
Abstract
Deathly circulations of highly pathogenic avian influenza (HPAI) stemming originally from the industrial poultry complex are causing mass mortality events in wild bird populations worldwide. In this article I explore what occurs as these drifting viral clouds envelop place. Focused on the islands of Orkney, Scotland, I describe my encounters with HPAI, and its effects on Papay's breeding seabirds—particularly its populations of solan geese (northern gannets), and bonxies (great skuas). Outlining the concept of viral atmospheres, which I condense from my experiences of mass avian mortality, as well as prior articulations of viruses as multispecies clouds, geographical understandings of atmospheres, and theories on the sociality of emotions, I use descriptions of these deathly encounters amidst the pervasive (and enveloping) meteorological phenomenon of Orkney to give emphasis to their various corporeal effects. These bodily consequences manifest not only in the tragic demise of many birds, but also the witnessing of this death, which shapes emotional response. Articulating how the practice of birdwatching forms an empathic relation with birds that reaches across the divide between two different embodied experiences’ of HPAI, I explore how the emotions engendered by such a connection form new ways of engaging with more-than-human worlds scarred by unruly capitalist impacts. In this instance, small, localised practices of care—such as collecting corpses to try and prevent the immediate spread of the virus—provide a way of navigating these novel realities, with personal and communal emotional experience of mass mortality producing currents of empathy that gather to alter the local trajectories of the outbreak.
I remember an early encounter with the virus. It was a day in June, near the beginning of my stay on Papay, an island in Orkney where I was researching seabird decline. 1 A cold sea fog, known locally as a haar, 2 had enveloped the island, and flights on the tiny eight-seater plane connecting it to the rest of the archipelago had all been cancelled. I was walking home along the beach that lines the eastern side of the island. Normally this commute is idyllic, stretching away in a curve of white sand bordered by clear, turquoise water. But on this occasion the vista disappeared into a murky gloom a few metres in front of me. Noticing a silhouette disturbing the smoothness of the water I stopped to look. It was a dead bird adrift on the tide and slowly being drawn into the shallows. Still fully feathered, its plumage moved with the tug of the waves, whilst its head and wings stirred up sand from the seabed. The corpse was that of an immature solan goose (northern gannet), 3 a large seabird. Amidst the strange atmosphere of the haar, the dead bird was an unsettling sight, but one I paid little heed. Death is not uncommon in the first few years of these animals’ lives (Wanless et al., 2006).
However, in the days following this encounter I began to see increasing numbers of sick birds floating on the water, resting on roads, or beached along the shoreline. Their posture—complete stillness, with wings and head folded to the body—became a familiar one I came to associate with the final hours before an individual's death. These birds, I was informed by the locals, were casualties of highly pathogenic avian influenza (HPAI). 4 A spate of infections caused by this virus had affected the island's birdlife the previous year, killing a few bonxies (great skuas), and its symptoms were recognised. But the spread of the illness to the solan geese and the number of birds visibly dying suggested a change in infection dynamics had occurred, and that the virus had shifted to new hosts (Lane et al., 2024).
These deaths form part of an ongoing HPAI panzootic stemming from the industrial poultry complex that has caused unprecedented numbers of deaths amongst populations of wild birds and poultry worldwide (ECDPC, 2020, 2022; WHO, 2023), prompting fears of zoonotic spillover. At the time of writing 71 countries in 5 continents have reported eruptions of HPAI, with the disease continuing to proliferate (WHO, 2023). Since October 2021, over 50,000 wild birds have died in the UK alone (Weston, 2023a). The outbreak of HPAI on Papay formed a distressing space of affective encounter that darkened my experiences of its spaces, and my memories of them.
