
Abstract
Sphingomyelin (SM) is a major sphingolipid in mammalian cells. Although SM is enriched in the outer leaflet of the cell plasma membrane, lipids are also observed in the inner leaflet of the plasma membrane and intracellular organelles such as endolysosomes, the Golgi apparatus and nuclei. SM is postulated to form clusters with glycosphingolipids (GSLs), cholesterol (Chol), and other SM molecules through hydrophobic interactions and hydrogen bonding. Thus, different clusters composed of SM, SM/Chol, SM/GSL and SM/GSL/Chol with different stoichiometries may exist in biomembranes. In addition, SM monomers may be located in the glycerophospholipid-rich areas of membranes. Recently developed SM-binding proteins (SBPs) distinguish these different SM assemblies. Here, we summarize the effects of intrinsic factors regulating the lipid-binding specificity of SBPs and extrinsic factors, such as the lipid phase and lipid density, on SM recognition by SBPs. The combination of different SBPs revealed the heterogeneity of SM domains in biomembranes.
Introduction
Sphingomyelin (SM) (Figure 1A) is a major sphingolipid in mammalian cells. SM is distributed ubiquitously in the animal body and SM is enriched in the outer leaflet of the cell plasma membrane. However, SM is also observed in the inner leaflet of the plasma membrane (Murate et al., 2015) and intracellular organelles such as endolysosomes (Makino et al., 2015, 2017), the Golgi apparatus (Bakrac et al., 2010; Deng et al., 2016) and nuclei (Lazzarini et al., 2015). In the outer leaflet of the plasma membrane, SM forms specific lipid raft domains together with cholesterol (Chol; Levental et al., 2020; Lingwood & Simons, 2010). Lipid rafts are postulated to be involved in a number of pathophysiological events. Compared to the role of SM in cell surface lipid domains, little is known about the role of intracellular SM. However, recent results suggest that intracellular SM has important pathophysiological roles (Ellison et al., 2020).

Structures of different lipids: (A) SM, (B) dihydro-SM, (C) triple-SM, (D) S1P, (E) SPC (lyso SM) (F) 594neg-SM, (G) PC, (H) CPE, and (I) PE.
Measuring the localization of endogenous SM is essential to understand the physiological role of the lipid. The proposed size of lipid rafts is in the range of the submicron scale. However, methods to visualize SM at the submicron scale in a leaflet-specific manner are limited. The recent discoveries of SM-binding proteins (SBPs) enabled visualization of endogenous SM at the submicron scale (Kishimoto et al., 2016; Tomishige et al., 2021; Yamaji-Hasegawa et al., 2016). These proteins revealed the detailed distribution of SM in the plasma membrane (Abe et al., 2012; Kiyokawa et al., 2005), the transbilayer asymmetry of SM (Murate et al., 2015), SM trafficking from the Golgi apparatus (Deng et al., 2016) and exposure of SM to the cytoplasmic leaflet of lysosomes under pathological conditions (Ellison et al., 2020). In these methods, SBPs labelled by fluorophores or fluorescent proteins were added to the medium to label cell surface SM in living cells. Alternatively, fluorescent protein-conjugated SBPs were expressed in the cytoplasma to label SM in the cytoplasmic leaflet of the plasma membrane and intracellular organelles. To observe SM in fixed cells by fluorescence or electron microscopy, cells were labelled with SBPs followed by fluorescent or immunogold-labelled antibodies. Thus, the results are primarily dependent on the SM–SBP interaction, which may be affected by different intrinsic and extrinsic factors. Understanding these factors is crucial to the utilization of SBPs to visualize cellular SM. Here, we summarize intrinsic and extrinsic factors affecting SBP binding to SM. Recent protocols (Abe & Kobayashi, 2021; Tomishige et al., 2021) are also helpful for SBPs utilization. The plasmids of SBPs developed by our group are available through the RIKEN BioResource Research Center DNA Bank (https://dna.brc.riken.jp/en/).
