
Abstract
Background
Triglyceride-rich low-density lipoproteins (TG-rich LDLs) in the plasma of patients with severe liver disease are reported to change macrophages into foam cells in vitro.
Methods
Male BALB/c mice were immunized with TG-rich LDLs isolated from the plasma of a patient with severe liver disease. The resulting monoclonal antibody (G11-6) was used in a sandwich enzyme-linked immunosorbent assay (ELISA) in combination with polyclonal anti-apolipoprotein B antibodies. The time course of copper-mediated LDL oxidation was monitored using this ELISA. The results were compared with those of the two commercial ELISAs for oxidized LDLs using DLH or ML25, thiobarbituric acid reactive substances and the optical absorbance for the conjugated dienes generated in lipid peroxides. Furthermore, the lipoprotein fractions separated by gel filtration were tested with this ELISA in healthy volunteers (n = 11) and patients (n = 3) with liver disease.
Results
G11-6 reacted with oxidized LDLs during only the early phase of copper oxidation, being distinct from the other monoclonal antibodies and methods. G11-6 was confirmed to react with TG-rich LDLs in patients, while it reacted with small LDL particles in normal controls.
Conclusions
The monoclonal antibody G11-6 is useful for detecting oxidized small LDLs in normal controls and oxidized TG-rich LDLs in patients with severe liver disease.
Introduction
Oxidative modification of low-density lipoproteins (LDLs) and the subsequent conversion of macrophages into foam cells in atherosclerotic lesions play a key role in early atherogenesis. 1–3 Oxidized LDLs have been detected at elevated levels in circulating plasma from patients with coronary artery disease using enzyme-linked immunosorbent assays (ELISAs). 4–7 Small dense LDLs are more susceptible to oxidation than larger, buoyant ones. 8 Copper-mediated LDL oxidation in vitro decreases LDL particle size, which could contribute to the generation of small, dense LDL particles. 9 Oxidation of lipids and lipoproteins has been implicated in a growing number of diseases including diabetes mellitus, 10 chronic kidney disease, 11 Alzheimer's disease, 12 autoimmune disease 13 and cancer, 14 as well as in ageing. 15
In our previous study, we observed abnormal triglyceride-rich LDLs (TG-rich LDLs) in patients with cholestatic liver disease. This lipoprotein species strongly promotes the conversion of macrophages into foam cells in vitro. 16 To further investigate its pathophysiological role, we developed a new monoclonal antibody against TG-rich LDLs. Its biochemical properties and reactivity with lipoproteins are described in the present report.
Materials and methods
Immunogen
TG-rich LDLs were used as immunogens to develop a new monoclonal antibody, designated as G11-6. Briefly, blood was drawn from a patient with severe cholestasis due to advanced primary biliary cirrhosis, who had provided informed consent. The change in electrophoretic mobility of serum samples was confirmed by agarose gel electrophoresis. 17 TG-rich LDLs were isolated from the serum sample by ultracentrifugation and gel chromatography, as previously reported. 17,18 The chemical composition of this lipoprotein fraction was determined. Lipids were measured by automated enzymatic methods using the following commercial kits: Cholestest CHO for total cholesterol (TC), Cholestest TG for triglycerides (TG), Pureauto S PL for phospholipids (PL), Qualigent LDL for LDL-cholesterol and Qualigent HDL for high-density lipoprotein (HDL)-cholesterol (Sekisui Medical Co, Ltd, Tokyo, Japan), and Determiner L FC for free cholesterol (FC) (Kyowa Medex Co, Ltd, Tokyo, Japan). Cholesterylester (CE) concentrations were calculated by multiplying the esterified cholesterol concentrations (obtained by subtracting FC from TC) by 1.72. Protein was determined by the method of Lowry, modified by Markwell et al. 19 Apolipoproteins were analysed by 3–10% sodium dodecyl sulphate polyacrylamide gel electrophoresis (SDS-PAGE) after reduction with 2-mercaptoethanol.
Immunization
Male BALB/c mice (6 weeks old) were immunized with the isolated TG-rich LDLs (0.5–1.0 g/L, 0.1 mL/injection) and were given booster injections two and six weeks later. 20 Three days after the final injection, spleen cells were collected and fused with P3/U1 mouse myeloma cells using 50% polyethylene glycol 1500 (Roche Diagnostics, Mannheim, Germany). The fused cells were cultured in Gibco RPMI 1640 medium (Invitrogen, Carlsbad, CA, USA) containing fetal bovine albumin (10%), hypoxanthine, aminopterin and thymidine in a 96-well microtiter plate.
Ten days after cell fusion, cells from antibody-containing wells were cloned by limiting dilution. The hybridoma clones were cultured in RPMI 1640 medium and injected intraperitoneally into mice primed with 0.5 mL pristine. Ascitic fluid was purified by ammonium sulphate precipitation followed by gel filtration chromatography on a Superose 6 column.