It is these emotional spaces that form the subject of this article. In what follows I describe a viral atmosphere condensed from my experiences of mass avian mortality, prior theories of viral outbreaks as ‘multispecies clouds’ (Lowe, 2010: 626), geographical understandings of the affective nature of atmospheres (Anderson, 2009; McCormack, 2008), and conceptions of the sociality of emotions (Ahmed, 2014). In doing so, I argue that attending to the affectivity which registers corporeally through immersion in the atmospheres produced by viral outbreak reveals new, empathetic ways of engaging with more-than-human worlds scarred by unruly capitalist impacts. In this instance, small, localised practices of care—such as collecting corpses to try and prevent the immediate spread of the virus—provide a way of navigating these novel realities, with personal and communal emotional experience of mass mortality producing currents of empathy that gather to alter the local trajectories of the outbreak. 5
Through my attention to viral atmospheres, I make contributions to several other literatures. Firstly, by exploring the consequences of immersion in these viral clouds, I bring multispecies approaches to viral becoming back down to earth, grounding these understandings in ethical theory by highlighting how an emotional openness to the effects of the virus on others—a ‘shared-but-different vulnerability’ (Green and Ginn, 2014: 152)—can help shape the nature of outbreak responses. And secondly, my attention to viral atmospheres also answers recent calls in geography to attend to animals’ atmospheres (Lorimer et al., 2019). Here, I explore the interspecies sociality that emerges within the particular atmospheres produced by multispecies viral clouds. Focusing on birdwatching as a form of active participant observation in more-than-human worlds, I argue that the affinities that emerge through this practice establish an ‘embodied empathy’ pivotal to the emergence of this shared-but-different vulnerability to viral affect (Despret, 2013: 70). This process discloses new modes of attuning-to and knowing animal worlds, as well as revealing different ways that viral affects manifest in more-than-human social realms beyond infection.
I detail my experiences of this shared-but-different vulnerability to viral outbreak through a sensory immersion in the enveloping meteorological phenomenon of Orkney. Not only the previously mentioned haar, but other forms of weather; the capricious, seemingly perpetual drizzle known locally (but nowhere near exhaustively) as a mugga, a fissowy murr, or—worst of all—a more prolonged, windblown weet driv (Lamb, 2012; Marwick, 1921). Immersion in such weather created a palpable atmosphere that focused my attention on the immediacy of the virus's local consequences, as well as reinforcing and amplifying my sense of emotional saturation. Here, a ‘moodiness’ emergent from this combination of internal emotion and outer meteorology clagged around my involvements with these spaces (Ladino, 2015: 135; see Ahmed, 2014). This mood condensed from day-to-day interactions with landscapes marked by mass death and shadowed by fear of zoonotic spillover, forming an affective ambience ‘saturated’ with feelings of sadness, anxiety, and vulnerability (Ahmed, 2014: 11). These feelings were amplified by recent experience of a global pandemic similarly stemming from viral zoonotic disease—that of Covid-19. Indeed, the personal and communal sense of vulnerability and anxiety towards viral outbreak I articulate here no doubt stems from the shifts in outlook provoked by this event. Further, I note that much of the literature on viral becoming referenced here stems from before 2020. My explorations of the affectivity engendered by outbreaks of HPAI returns to these literatures after their worries have been realised, exploring how they have been altered by experience of zoonotic spillover.
In what follows I have divided my experiences with the virus into two encounters. The first of these turns to the birds met earlier, the solan geese. Through descriptions of my engagements with these creatures I highlight how the practice of birdwatching initially drew me into these viral atmospheres, allowing an empathic affinity to emerge. I then provide some background on the emergence of HPAI, highlighting its anthropogenic origins and the fears surrounding its potential for zoonotic transmission—both significant elements of the affective atmospheres engendered by this virus. A review of critical literature engaging with previous outbreaks of HPAI then follows. Here, I build on analyses of the biopolitics underpinning prior outbreak responses (e.g., Hinchliffe et al., 2013, 2016; Hinchliffe and Lavau, 2013), arguing that attention to emotion reveals new kinds of experience, knowledge, and practice that alter how multispecies relations unfold amidst ongoing proliferations of these diseases, offering an alternative to prevalent practices of securitisation and surveillance. To develop this, I then define the concept of viral atmospheres, which I use to frame my own experience of the emotional sociality of these outbreaks. I explore these further in the final empirical section and coda, which recount a day spent collecting the corpses of bonxies from their breeding ground on Papay. Here, the activity of corpse collection is articulated as a difficult act of care (Puig de la Bellacasa, 2010, 2012, 2017), with these interventions into the spaces of multispecies viral becoming stemming from felt, emotional connection to animal worlds—a process which also reveals a shared-but-different vulnerability to viral affects.
