 46 and the latter may predispose the elderly to cancer and neurological disorders, such as neurodegenerative disorders, and adult onset autism and schizophrenia. 47, 48 Some of these epigenetic influences may be influential in the long-observed gender differences in susceptibility, which in turn may be mediated by sex hormone-induced differences in epigenetic regulation. 49 The importance of environmental influences has been confirmed by Fraga and colleagues in studies of the accumulation of epigenetic differences between monozygotic twins over their lifetimes, 50 work that also confirms the association with ageing. 51 This supports the possibility, first pioneered by Holliday, that epigenetic research may afford new insights into the process of ageing per se, with the potential of therapeutic applications. 52
Epigenetics in inflammatory conditions
Epigenetic dysregulation is being increasingly linked to the autoimmune diseases, 53 which often show discordance in expression among identical twins. 54, 55, 56 This has encouraged new lines of investigation in common inflammatory diseases, such as rheumatoid arthritis 57, 58 and SLE, 59, 60, 61 that are likely to result in novel therapeutic strategies. A single illustration may help to emphasize the potential importance of specific epigenetic changes within a cell. Bone marrow-derived antigen-presenting cells (APCs) play an important role in the initiation of T-cell responses and in the induction of T-cell tolerance. In a recent study, Villagra and colleagues, showed that a single histone deacetylase, HDAC11, negatively regulated the production of the cytokine interleukin 10 (IL-10) by APCs. 62 In effect, where there was over-expression of HDAC11, it inhibited IL-10 production and this primed naïve T cells and restored the responsiveness of tolerant CD4+ T cells, meanwhile where there was under production of HDAC11, this led to increased expression of IL-10 and impairment of antigen-specific T-cell responses. In effect, a single histone pathway was seen to regulate immune tolerance, with potential implications for autoimmune disorders in general. In the words of Wilson, which echo those of other experts, ‘the importance of the epigenome in the pathogenesis of common human diseases is likely to be as significant as that of traditional genetic mutations’. 41 Epigenetics is thus seen to be a relatively new and exciting discipline with major extrapolations to medicine.
Hybridogenesis, polyploidy and genetic duplications
The sperm and ovum are haploid, with a single set of 23 chromosomes, otherwise all normal human cells are diploid, with 46 chromosomes. A cell or organism with additional sets of chromosomes is said to be polyploid. Polyploidy arises naturally through two different mechanisms. 63, 64 A mutational dysfunction of the first reduction division of meiosis will give rise to autotetraploidy in which the duplicated sets of chromosomes are identical. Hybridisation, or sexual crossing between different, usually closely-related, species can give rise to a novel homoploid genome – one in which the number of chromosomes is normal but the genetic content is the sum of haploid sets of chromosomes from each of the parental species – or to a tetraploid genome, made up of two dissimilar sets of chromosomes. 65, 66 Where autotetraploidy creates redundant genes that may evolve to new genes, hybridogenesis, whether resulting in homoploidy or allotetraploidy, gives rise to an immediate increase in genetic complexity. 67, 68 Thus hybridisation is capable of giving rise to rapid hereditary change, and evolutionary novelty, which follows a reticulate rather than a linear pattern. 69, 70 It is also important to grasp that polyploidy can give rise to genetic instability that can prove fatal, and a newly polyploid genome needs to be quickly stabilised through the rapid mobilization of complex genetic, epigenetic and endogenous retroviral mechanisms. 71, 72, 73, 74
The genetic evidence suggests that humans have inherited two rounds of whole genomic duplications from distant ancestors, so that where the early chordates, such as Amphioxus, have only a single copy of key developmental sequences, such as the Hox cluster, we have four. 75, 76, 77 This is known as the one-to-four rule and it has played an important role in the evolution of the human genome, with key effects on body patterning during embryological development. These two rounds of duplication could have arisen through autotetraploidy or allotetraploidy, but the fact that the two episodes of tetraploidy arose in fish-like ancestors, coupled with the fact that fish readily hybridise with resultant tetraploidy, suggests that hybridisation is the more likely explanation of our tetraploid ancestry. Polyploidy is also a common finding in cells undergoing a variety of stresses, including oxidative damage, wound healing, viral infection and the processes of ageing. In fact, it is a key stage in the development of a variety of pathological conditions, particularly cancers, where it can arise through cell fusion, a developmental programme known as endoreplication, or through defects arising in abortive cell cycles. 78 The consequent genetic instability may provide a route to aneuploidy and thereby contribute to the process of carcinogenesis. 79, 80
Segmental genomic duplication, whether arising through hybridisation, mutation or recombination of HERVs, LINEs and Alus, is common in the human genome, 81, 82 and it has played an important part in the evolution of zinc-finger, olfactory-receptor, certain globin, opsin, coagulation and fibrinolytic genes. 83 Duplication has been linked to some cases of Angelman syndrome, where the partial deletion of chromosome 15 appears to result from an overly duplicated segment of the chromosome, and duplication is also one of the genetic causes of Charcot-Marie-Tooth syndrome, which was shown by Lupski et al. to result from an additional copy of the gene encoding the protein, PMP, which is a major component of myelin. 84 Another duplication, on chromosome 15, has been linked to susceptibility to panic and phobic disorders, with a curious non-Mendelian inheritance that is also linked to joint laxity. 85 Detailed genetic analysis of various chromosomes is revealing widespread deletions and duplication of segments, which appear to be primate-associated, 81, 86, 87, 88 and are of growing importance to our understanding of the genetic predisposition to disease, including autism, survival in breast cancer and Cowden's disease. 89, 90, 91
Conclusion
It is clear that the evolutionary mechanisms that give rise to hereditary genetic and genomic change are the same mechanisms that give rise to the genetic basis of human disease. Thus, recent advances in our understanding of evolutionary biology have important implications for medicine. Rather than group different mechanisms under the former catch-all of ‘mutation’, we should consider a spectrum of clearly-defined mechanisms, with mutation restored to its original and mechanistically precise and useful definition. This offers a more comprehensive understanding of human evolution, development and physiology as well as possible new avenues of research relevant to a wide diversity of disease. Of particular note is the repeated observation of how each of the driving mechanisms for hereditary genetic change have commonalities with the pathogenetic mechanisms of cancer. This is in keeping with the proposals from various authors and fields that carcinogenesis might be seen as an evolution of the cell towards independence, involving all mechanisms of hereditary change and with selection switching from cooperation to selfish independence. 92, 93, 94, 95, 96
Footnotes
DECLARATIONS