Antibody characterization
The selected antibodies were isotyped using a commercially available kit (IsoStrip Mouse Monoclonal Antibody Isotyping Kit; Roche Diagnostics) according to the manufacturer's instructions.
ELISA
The wells of a 96-well microtiter plate (Nalge Nunc International, Reskilde, Denmark) were coated with 50 μL of G11-6 solution (5 mg/L in 10 mmol/L phosphate-buffered saline [PBS, pH 7.4]), and the plate was incubated for two hours at 37°C. The wells were then blocked with 150 μL of 1% bovine serum albumin in PBS for two hours at 37°C. After four washes with PBS containing 0.05% Tween 20 (PBS/T), a 50 μL sample was added to each well, and the plate was incubated overnight at 4°C. After washing, 50 μL of biotinylated goat polyclonal antibody against human apolipoprotein B (ApoB) (10 mg/L; BW127; WatPa Enterprises, Auckland, New Zealand) was added to each well, and the plate was incubated for one hour at room temperature. After washing, 50 μL of alkaline phosphatase-conjugated streptavidin (1:250 in PBS/T; Zymed Laboratories, San Francisco, CA, USA) was added to each well, and the plate was incubated for 30 min at room temperature. After washing, 100 μL of p-nitrophenyl phosphate (1 g/L) in diethanolamine solution was added to each well and allowed to react with the enzyme for 60 min at room temperature. The absorbance at 405 nm was then measured against a reference absorbance at 620 nm using a microplate reader (Bio-Rad Laboratories, Hercules, CA, USA).
Apolipoproteins and oxidative stress markers
ApoB was measured by sandwich ELISA with the BW127 anti-ApoB antibody using the same procedures as those used in the ELISA with G11-6. Two commercial ELISA kits for oxidized LDL measurement were used. The MX kit (Kyowa Medex Co, Ltd) using the DLH monoclonal antibody detects oxidation-induced short-chain phosphatidylcholine (oxPC). 21 The MDA-LDL ELISA kit (Sekisui Medical Co, Ltd) using the ML25 monoclonal antibody detects malondialdehyde (MDA)-modified ApoB. 22 The lipoproteins detected by the MX and MDA-LDL ELISA kits were designated oxPC-LDLs and MDA-LDLs, respectively.
Thiobarbituric acid reactive substances (TBARS) were measured using a commercial colorimetric kit (Cayman Chemical Co., Ann Arbor, MI, USA), and levels of conjugated dienes were monitored by following the absorbance at 234 nm. 23
Copper-mediated oxidation of LDLs purified from serum
LDLs were purified from serum by sequential ultracentrifugation in a near-vertical tube rotor (MLN-80; Beckman Coulter, Fullerton, CA, USA) in an Optima MAX ultracentrifuge (Beckman Coulter). 18,24,25 Each serum sample (2.0 mL) was adjusted with KBr solution to a density of 1.019 kg/L and then centrifuged at 40,000 rpm for 20 h at 15°C. The upper fraction (2.5 mL) containing chylomicron, very low-density lipoprotein and intermediate-density lipoprotein was removed. The lower fraction was adjusted with KBr solution to a density of 1.063 kg/L and centrifuged at 50,000 rpm for 18 h at 15°C. The resulting LDL-containing upper fraction (2.5 mL) and the resulting lower fraction, which contained HDL and other serum proteins, were both recovered and assayed for their protein concentrations using a modified Lowry method. 19 Excellent separation of the lipoproteins was confirmed by polyacrylamide gel electrophoresis (LipoPhor; Jokoh Co, Ltd, Tokyo, Japan) (data not shown).
To prepare copper-oxidized LDLs, the purified LDL fraction was dialysed against PBS for 16 h at 4°C and diluted with PBS to a protein concentration of 0.5 g/L. Aliquots of diluted LDLs were then incubated in the presence of 3.3 μmoL/L CuSO4 at 37°C. After various lengths of time (0–8 h), the reaction mixtures were immediately analysed after dilution to the specified protein concentrations using the following assays: G11-6 ELISA (50 mg protein/L), ApoB (0.1 mg/L), oxPC-LDLs (0.2 mg/L), MDA-LDLs (0.5 mg/L), TBARS (not diluted) and conjugated diene (40 mg/L). All data are presented as a ratio to the baseline value (0 h). The change in the electrophoretic mobility of LDLs was confirmed by agarose gel electrophoresis. 17
Preparation of MDA-modified LDLs
MDA solution (0.02 mol/L) in 0.1 mol/L phosphate buffer (pH 6.4) was prepared from malondialdehyde tetrabutylammonium salt (Sigma, St Louis, MO, USA). LDLs (0.5 g/L protein) were dialysed against 10 mmol/L phosphate buffer (pH 7.4) containing 150 mmol/L NaCl and 0.268 mmol/L ethylenediaminetetraacetic acid (EDTA), and modified by mixing equal volumes of 0.02 moL/L MDA solution followed by incubation for three or 18 h at 4°C. To remove excess MDA, the mixture was dialysed against 10 mmol/L phosphate buffer (pH 7.4) containing 150 mmol/L NaCl and 0.268 mmol/L EDTA. The reaction mixtures were analysed by the following assays after dilution to the specified protein concentrations: G11-6 ELISA (50 mg protein/L), MDA-LDLs (0.5 mg/L) and TBARS (not diluted). All data are presented as ratios of the baseline value (0 h).