Sula
Recently, geographers have turned to the subject of animals’ atmospheres to reveal new aspects of the ‘affective, felt and emotional dimensions’ of animal worlds (Lorimer et al., 2019: 29). Animal atmospheres are perceived to emerge through social relation—perhaps as ecologies of fear between predator and prey (Brown et al., 1999), or through the sensory extravaganza of multispecies landscapes replete with shifting combinations of pheromones, sounds, and smells. But, as Jamie Lorimer et al. (2019) argue, attendance to these affective atmospheres also opens up new possibilities for engaging with animal life in the Anthropocene, highlighting not only new dimensions to animal worlds, but human impacts in them, such as aerial spraying of chemicals to control pests, or the disruptive shifts caused by climate change.
Yet there remain significant epistemic, methodological, and representational challenges to knowing and evoking these animal atmospheres (Lorimer et al., 2019). Gaining understandings of non-human sensory worlds, affective experiences, and how they are being disrupted presents a challenge, with risky, speculative ethological approaches to these realms and their meanings often accused either of anthropomorphism, or of imperilling the objective distance of proper scientific practice (Despret, 2013). Focusing on the presence of the researcher in these accounts, however, can foster new knowledge of human-animal relations (Lorimer et al., 2019). Indeed, for Despret (2013), attending to the position of the observer/scientist's body in relation to animal subjects can nurture more sympathetic attitudes. Here, ‘the scientist risks being touched/affected by what matters for the animal he/she observes’ (Ibid.: 57), opening up an ‘[e]mbodied empathy’ with their subjects (Ibid.: 70). As Lorimer et al. (2019: 34) argue, this empathetic ethological approach offers the possibility ‘not simply to attune to animals, but to attune with them’, revealing not only something of particular animals’ lifeworlds, but allowing felt connections to these worlds to emerge, fostering new understandings of more-than-human relational phenomena. Building on these ideas, in this section I reflect on my own process of attunement to the emotional atmosphere of the viral cloud. Here I find the practice of birdwatching offered a way of ‘learning to be affected’ by avian experience of HPAI (Lorimer et al., 2019: 34; see Despret, 2013), of empathising, and so attuning to a ‘shared-but-different vulnerability’ to the viruses’ effects as they manifest in a more-than-human social realm (Green and Ginn, 2014: 152).
The practice of birdwatching I focus on is a specific one. Rather than an act of rigorous ornithological observation, or the extremes of twitching (the single-minded pursuit of rarities and vagrants to tick them off a list), birdwatching is a casual activity that entangles the becoming of the self with others (Bonta, 2010). To theorise this practice on a personal level, I would say that I enjoy the company of birds. I find the moments where I am distracted from my narrow concerns and reminded of the plurality of overlapping worlds by a fleeting encounter to be meaningful. Similar to Hayden Lorimer's (2010) confrontations with seals whilst running, or Kathleen Jamie's (2005: 39) feminist take on her own engagements with birds, which she notices ‘[b]etween the laundry and fetching the kids from school’, my approach here theorises these encounters as a vitalist ethology. Here, momentary conscious attention to ‘the sheer vibrancy’ of the world's ongoing becoming briefly reveals the differing rhythms of interspecies sociality underpinning more-than-human emergences, facilitating understandings of how relations materialise (Lorimer, 2010: 60). In developing this, my focus lingers on the solan geese, the birds encountered earlier in the introduction. As I got to know Papay I frequently saw these birds flying offshore on foraging trips whilst I proceeded on my own: the hour's walk to-and-from the island shop. Often stopping, I would watch the birds tip steeply into angled or near-vertical plunges in the presence of a shoal, folding their wings back at the very last moment so they pierced the water like an arrow. A fruitful dive would often precipitate an excited shuffle in the surfacing bird, who would shake its head, and dip its beak in the water.