Intrinsic Factors Regulating the Lipid-Binding Specificity of SBPs
SM Reporters
Lysenin
Lysenin is an earthworm-derived 33-kDa pore-forming toxin (Shogomori & Kobayashi, 2008; Yamaji et al., 1998; Yamaji-Hasegawa et al., 2003; Yilmaz et al., 2013) that specifically binds SM (KD = 5.3 × 10−9 M) (Yamaji et al., 1998). SM shares a phosphocholine head group with phosphatidylcholine (PC) (Figure 1G). However, enzyme-linked immunosorbent assay (ELISA) and thin-layer chromatography immunostaining assays indicated that lysenin bound SM but not PC (Yamaji et al., 1998). Lysenin also did not bind glycerophospholipids, ceramide, sphingosine, sphingosine-1-phosphate (S1P) (Figure 1D) or sphingosylphosphorylcholine (SPC, lyso SM) (Figure 1E) (Yamaji et al., 1998), suggesting that the ceramide-conjugated phosphocholine structure is recognized by lysenin. Lysenin bound dihydrosphingomyelin (dihydro-SM) (Figure 1B) and the SM analogue, (2S,3S)-{2-[(2-hexadecanoylamino-3-hydroxyoctadec-4-ynyloxy)-hydroxyphosphoryloxy]ethyl}trimethylammonium (triple-SM) (Figure 1C) (Kinoshita et al., 2013), suggesting that the double bond at the interfacial region of sphingosine in SM is not crucial for recognition by lysenin (Figure 2).

Binding of lysenin and EqtII–EGFP to SM, dihydro-SM and triple-SM. Binding of the proteins to the lipids was measured by ELISA as in Yamaji-Hasegawa et al. (2003).
The crystal structure of lysenin revealed an N-terminal pore-forming module and a C-terminal beta-trefoil motif (De Colibus et al., 2012). Both modules contain SM or phosphocholine-binding sites. An N-terminal truncation mutant of lysenin retained specific binding to SM (KD = 1.9 × 10−7 M) but was not toxic, suggesting that the C-terminal binding site is sufficient for SM binding. The shortest C-terminal fragment (amino acids 161–297) that binds SM was designated NT-Lys (Kiyokawa et al., 2005). Enhanced green fluorescent protein (EGFP)–NT-Lys bound SM but not SPC in ELISA (Makino et al., 2015). EGFP–NT-Lys also bound 594neg-SM, where ATTO594 dye is conjugated to propargyl-SM (Figure 1F) (Kinoshita et al., 2017), indicating that adding a fluorophore moiety to the headgroup of SM does not inhibit lysenin binding. A variant with a single tryptophan to alanine mutation at position 20 of lysenin, W20A, was nontoxic and weakly bound SM when conjugated to maltose-binding protein (MBP) (Kiyokawa et al., 2004) or glutathione S-transferase (GST) (Kulma et al., 2010).
Fluorescent protein-conjugated lysenin (Canals et al., 2010; Ishitsuka et al., 2004; Kidani et al., 2012; Yachi et al., 2012), lysenin binding followed by anti-lysenin antibody detection (Kavishwar et al., 2011; Makino et al., 2015; Taksir et al., 2012; Yamaji et al., 1998) and recombinant MBP- or GST-lysenin binding followed by anti-MBP or anti-GST detection (Kiyokawa et al., 2004; Kulma et al., 2012; Makino et al., 2015; Nakai et al., 2000; Skocaj et al., 2014; Yoshida et al., 2001) have been used to localize SM in model membranes (Ishitsuka et al., 2004; Makino et al., 2015), in the plasma membrane of fixed cells (Canals et al., 2010; Kavishwar et al., 2011; Kidani et al., 2012; Kulma et al., 2012; Nakai et al., 2000; Skocaj et al., 2014), in the endocytic compartments of fixed and permeabilized cells (Kiyokawa et al., 2004; Yachi et al., 2012; Yamaji et al., 1998) and the sections of cells and organs (Makino et al., 2015; Taksir et al., 2012; Yoshida et al., 2001). However, the binding of lysenin at physiological temperature was followed by oligomerization of the protein and pore formation in the membrane, which is accompanied by reorganization of the membrane (Alam et al., 2012; Aoki et al., 2010; Bokori-Brown et al., 2016; Podobnik et al., 2016; Yamaji-Hasegawa et al., 2003; Yilmaz & Kobayashi, 2015; Yilmaz et al., 2013, 2018). Oligomerization of lysenin is temperature-dependent (Yamaji-Hasegawa et al., 2003). Ikenouchi et al. (2012, 2013) incubated paraformaldehyde-fixed cells with GFP- or monomeric red fluorescent protein-tagged lysenin on ice followed by fixation on ice to prevent lysenin oligomerization. Using this procedure, they showed selective labelling of the apical membranes of mouse mammary gland-derived epithelial cells with lysenin in a tight junction-independent manner (Ikenouchi et al., 2012) and the enrichment of SM clusters in microvilli (Ikenouchi et al., 2013). Full-length lysenin or MBP-lysenin was also used to label SM in sodium dodecyl sulphate-digested freeze fracture replica labelling, where lipids were physically fixed to a metal cast (Murate et al., 2015). Atomic force microscopy tips conjugated with lysenin were used to measure the binding force of SM and lysenin, and localize SM-rich domains in a supported bilayer (Dumitru et al., 2018; Wang et al., 2012).