Gel filtration chromatography
Serum samples were fractionated by gel filtration chromatography on a Superose 6 column (GE Healthcare, Little Chalfont, UK) in a liquid chromatography apparatus (Shimadzu, Kyoto, Japan) using a procedure modified from a previous report. 26 The column was eluted with 50 mmol/L phosphate buffer (pH 7.4) containing 150 mmol/L NaCl and 1 mmol/L EDTA at a flow rate of 0.5 mL/min. Eluted fractions (0.5 mL each) were subjected to lipid analyses and immunoassays.
Serum samples
Clinical parameters and serum lipids in studied groups
*P < 0.05 versus healthy volunteers
Procedural ethics
All individuals provided written informed consent to participate in the study. The study was approved by the ethics review board of the Faculty of Health Sciences, Hokkaido University (approval number 08-57).
Statistical analysis
All clinical parameters except for sex between the groups were compared using a non-parametric Mann-Whitney U test. The influence of sex among groups was assessed by Fisher's exact probability test. Differences were considered to be statistically significant at P < 0.05.
Results
Chemical composition of the immunogen
The serum sample from the immunogen donor showed only a β-band in an agarose gel (Figure 1). This β-band migrated faster and broader than the normal examples did. The chemical composition (weight%) of the β-migrating lipoproteins isolated by ultracentrifugation and gel filtration demonstrated an abundance of LDLs with triglycerides: CE, 9.4%; FC, 13.8%; TG, 28.7%; PL, 29.1%; and proteins, 19.0% (normal: CE, 39.1%; FC, 7.2%; TG, 8.4%; PL, 22.4%; and proteins, 21.3%