As more-than-representational theories stemming from geography have sought to articulate and explore, life gains meaning through such brief, everyday encounters, and unexceptional interactions (Lorimer, 2005). For Hayden Lorimer (2010), ethological accounts blur geographical and biological knowledge with personal reflection, storying a more-than-human sociality that emerges when open-minded observation rubs up against animal lives that are always ontologically other. Such expression not only chronicles animal lives, but also narrates how interspecies sociality is involved in the figuring of self. Emotions are a key set of relations through which these transient, and often ordinary relational encounters are comprehended (Anderson and Smith, 2001). Indeed, it was from my routine observations of the solan geese that the affective atmosphere of the viral outbreak emerged. My initial fondness for these birds—which emerged through these passing observational moments—lead to a curiosity about them, and a desire to visit their nearby colony to deepen my understandings of their lifeworlds.
Solan geese do not breed on Papay. The birds I encountered amidst my day-to-day activities would have come from the neighbouring island of Westray, where they nest on the cliffs at Noup Head (Figure 1). This colony is young, with the first birds noted to breed here in 2003 (Wanless et al., 2005). Since then, counts made almost annually have revealed an increasing population, with an estimate of at least 1500 apparently occupied nests in 2022 (RSPB unpublished), though a 2016 aerial survey suggests a far higher figure (Murray et al., 2017). The scenery on Westray is dramatic. The cliffs are over 70 metres high, dropping sheer into the Atlantic below. They are also packed with seabirds—over 30,000 breed along a two-mile-long section of coast here (RSPB, personal communication), dwarfing anything experienceable on Papay.
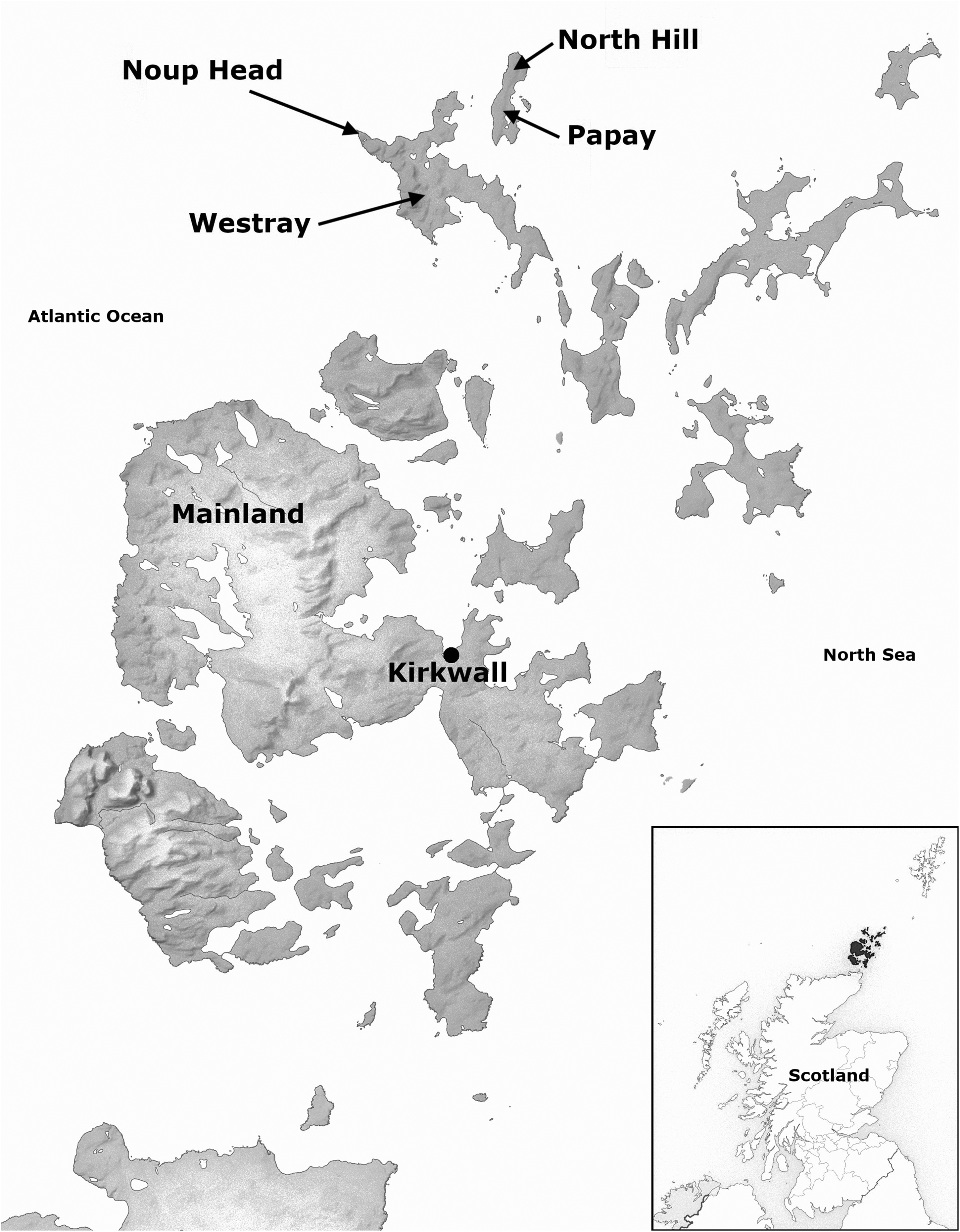
Map of Orkney showing key locations mentioned in this article. Image: Wikimedia Commons.
I remember my visit to these cliffs vividly. Similar to my encounters with the virus in the haar described above, it was a day of eerie meteorological envelopment. Out to sea, a grey, vaporous mugga—a soft, drizzling mist—drifted on the wind, masking the horizon (see Lamb, 2012), forming an atmosphere of enclosure that drew attention to and amplified the noise of the seabird host below. A little way along the clifftop path, a promontory offered a view of the gannetry. The face of the cliff was composed from layers of eroded strata, stained white with guano, which gave it an oozing quality, as if the colony was dripping downwards (Figure 2). The profusion was astonishing. Innumerable solan geese lined the ledges and intermingled with them on narrower shelves were guillemots and kittiwakes. Many more birds flew, circling, diving, and wheeling above the dark water washing in and out of huge sea caves at the base.