Nonoligomerizable, nontoxic NT-Lys has been used to label both fixed and living cells (Abe et al., 2012; Carquin et al., 2014; Ellison et al., 2020; He et al., 2017; Kasahara et al., 2013; Kishimoto et al., 2020; Kiyokawa et al., 2005; Leonard et al., 2018; Makino et al., 2015; Mizuno et al., 2011; Mound et al., 2017). To study living cells, NT-Lys was labelled with a fluorophore or fluorescent protein conjugated to the N-terminus of NT-Lys. Fluorescent NT-Lys has been used to monitor the dynamics of SM accumulation in the cleavage furrow during cell division (Abe et al., 2012), SM redistribution during cell attachment (Kishimoto et al., 2020) and SM exposure to the cytoplasmic leaflet of endolysosomes after bacterial infection (Ellison et al., 2020) in living cells. Recently, Bhattacharya et al. (2021) reported a unique application of NT-Lys to monitor enzymatic in vitro synthesis of SM.
Equinatoxin II
Equinatoxin II (EqtII) is a sea anemone-derived 20-kDa pore-forming toxin belonging to the actinoporin family (Anderluh et al., 1996; Rojko et al., 2016). Specific binding of EqtII and EqtII–EGFP to SM (KD = 7.5 × 10−9 M) (Hong et al., 2002) has been demonstrated by dot-blot (Bakrac et al., 2008), surface plasmon resonance (Bakrac et al., 2008) and ELISA (Makino et al., 2015). ELISA showed that EqtII–EGFP also binds SPC (Makino et al., 2015), dihydro-SM and triple-SM (Figure 2). The molecular mechanisms of pore formation by EqtII and another SM-binding actinoporin, sticholysin, have been extensively studied (Rivera-de-Torre et al., 2020; Rojko et al., 2016). Pore formation is a multiple-step process including binding to SM, an intramolecular conformational change in the protein, oligomerization on the membrane, and pore formation. Inhibition of the intramolecular conformational change by disulphide bridge formation in the double cysteine mutant EqtII(8–69) (V8C, K69C) prevented the haemolytic activity without perturbing SM-binding activity under oxidized conditions (Hong et al., 2002; Kristan et al., 2004; Rojko et al., 2013). Similar to EqtII–EGFP, EqtII(8–69)–EGFP bound SM and SPC, but not glycerophospholipids and glycosphingolipids (GSLs) by ELISA (Makino et al., 2015). Since the nontoxic property of EqtII(8–69) is dependent on the oxidative environment, application of this protein in the intracellular reduced condition is difficult. Recently, a nontoxic double mutant of EqtII, Eqt(V22W, Y108I) was reported (Deng et al., 2016). This protein was bound to the cell surface and the labelling was abolished by SM removal by sphingomyelinase (SMase). This protein was used to follow the release of SM-containing vesicles from the trans-Golgi network and SM transport to the plasma membrane in living cells (Deng et al., 2016).
SM/Chol Reporters
Aegerolysins
Mushroom-derived aegerolysin proteins are ∼15-kDa membrane-binding components of two-component pore-forming toxins (Butala et al., 2017). The binding of aegerolysins to the target membrane is followed by the recruitment of pore-forming subunits, oligomerization of the protein complex, and pore formation. However, aegerolysins themselves are nontoxic, although a high concentration of ostreolysin A (OlyA) induces plasma membrane shedding (Skocaj et al., 2016). Screening of SM/Chol-binding proteins from the extract of the mushroom Pleurotus eryngii identified the aegerolysin family protein, pleurotolysin A2 (PlyA2) (Bhat et al., 2013). PlyA2 is 80% identical to OlyA. C-terminally EGFP-tagged PlyA2 (PlyA2-EGFP) but not N-terminally tagged protein (EGFP-PlyA2) also bound SM/Chol but not GSL/Chol. Examining different sterols revealed the importance of the stereoconfiguration of the 3-hydroxyl group of sterol for binding (Bhat et al., 2013). PlyA2-EGFP labelled plasma membrane and late endosomes of HeLa cells. This binding was abolished by pretreatment of cells with SMase to remove SM or methyl-β-cyclodextrin to remove Chol (Bhat et al., 2013). Similar to PlyA2-EGFP, OlyA-mCherry bound SM/Chol and labelled living and fixed cells in an SM- and a Chol-dependent manner (Skocaj et al., 2014). The conformation of SM is speculated to be altered by the presence of Chol (Endapally et al., 2019). Aegerolysins recognize only Chol-bound conformations whereas lysenin and EqtII bind both Chol-bound and Chol-free conformations. Biochemical and structural analyses revealed that a single glutamic acid residue of OlyA is involved in the selective recognition of the Chol-bound conformation of SM (Endapally et al., 2019). Despite its specificity, the affinity of PlyA2-EGFP and OlyA-EGFP to SM/Chol was too weak to measure KD by quartz crystal microbalance with dissipation monitoring (QCM-D) (Bhat et al., 2015). Erylysin A (EryA) is 98% homologous to PlyA2, but EryA-EGFP does not bind SM/Chol or SM (Bhat et al., 2015; Panevska et al., 2019a). The lipid specificity of SM/Chol reporters was recently summarized (Grundner et al., 2021).
