Epigenetic memory refers to the inheritance of epigenetic marks to the DNA or chromatin-based changes such as DNA methylation and histone modifications.1 The methyltransferase DNMT1 transfers the existing methylation pattern during DNA replication which ensures inheritance or memory. Histone modifications like the addition of an acetyl group lead to an open chromatin. Epigenetic marks can be dynamically regulated in response to environmental cues including diet, stress, exposure to toxins, developmental signals, and cellular stressors.2 Exposures during fetal development and early childhood can determine the epigenetic landscape of an individual. Physical, chemical, or even psychological changes can contribute to a genetic alteration without changing the sequence of the genome. Eventually, these changes can be passed down through generations, as epigenetic memory potentially leads to significant health outcomes. Epigenetic memory is primarily of three types, cellular memory, transcriptional memory, and transgenerational memory.3Cellular epigenetic memory mostly refers to chromatin structure and gene expression that induce cellular differentiation which are maintained through cell divisions. The repressive histone marks during mitosis are transmitted during DNA replication and are also maintained through cell divisions.4 A proposed “Epigenetic priming” model of carcinogenesis states that the tumor origin may not only be driven by an oncogenic hit.5 The lack of TET2 or DNMT3A has been demonstrated to induce hematopoietic stem cell malignancies.6, 7 An environmental trigger or a genetic predisposition in combination with epigenetic priming was found to be equivalent to an oncogenic hit.
Transcriptional epigenetic memory refers to the retention of gene transcription patterns occurring even after several nuclear alterations with expected differential gene transcription. During each replication process, the epigenetic marks need to be identically re-established to maintain a cellular clonal differentiation yet heterogeneity is a common phenomenon in cancer. Recently, transcriptional and epigenetic identity in cancer cells was studied using RNA expression and DNA methylation analysis using single-cell sequencing.8 The study reports clonally stable variation in gene expression of epithelial origin and a reciprocal variation in EMT gene expression. The single cells and clonal methylation analysis showed actively dividing cancer cells with a high degree of epigenetic persistence although being targeted by epigenetic modulators or other trans-factors.2 Certain epigenetic remarks at the organism level are transferred more precautious to the next generation and are known as transgenerational memory. Epigenomic regulators like DNA methylation, histone moieties, and non-coding RNAs remain the known mechanisms by which they are transmitted across generations.9 Both paternal and maternal exposure during the pre-conception window and pregnancy respectively contribute to any transgenerational epigenetic memory. Animal model studies demonstrated that environmental exposure to stress and nutritional status leads to transgenerational inheritance, where a cancer-predisposed condition is transferred to subsequent generations. Maternal exposure to a fat-rich diet during gestation resulted in an increased breast cancer predisposition in F1, F2, and even in F3 generations.10 Also, paternal obesity and malnutrition before conception were associated with the progression of breast cancer in offspring.11 The molecular basis of paternal stress can be attributed to epigenetic alterations in sperm cells. Several non-coding RNAs like miRNAs and tRNAs play a significant role in inducing differential gene expression in a few early cell divisions of embryonic development.12, 13 Epigenetic inheritance through germline inheritance is frequently reported in familial cancer syndromes.
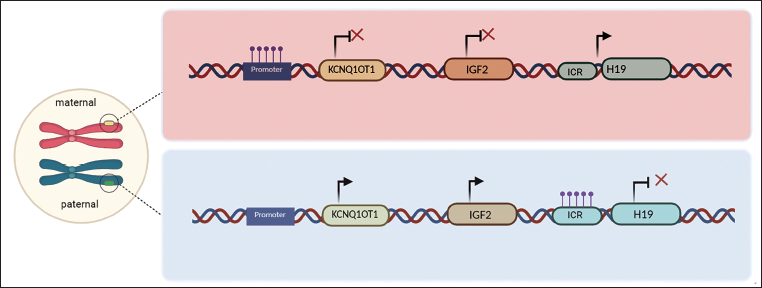
Genomic imprinting also plays an important role in determining the transgenerational epigenetic memory. Epigenetic modifications in IGF2, lncRNA KCNQ1OT1 were found associated with Beckwith–Wiedemann syndrome (BWS).14, 15 Loss of imprinting (LOI) in the IGF2 and H19 locus is associated with various cancer types. In Wilms’ tumor, IGF2 LOI leads to a maternal allele expression which is observed with increased methylation and reduced expression from H19 maternal copy.16 The different modes of epigenetic memory and their persistence affect and promote cancer initiation and progression. A comprehensive analysis of epigenetic memory is practically challenging to study the transfer of information as it involves animal models across many generations. However, both cellular memory and transcriptional memory of epigenetic factors are postulated and accumulated with evidence to have a significant role in cancer development. A multi-platform insight through spatial gene expression, single-cell RNA sequencing, and MNase-seq can illustrate the maintenance of epigenetic signature and a cohesive understanding of its precise role in tumorigenesis.
Epigenetic memory based on genomic imprinting
Cells carrying the maternal chromosome have specific methylation memory at promoter of KCNQ1OT1 lncRNA and not in the H19 ICR. The paternal origin possess methylation at ICR of H19 and not in the promoter of KCNQ1OT1 lncRNA which determines the expression of IGF2 leading to tumor progression.
References
1.
Vicente-DueñasC, HauerJ, CobaledaC, BorkhardtA, and Sánchez-GarcíaI, Trends Cancer4, 408 (2018).