Seabird colony at Noup Head. The birds visible are mainly common guillemots, which were less affected by the HPAI outbreak at this locale. Image © the author.
As I stood there, the drifting mist and rain shifted direction, amassing around the promontory and catching in my hair and eyelashes. No longer afforded a broader view of the spectacle I reached for my binoculars and began to scan the cliff face that had become suddenly ghostly amidst the cloud, searching for the birds. I was presented with a bleak sight. On many of the seaweed nests were the spreadeagled carcasses of adult gannets. Occasionally the fluffy corpse of a chick lay beneath them. A movement below drew my gaze downwards to the water. The bodies of more birds, both chicks and adults were floating face down on the shifting surface. The virus seemed to have struck randomly; amongst all the death some individuals had evaded infection, and in some instances had incorporated the bodies of the dead into their nest structures. Gannets cement their nests to the narrow ledges and slopes of their colonies by voiding sticky, viscous excreta onto its sides, but project their ‘whitewash’ clear of the site (Nelson, 1978: 126). Given the oral-faecal spread of avian influenza viruses it is not difficult to see how HPAI might spread through such spaces (Lane et al., 2024). Here the nest itself, as well as its wider vicinity become contaminated environments, capable of infecting those who come into proximity.
Within the vaporous confines of the mugga, this deathly spectacle left a profound impression. My affectionate orientation towards these birds moulded the affective nature of this contact into emotion (see Ahmed, 2006, 2014). Feelings of horror, anxiety, and sadness reworked my being. Held in close vicinity with mass mortality by the atmospheric conditions, these emotions clagged around me, and I found myself altered; reshaped by contact with the virus. Though this differed from the infection that afflicted the birds, I was nonetheless affected by its proximity. Here, the casual activity of birdwatching formed a ‘partial connection’ at the margins of two different embodied experiences’ of HPAI (Despret, 2013: 61). Through my attunement to these birds’ experience, an empathic affinity emerged that formed the grounds for my subsequent care work in these spaces of viral relation—collecting carcasses to try and reduce its spread amongst Papay's birds. Building towards this, I now provide context by tracing the viral reservoir from which these outbreaks emerged.