Nakanori
Nakanori (mid-raft rider in Japanese) is an additional SM/Chol-binding protein from the edible mushroom Grifola frondosa (Makino et al., 2017). Nakanori shows no sequence homology with other SM- or SM/Chol-binding proteins. However, the crystal structure of nakanori overlaps with that of the SM-binding toxin, sticholysin, with an extra N-terminus (Makino et al., 2017). Unlike sticholysin, nakanori was not toxic, perhaps because the extra N-terminus may inhibit oligomerization and pore formation. In contrast to aegerolysins, nakanori showed high affinity (KD = 1.4 × 10−7 M) to the SM/Chol membrane (Makino et al., 2017). Nakanori did not bind glycerolipids/Chol and GSL/Chol. More than 30 mol% Chol was required for binding to SM/Chol-containing membranes. Binding experiments of nakanori to SM/PC/Chol liposomes with different PC contents suggested that nakanori bound pre-existing SM/Chol domains and was not able to induce the formation of SM/Chol complexes (Makino et al., 2017). Nakanori identified SM/Chol-rich lipid domains on the cell surface and in intracellular membranes. Nakanori was also applied to follow the dynamics of SM/Chol domains on the cell surface by single-molecule tracking (Makino et al., 2017).
Ceramide Phosphoethanolamine (CPE) Reporters
Very weak binding of aegerolysins to SM/Chol led to the screening of the high-affinity lipid ligands of proteins. Screening identified CPE as a high-affinity ligand. CPE is an analogue of SM in which the phosphocholine moiety of SM is replaced with phosphoethanolamine (Murate et al., 2020; Panevska et al., 2019b) (Figure 1H). CPE is a major sphingolipid in insects and parasites but exists only in trace amounts in mammalian cells. The KD values of PlyA2-EGFP, OlyA-EGFP and EryA-EGFP to CPE/Chol (1:1) were 1.2 × 10−8, 1.2 × 10−9 and 1.3 × 10−9 M, respectively (Bhat et al., 2015). PlyA2 also weakly bound CPE in the absence of Chol (KD = 4.1 × 10−5 M) (Bhat et al., 2015). Similar to PlyA2-EGFP-binding to SM/sterol, the binding of OlyA-EGFP to CPE/sterol was dependent on the 3-hydroxyl group of sterol (Bhat et al., 2015). Phosphatidylethanolamine (PE) (Figure 1I) shares a small headgroup with CPE. However, aegerolysins did not bind PE/Chol (Bhat et al., 2015). PlyA2-EGFP revealed the enrichment of CPE in the central nervous system of Drosophila larvae and the EryA-EGFP stage-specifically labelled parasite Trypanosoma brucei (Bhat et al., 2015). PlyA2, OlyA and EryA showed selective toxicity towards western corn rootworm larvae and adults, and Colorado potato beetle larvae when mixed with pore-forming subunit PlyB (Panevska et al., 2019a). The W6A mutant of PlyA2-His inhibited the binding of the protein to CPE/PC but not SM/Chol and CPE/Chol. In contrast, the W28A mutant inhibited binding to SM/Chol but not CPE/PC (Bhat et al., 2015). Although the binding of PlyA2-EGFP to HeLa cells was completely inhibited by preincubation of the protein with SM/Chol liposomes, the binding was partially inhibited by CPE/Chol and was not affected by CPE/PC (Bhat et al., 2015). These results suggest that the binding site of PlyA2 to SM and CPE may be different. Similar to aegerolysins, nakanori bound CPE/Chol (Figure 3). However, the KD to CPE/Chol (1.9 × 10−6 M) was 100 to 1,000 times higher than those of aegerolysins. EqtII–EGFP but not EGFP–NT-Lys also bound CPE/Chol (Figure 4), which is interesting when we consider the physiological target of EqtII since CPE is widely distributed in Mollusca (Panevska et al., 2019b) who eats sea anemone, a producer of EqtII.