16
). SDS-PAGE showed that the TG-rich LDLs contained a 550 kDa protein as the major apolipoprotein (data not shown).
Typical electrophoretic patterns of serum lipoproteins on agarose gel electrophoresis: a normal control (a) and a patient with severe liver disease (b). Lipoproteins were stained with Fat Red 7B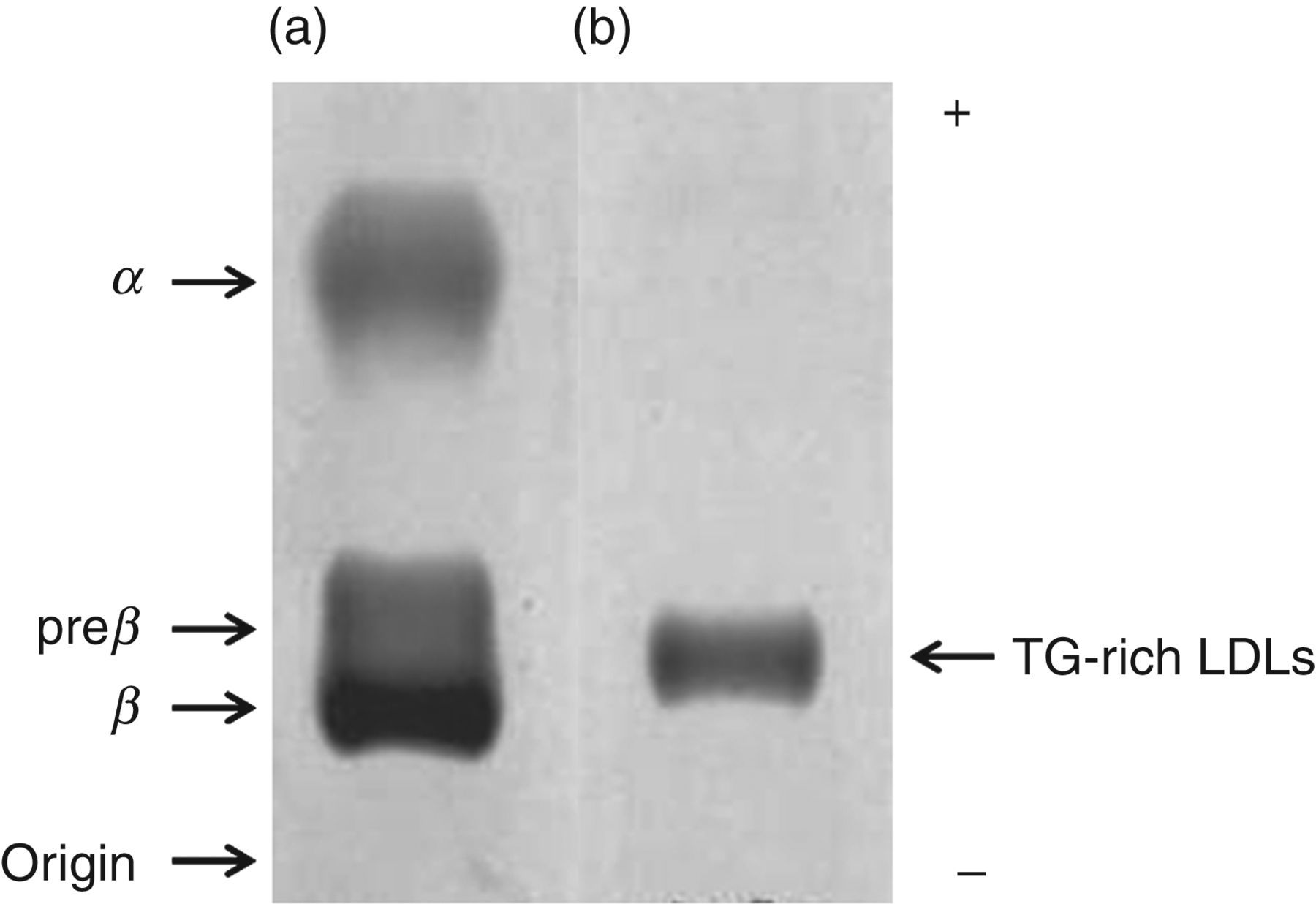
Biochemical characteristics
Hybridomas more reactive to copper-oxidized LDLs and less reactive to native LDLs were screened. Of a number of positive clones, G11-6 was selected because it showed a significant reaction with copper-oxidized LDLs. The isotype of G11-6 was IgM, with a κ light chain.
Copper-mediated oxidation of LDLs
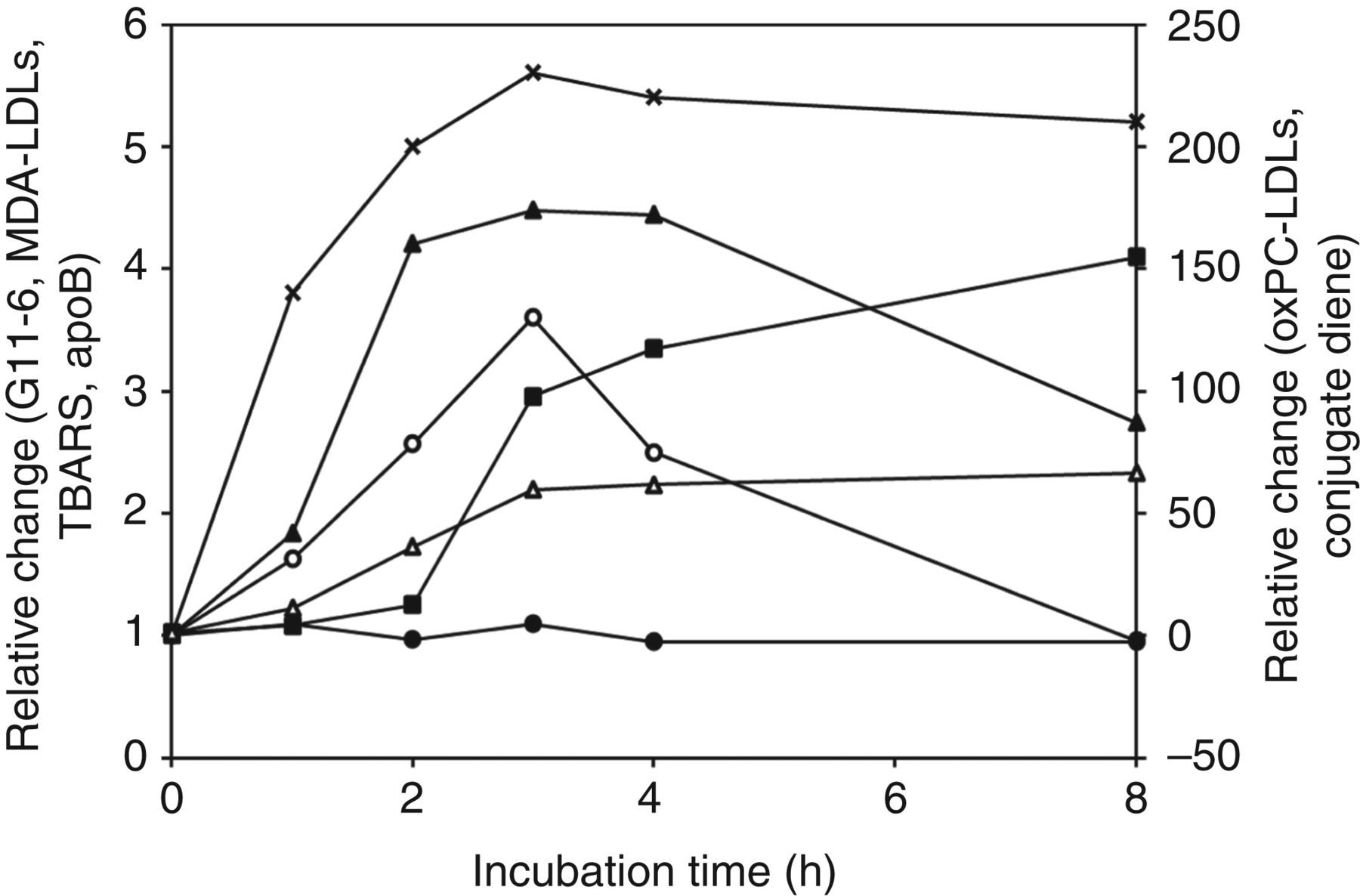
LDL oxidation was confirmed by a mobility shift in agarose gel electrophoresis (Figure 2). The reactivity of LDLs with the ELISA using G11-6 increased promptly and linearly from the beginning of copper oxidation, peaking at three hours, and then declined to a basal level at eight hours (Figure 3). TBARS also showed a prompt elevation and reached a maximum at three hours, but remained elevated at eight hours. A lag occurred in MDA-LDLs and conjugated dienes until one hour, and in oxPC-LDLs until two hours. MDA-LDLs reached a maximum at three hours and declined after four hours, but remained elevated at eight hours. The conjugated dienes and oxPC-LDLs did not decline at any time point. The ApoB ELISA remained constant for eight hours.