Viral reservoir
The global spread of these outbreaks follows ‘tragic patterns’ (Hinchliffe, 2004: 228) that map to the spaces of avian migrations. HPAI was first detected in UK seabirds during the summer of 2021 when bonxies began dying at their breeding sites on Fair Isle in Shetland after returning from their pelagic wintering grounds. Hundreds were witnessed to die. Further large-scale dying's then proceeded to occur across multiple Scottish islands with little population interchange, reflecting independent incursions of the disease (Banyard et al., 2022). Then, between October 2021 and March 2022, over 13,000 barnacle geese—a third of the population—died around the Solway Firth where they overwinter following migration from Svalbard (RSPB, n.d.). My own encounters with seabird deaths in Orkney during the summer of 2022 follow on the wings of these mass mortalities, tracing a small element of the virus's re-emergence the following breeding season.
HPAI is a zoonosis; a disease harboured in non-human animals but capable of being transmitted to humans (Keck and Lynteris, 2018). The lineage of HPAI causing these unprecedented numbers of avian deaths has been traced back to a precursor first isolated in 1996 from farmed geese in Guangdong Province, China (Wan, 2012; WHO, 2023). Reassortments of this virus were responsible for the outbreak of H5N1 that caused a cluster of human deaths in Hong Kong in 1997. The high lethality of these human cases elicited concern regarding the potential of this new virus to cause a deadly pandemic—a fear that has coloured all further outbreaks of this disease worldwide (Keck, 2022).
Anxiety regarding viral spillover stems from previous outbreaks of infectious zoonotic diseases, including the 1918–20 flu pandemic, Ebola, the HIV/AIDS epidemic, and—more recently—Covid-19. Fear that similar contagions caused by novel pathogens jumping from animal populations to spark deadly pandemics have radically altered responses to disease outbreaks globally (Lakoff, 2017). Modelling the human risks posed by particular diseases now occurs through imagining and preparing for the ‘worst-case scenario’ (Keck, 2022: 7). Indeed, following discovery that the source of these deadly human infections in Hong Kong was poultry, more than 1.5 million chickens in the city's farms and markets were rapidly culled to prevent the virus's spread (Chan, 2002). However, over the next six years the virus continued to circulate in factory-farm units in Southern China, where it underwent a series of reassortment events. In early 2003, more human cases were detected in Hong Kong, stemming from one of these novel genetic variants (Li et al., 2004; Webster et al., 2006). Attempts at containment this time were less successful, and from the end of 2003, outbreaks of HPAI spread across China and into Thailand, Vietnam, Indonesia, Laos, Cambodia, Korea, Japan, and Malaysia, becoming endemic within poultry populations (Li et al., 2004).
During this eruption, the virus once again jumped species, infecting over 440 people and killing 262 of them (Wallace, 2009). Amongst these cases were a few incidents of human-to-human transmission (Kandun et al., 2006; Yang et al., 2007), again raising fears that this outbreak might result in a deadly global pandemic (Lowe, 2010). In May 2005, another novel variant of HPAI was identified near Qinghai Lake in northwest China, this time amongst migratory wild bar-headed geese (Chen et al., 2005). This lake, the largest in China, is an important stopover point on several bird migration routes across Asia. It is also surrounded by large-scale poultry farms, and an interlinked fish farm industry that uses chicken faeces as feed (Wilbert, 2006). From this outbreak the virus became transcontinental, spreading across Europe and Africa (Salzberg et al., 2007). The mechanism for such spread seems not to be long distance avian migration, but rather the slow transfer of the virus over short distances as birds move and succumb to the disease, as well as trade in live poultry and their products (Weber and Stilianakis, 2007).