QCM sensorgram of the binding of nakanori to equimolar CPE/Chol membranes. Quartz crystal microbalance with dissipation monitoring (QCM-D) measurement was performed according to a published method (Bhat et al., 2015). The indicated concentrations of recombinant protein and PBS were added stepwise while ΔF was measured (red) and the dissociation constant (KD) of nakanori was estimated from the fitting curves of the association and dissociation (blue).

Binding of EGFP–NT-Lys and EqtII–EGFP to CPE/Chol and CPE/PC. Binding of the proteins to the lipids was measured by ELISA as in Yamaji-Hasegawa et al. (2003).
Extrinsic Factors: Effect of Phase and Lipid Density
Lysenin bound both the gel state {stearoyl SM (C18:0 SM, gel to liquid crystalline phase transition temperature, Tm, at 52.8°C [Bunow, 1979])} and liquid crystalline state {oleoyl SM (C18:1 SM, Tm at 33.0°C [Ahmad et al., 1985])} at 37°C (Yamaji-Hasegawa et al., 2003), suggesting that the phase state of SM does not affect lysenin binding. However, lysenin bound egg SM {eSM, a major component is palmitoyl SM (C16:0 SM, Tm at 41°C [Marsh, 2013])} in dioleoyl PC (diC18:1 PC, Tm at −20°C [Marsh, 2013]) but not in dipalmitoyl PC (diC16:0 PC, Tm at 41°C [Marsh, 2013]) (Ishitsuka et al., 2004; Makino et al., 2015). The stoichiometry of SM/lysenin was calculated to be 5 to 6 by isothermal calorimetry (ITC) (Ishitsuka & Kobayashi, 2007; Ishitsuka et al., 2004). In diC18:1 PC, SM is speculated to form clusters of >5 to 6 molecules. However, in diC16:0 PC, mixing C16:0 PC and SM hinders the formation of SM clusters (Ishitsuka et al., 2004; Makino et al., 2015). Most GSLs have high Tm. Similar to diC16:0 PC, GSLs inhibited lysenin binding to SM both in model and cell membranes (Ishitsuka et al., 2004; Kiyokawa et al., 2005; Makino et al., 2015).
In phase-separated eSM/diC18:1 PC/Chol giant unilamellar vesicles (GUVs), mCherry–NT-Lys bound exclusively liquid-ordered domains, whereas EqtII–EGFP selectively bound disordered domains (Figure 5) (Makino et al., 2015). Atomic force microscopy showed that in SM/diC18:1 PC supported bilayer, EGFP–NT-Lys bound SM-rich lipid domains, whereas EqtII–GFP bound SM-poor, diC18:1 PC-rich lipid domains (Figure 6) (Makino et al., 2015). Preferential partitioning of EqtII to the liquid disordered phase was also shown using phase-separated droplet interface bilayers (Rojko et al., 2014). However, in contrast to lysenin, EqtII bound a solid eSM/diC16:0 PC membrane (Makino et al., 2015). EqtII is speculated to bind SM when the density of lipids in the membrane is low. Clustering of SM may inhibit the binding of EqtII (Makino et al., 2015).

Binding of mCherry–NT-Lys and EqtII–GFP to GUVs composed of eSM/diC18:1PC/Chol (2:2:1). Fluorescence images displaying exclusive labelling were obtained by confocal microscopy. Scale bar =20 μm. Adapted from Makino et al. (2015).

Atomic force microscopy images of EGFP–NT-Lys and EqtII–GFP bound to different regions of the eSM/diC18:1PC bilayer. The scale bars next to each image indicate the height (nanometres). The scan sizes of images from top to bottom are 400 × 400 nm and 1 × 1 μm, respectively. The eSM/diC18:1PC bilayer separated into three phases: eSM-rich, eSM/diC18:1PC-mixed, and diC18:1PC-rich phases. On the images to the right, the borders of eSM-rich and eSM/diC18:1PC-mixed domains are indicated by dashed and solid lines, respectively. The white arrows point to the proteins. Adapted from Makino et al. (2015).