The change in electrophoretic mobility on agarose gel during copper oxidation of normal LDLs The time course of copper-mediated LDL oxidation as determined by ELISA using G11-6 (○, left axis), and by MDA-LDLs (▴, left axis), TBARS (×, left axis), apoB (•, left axis), oxPC-LDLs (▪, right axis) and conjugated dienes (Δ, right axis). All data are presented as the ratio of absorbance at each time point to that for the zero time. ELISA, enzyme-linked immunosorbent assay; MDA, malondialdehyde; TBARS, thiobarbituric acid-reactive substances; oxPC, oxidation-induced short-chain phosphatidylcholine; apoB, apolipoprotein B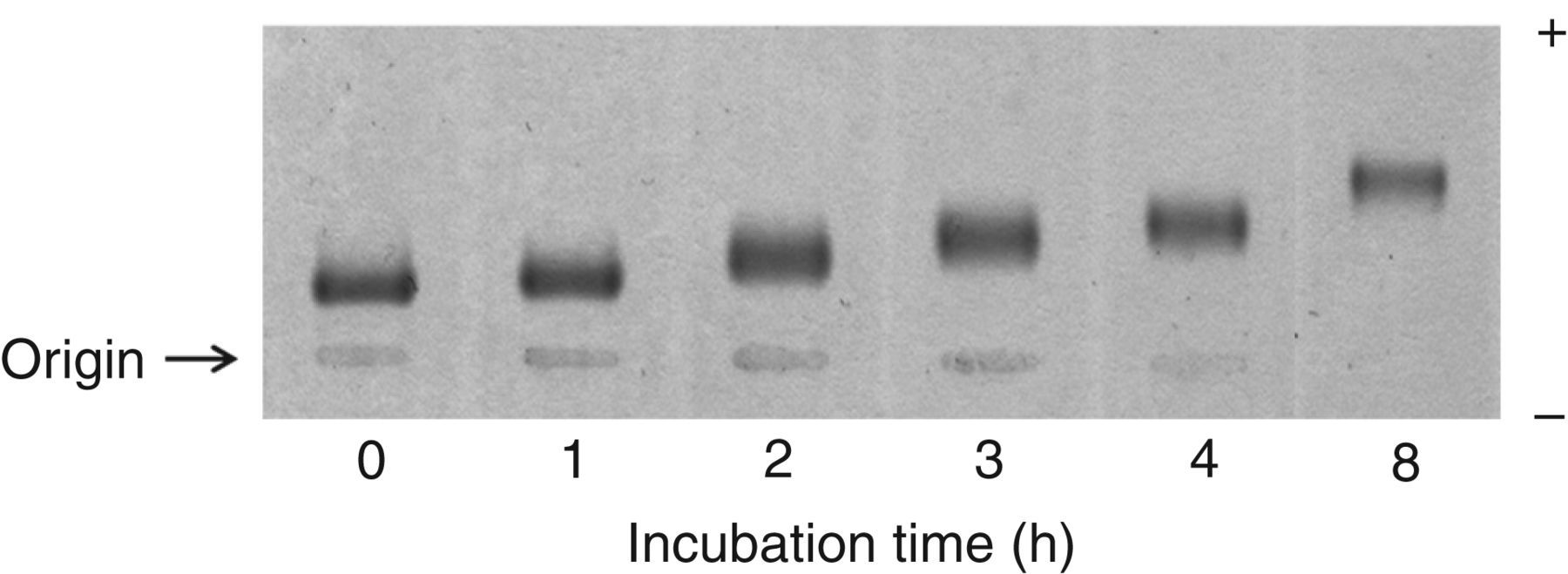
Interaction between the G11-6 and MDA-modified LDLs
The G11-6 ELISA did not react with MDA-modified LDLs, while the MDA-LDL ELISA and TBARS exhibited clear responses (Figure 4).