The binding of PlyA2, OlyA and nakanori required a high membrane concentration (>30%) of Chol. Thus, these proteins selectively bound liquid-ordered SM/Chol lipid domains. Super-resolution microscopy indicated that nakanori-positive SM/Chol-rich domains were part of the SM-rich domains labelled with NT-Lys (Figure 7).

PALM/dSTORM images of HeLa cells doubly labelled with pAmCherry-nakanori (green) and Alexa647–NT-Lys (red). The boxed area in the left panel is enlarged in the right panel. Adapted from Makino et al. (2017).
Conclusion and Perspectives
Table 1 summarizes the binding specificity of different SBPs and Figure 8 schematically shows the binding of different SBPs to various lipid domains. SM is speculated to form clusters with GSLs, Chol and other SM molecules through the hydrophobic interaction of the saturated acyl moiety and hydrogen bonds. Thus, different clusters composed of SM, SM/Chol, SM/GSL and SM/GSL/Chol with different stoichiometries may exist in the plasma membrane. In addition, SM monomers may be located in the glycerophospholipid-rich areas (Figure 8). Lysenin and NT-Lys bind SM and SM/Chol clusters but not SM/GSL clusters. In contrast to lysenin, EqtII binds SM/GSL clusters. Aegerolysins and nakanori selectively label SM/Chol clusters. SM/Chol clusters form domains belonging to the lysenin-labelled domains of SM clusters (Figure 7). Thus, each SBP highlights specific SM pools. Previously, different SBPs revealed alterations in SM distributions in pathological conditions (Makino et al., 2015, 2017).
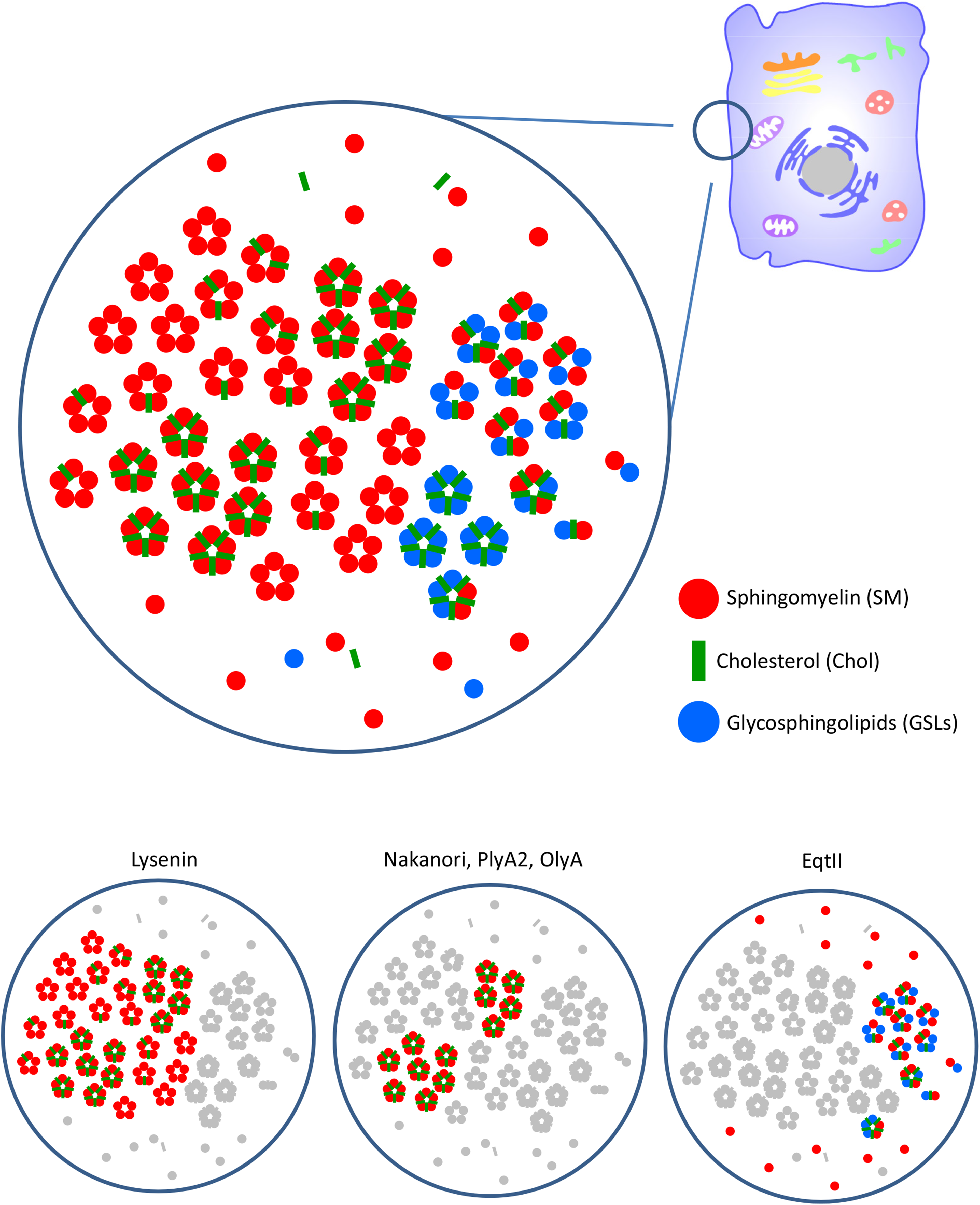