The reactivity of the MDA-modified LDLs with ELISA using G11-6 (○, left axis), MDA-LDL ELISA (▴, left axis) and TBARS assay (×, right axis). All data are presented as the ratio of absorbance at each time point to that for the zero time. ELISA, enzyme-linked immunosorbent assay; MDA, malondialdehyde; TBARS, thiobarbituric acid reactive substances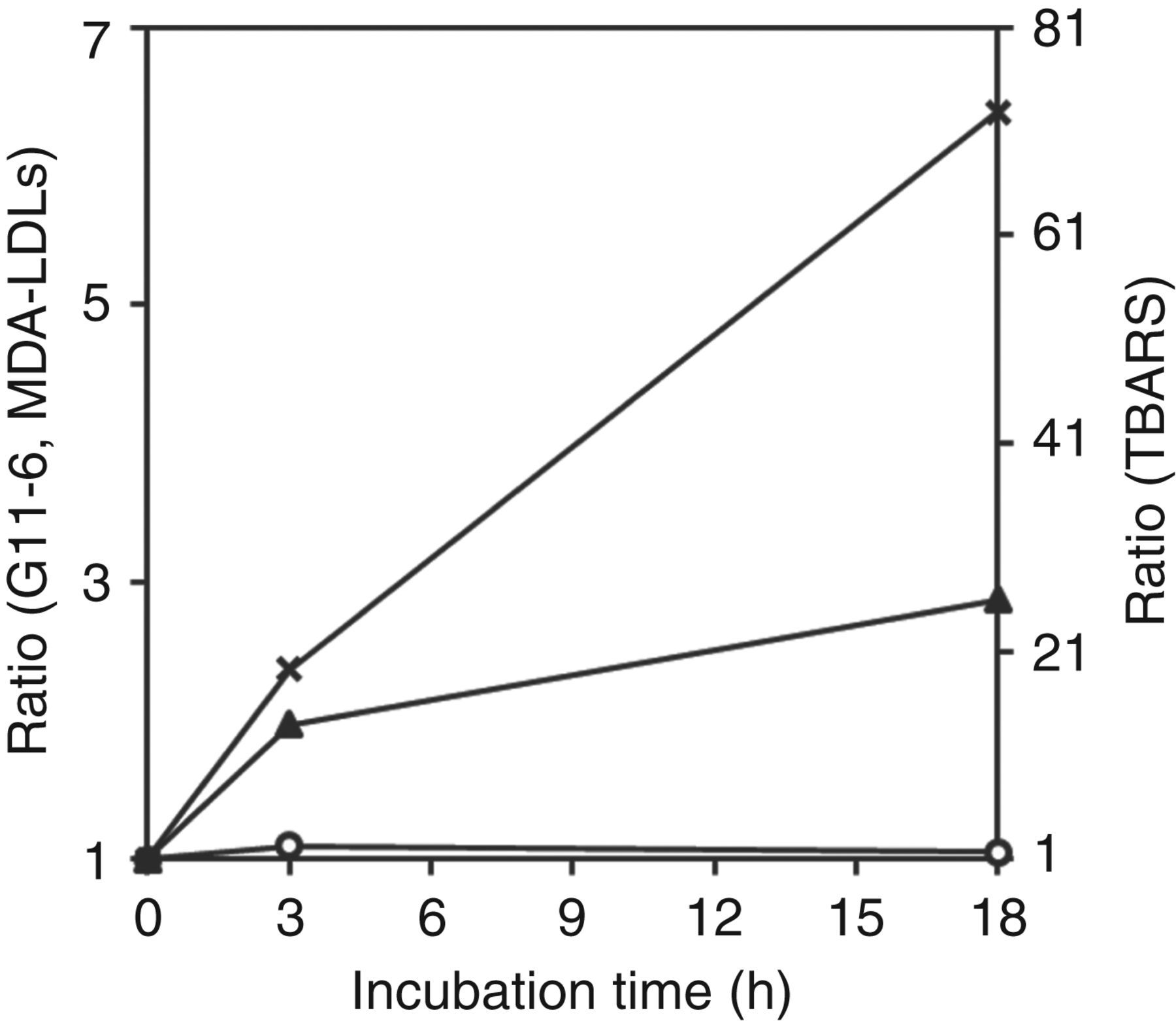
Gel filtration chromatography
The serum sample obtained from a male patient with severe liver disease (TC, 1.72 mmol/L; TG, 0.58 mmol/L, PL, 1.16 mmol/L, HDL-C, 0.10 mmol/L; and LDL-C, 0.91 mmol/L) was separated by gel filtration high-performance liquid chromatography (Figure 5A). The major lipoproteins eluted in fractions 11–15 were characterized by a high TG content, demonstrating the characteristics of the TG-rich LDLs associated with severe liver disease. The reactivity of G11-6 with the TG-rich LDLs was confirmed by the overlapping profiles. Essentially, identical elution profiles were observed in all three patients with severe liver disease examined in this study. In contrast, as shown in Figure 5B, serum from a normal male (TC, 5.69 mmol/L; TG, 1.28 mmol/L; PL, 3.07 mmol/L; HDL-C, 1.72 mmol/L; and LDL-C, 3.48 mmol/L) was separated using an identical method. Normal LDLs eluted in identical fractions as the TG-rich LDLs (Figure 5A). Lipoproteins reactive with G11-6 eluted slower than the peak of LDL-cholesterol, indicating that G11-6 reacted with small, dense LDLs. All healthy volunteers (n = 11) yielded essentially the same results.
Elution profile in gel filtration HPLC for (a) a serum sample from a representative patient with severe liver disease and (b) a serum sample from a representative healthy volunteer. Total cholesterol (•, left axis), triglycerides (▴, left axis), phospholipids (×, left axis) and the ELISA using G11-6 (○, right axis). a, void volume; b, large, buoyant LDLs; c, small dense LDLs; d, HDLs; e, TG-rich LDLs; ELISA, enzyme-linked immunosorbent assay; HPLC, high-performance liquid chormatography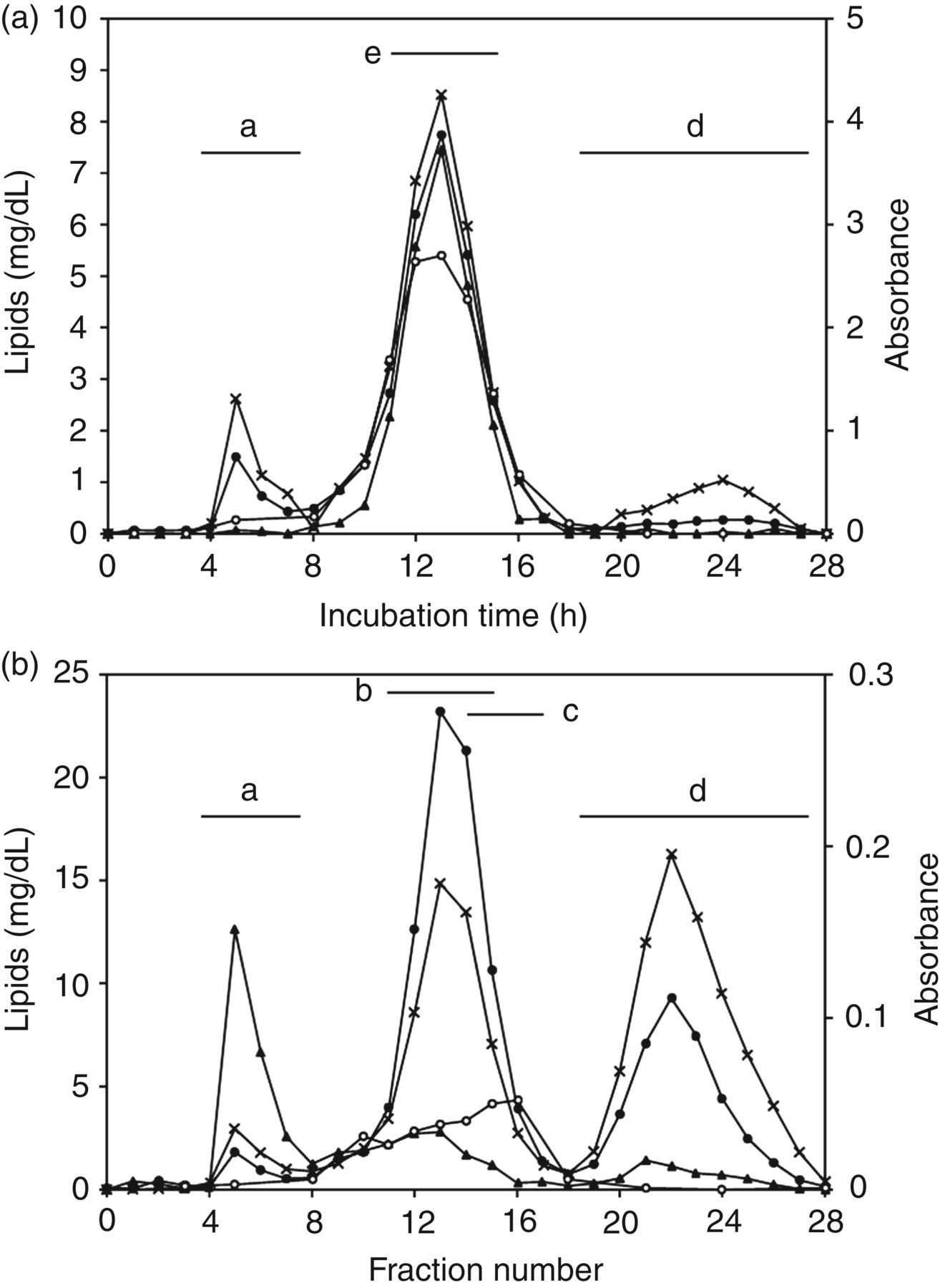
Discussion
Previous attempts to develop monoclonal antibodies against oxidized LDLs have used, as immunogens, the homogenates of human atheromatous plaques or LDLs modified in vitro, such as by MDA treatment or metal-induced oxidation. 3,22,27,28 In this study, naturally oxidized lipoproteins, TG-rich LDLs, circulating in a patient with severe liver disease were employed as the immunogen. To the best of our knowledge, G11-6 represents the only monoclonal antibody against circulating oxidized LDLs.