Speculative distribution of plasma membrane SM and other lipids detected by SBPs. SM forms lipid clusters with Chol and GSLs with different stoichiometries. Alternatively, SM distributes as a monomer in the glycerolipid-rich membrane. Glycerolipids are not shown in the model. Lysenin preferentially binds clusters of 5 to 6 SM molecules. Chol does not affect lysenin binding. Lysenin does not bind when SM forms a co-cluster with GSLs. Nakanori, PlyA2, and OlyA bind the SM/Chol clusters when the Chol/SM ratio is higher than 30%. EqtII does not bind SM clusters. EqtII labels the SM monomer and SM co-clustered with GSLs.
Lipid Specificity of Sphingomyelin-Binding Proteins.
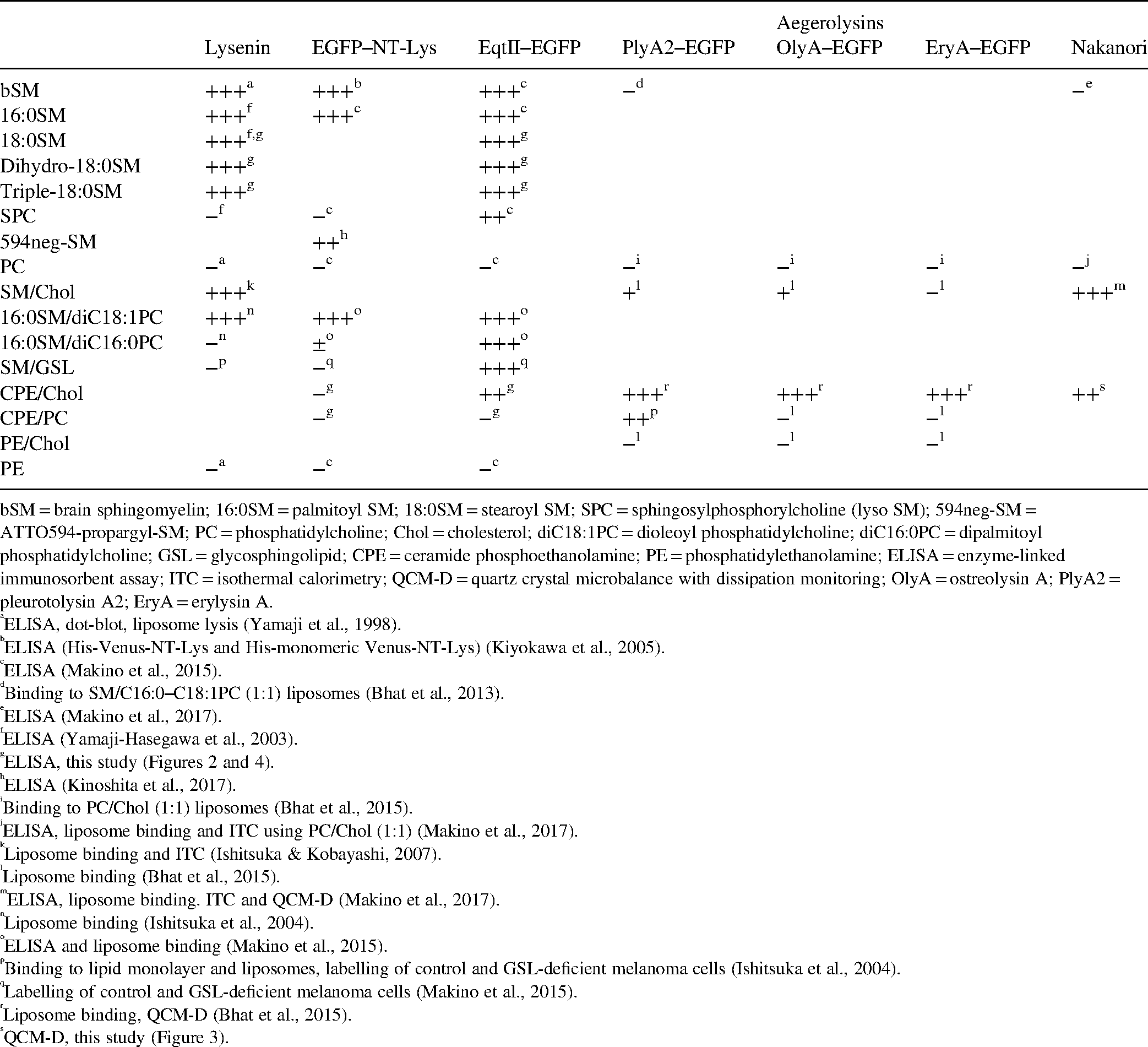
bSM = brain sphingomyelin; 16:0SM = palmitoyl SM; 18:0SM = stearoyl SM; SPC = sphingosylphosphorylcholine (lyso SM); 594neg-SM = ATTO594-propargyl-SM; PC = phosphatidylcholine; Chol = cholesterol; diC18:1PC = dioleoyl phosphatidylcholine; diC16:0PC = dipalmitoyl phosphatidylcholine; GSL = glycosphingolipid; CPE = ceramide phosphoethanolamine; PE = phosphatidylethanolamine; ELISA = enzyme-linked immunosorbent assay; ITC = isothermal calorimetry; QCM-D = quartz crystal microbalance with dissipation monitoring; OlyA = ostreolysin A; PlyA2 = pleurotolysin A2; EryA = erylysin A.
ELISA, dot-blot, liposome lysis (Yamaji et al., 1998).
ELISA (His-Venus-NT-Lys and His-monomeric Venus-NT-Lys) (Kiyokawa et al., 2005).
ELISA (Makino et al., 2015).
Binding to SM/C16:0–C18:1PC (1:1) liposomes (Bhat et al., 2013).
ELISA (Makino et al., 2017).
ELISA (Yamaji-Hasegawa et al., 2003).
ELISA (Kinoshita et al., 2017).
Binding to PC/Chol (1:1) liposomes (Bhat et al., 2015).
ELISA, liposome binding and ITC using PC/Chol (1:1) (Makino et al., 2017).
Liposome binding and ITC (Ishitsuka & Kobayashi, 2007).
Liposome binding (Bhat et al., 2015).
ELISA, liposome binding. ITC and QCM-D (Makino et al., 2017).
Liposome binding (Ishitsuka et al., 2004).
ELISA and liposome binding (Makino et al., 2015).
Binding to lipid monolayer and liposomes, labelling of control and GSL-deficient melanoma cells (Ishitsuka et al., 2004).