TG-rich LDLs used as immunogens were strikingly rich in TG content, but poor in CE, as previously reported. 29,30 The major apolipoprotein in those studies was apoB-100, which originated in the liver. Our previous study reported that serum MDA-LDL concentration was high in patients with severe liver disease and that the TG-rich LDLs isolated from plasma of a patient with severe liver disease promoted the conversion of macrophages into foam cells in vitro as efficiently as copper-oxidized LDLs. 16 Furthermore, the striking increase in the hydroperoxides of cholesterylesters and TGs in plasma from patients with liver failure has been reported. 31 Although the immunogens used in this study were not analysed for any oxidized material, it is very likely that the immunogens used here are oxidized LDLs.
It is often the case that the extent of lipoprotein oxidation is defined by the change in the electrophoretic mobility of LDLs in agarose gel electrophoresis, by the loss of recognition for LDLs by the LDL receptor, and by the strength of the oxidation-related response in cells to oxidized LDLs. 32 In this study, the relative mobility, which was measured by dividing the electromobility of oxidized LDLs by that of native LDLs, for our copper-oxidized LDLs (oxidized for 3–4 h), was similar to that for mildly modified LDLs reported by Itabe et al. 33 As shown in Figure 3, G11-6 did not recognize heavily oxidized LDLs, but did detect oxidized lipoproteins during the early process of oxidation. The rapid decline in the reactivity of oxidized LDLs with G11-6 after three hours was not due to the deterioration of ApoB-100, because the reactivity of the ApoB ELISA was constant.
This delayed and prolonged time course for oxPC-LDLs detection clearly demonstrates that the DLH antibody recognizes more oxidized LDLs than do G11-6 or ML25. The G11-6 epitope remains unknown, but is different than that for ML25, as evidenced by the diverse time courses during copper oxidation (Figure 3), and by the lack of G11-6 reactivity with MDA-modified LDLs (Figure 4). In addition, ML25 requires sodium dodecyl sulphate in the reaction buffer to expose the epitope, while G11-6 does not.
It is of interest that DLH and ML25 both exhibited a lag time, but G11-6 did not (Figure 3). DLH and ML25 recognize short acyl-chained phosphatidylcholines and MDA-modified ApoB, respectively. Both structures are thought to be produced during lipid oxidation later than lipid hydroperoxide formation. 34 The identity of the G11-6 epitope is a focus of research in our laboratory.
Our gel filtration study confirmed that G11-6 reacts with TG-rich LDLs in the blood of patients with severe liver disease (Figure 5A), which was expected, because TG-rich LDLs were used to raise the G11-6. It is noteworthy that G11-6 can bind to small, dense LDLs in normolipidemic sera (Figure 5B), indicating that identical or very similar structures relevant to oxidation exist on the surface of both lipoproteins. Accordingly, small, dense LDLs have been reported to be oxidized, 22 or more easily oxidizable than large, buoyant ones. 8 Hence, our observations provide support for the role of small, dense LDLs in atherosclerosis.
The biological effect of LDL oxidation is reported to be diverse and depends on oxidation level. 32 Mildly oxidized LDLs induce overall cell death, such as necrosis and apoptosis, more than moderately oxidized LDLs in human monocyte/macrophage systems, where the former are defined as containing more lipid hydroperoxides, but less oxysterol, MDA and negative charge than the latter. 35,36 Further, mildly oxidized LDLs are reported to exhibit cytotoxic and proinflammatory activities. 37 In this context, the oxidized lipoproteins reacting with G11-6 may be more important for clinical and research purposes than heavily oxidized LDLs.
In terms of liver disease, G11-6 might serve as a tool for evaluating oxidation and inflammation in the liver. 38 Recent studies have suggested that antioxidant therapy improves liver function in patients with chronic liver disease, such as chronic hepatitis C and non-alcoholic steatohepatitis. 37,39 The potential utility of G11-6 in such antioxidant therapies for liver disease should be tested. We are now preparing for a large-scale clinical study for this purpose.
The generation mechanism for the TG-rich LDLs in liver disease also remains to be addressed in the future. It has been reported that the MDR2 transporter, which is involved in toxicant and drug metabolism, is involved in lipoprotein X secretion. 40 The interactions between lipoprotein metabolism with such detoxification systems remain to be explored. G11-6 may prove useful for such studies.
DECLARATIONS