Labelling of control and GSL-deficient melanoma cells (Makino et al., 2015).
Liposome binding, QCM-D (Bhat et al., 2015).
QCM-D, this study (Figure 3).
In the cell membranes, SM rapidly diffuses laterally and moves from one membrane to another during endocytosis or budding. The binding of SBPs may partially inhibit SM dynamics. Indeed, the binding of OlyA changed the cell surface Chol pool and disrupted intracellular trafficking of Chol (Johnson et al., 2019). Inhibition of lipid dynamics by high concentrations of SBPs may explain membrane shedding induced by OlyA (Skocaj et al., 2016) and the influenza virus budding inhibition by nakanori (Makino et al., 2017). Thus, interpretation of the use of SBPs in live cell assays needs to be accompanied with some caution.
The distribution and dynamics of SM and SM/Chol-rich lipid domains remain controversial. Considering their precise binding specificity and potential drawbacks, SBPs appear to be useful tools, when combined with biochemical and biophysical analyses, to study the distribution, dynamics, and function of SM.
Footnotes
Acknowledgments
We are grateful to Brigitte Pollet for her technical assistance.
Declaration of Conflicting Interests
The authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The authors disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: Grants-in-Aid from the Ministry of Education, Culture, Sports, Science, and Technology of Japan (25293015 to T.K.), Agence Nationale pour la Recherche (A20R417C to T.K.), Agence Nationale de Recherche sur le Sida et les Hépatites virale (18365 to T.K.), Ligue Contre le Cancer (to T.K.), Vaincre les Maladies Lysosomales (19/LBPH/S44 to T.K.), Eucor seed money grant, RIKEN Integrated Lipidology Program and RIKEN Glyco-lipidologue Program (to T.K.).