Low pathogenic strains of avian flu are endemic in wild birds. They have evolved alongside such populations and rarely kill their hosts (Capua and Alexander, 2006; Perez and de Wit, 2017; Wilbert, 2006). The highly pathogenic strains killing so many birds appear to be a poultry virus, likely evolving from low pathogenic forms within the crowded conditions of factory farms (Capua and Alexander, 2006). Here, large-scale, intensive poultry production units act as ‘factories’ for developing more virulent strains (Wilbert, 2006: 3). Viruses, such as influenzas, constantly replicate themselves, but do so unfaithfully, producing genetic mutations that allow them to adapt to changes in host environment and perpetuate a chain of transmission as immunity develops (Lowe, 2010). Within these industrial monocultures of genetically identical animals, extreme proximity and poor living conditions stress birds, depressing their immune systems and enabling easy transmission of viruses and other diseases amongst the units’ huge captive populations. Further, high industrial throughput constantly renews a reservoir of susceptible hosts. Usually, an epidemic caused by a particular strain of a virus will burn out once it has run out of hosts, either through the development of immunity, or extreme levels of mortality. But the constant replenishment of these farms with susceptible poultry lacking immunity, and the removal of birds either for slaughter or culling creates conditions in which a perpetual outbreak can occur, enabling pathogens to develop greater virulence (Wallace, 2009). Increasing production rates in these units also cause resident influenza viruses to increase their transmission rate, furnishing this extreme virulence with an ability to spread at speed (Wallace, 2009).
Attention to how reservoirs of low-pathogenic disease exist within wild bird populations reiterate how bodies are always multispecies mixes (Haraway, 2008; Hinchliffe, 2004). Animals have a kinship with the viruses they host and co-evolve alongside (Fuentes and Porter, 2018). Within the human body, the number of viruses outstrips both human and bacterial cells, forming a significant element of its genetic material (Lowe, 2014). The remnants of retroviruses in the human genome have an evolutionary role, guiding the mutation of DNA (MacPhail, 2004). For example, the viral gene, syncytin, which is found in mammals, codes for placental proteins that enable a foetus to draw nutrients from its mother, indicating that viral co-becoming has been key to the emergence and continuance of mammalian life (Lowe, 2014). Clearly, then, it is not ‘pathogens per se’ that are the issue here, but rather the production of increasingly extreme pathogenicity (Hinchliffe et al., 2016: 71). Underlying these outbreaks of HPAI are particular interventions into animal becoming that have sought to direct it towards multinational profitability—a fact which alters the complexion of my encounter with the solan geese on Westray. Amidst the atmospheric confines of the mugga I was pulled into proximity not only with mass death, but also the spatiotemporally dislocated violences towards avian life stemming from intensive, industrial poultry production, as well as the threat of zoonotic contagion.
Outbreak response
Multiple responses stemming from geography, anthropology, and the environmental humanities have engaged with HPAI and the threat of contagion it promises (e.g., Lowe, 2010, 2014; Porter, 2012; Wallace, 2009; Wilbert, 2006). These include the aforementioned critical explorations of the origins of these viruses (Wallace, 2009; Wilbert, 2006). Acting as a counter to industry and governmental attempts to blame these outbreaks on wild birds and/or small-scale poultry producers, these ‘political virologies’ reveal something of the bare life of animals caught within intensive farming systems (Wallace, 2009: 916). They also emphasise the influence of neoliberal deregulation in developing the high-throughput factory systems that have modified avian life and its pathologies so completely within the past few decades (Wallace, 2009). Elsewhere, much anthropological literature analyses pandemic preparedness, highlighting how its anticipatory visions have shaped society, forming new publics and regulatory states (Caduff, 2015, 2021), new systems of biopolitical surveillance (Keck, 2020; Lynteris, 2016), and moulding imaginaries of human extinction (Lynteris, 2019).
Alongside these critiques, other responses to HPAI engage with how these outbreaks have overwhelmed attempts at containment (Hinchliffe et al., 2013; Hinchliffe and Lavau, 2013). Here, biosecurity is revealed as a practice that shores-up certain conceptualisations of disease; ones in which outbreaks can be avoided through walling-off populations from an outside world of infection. Like assumptions around healthy bodies as pure spaces free of pathogens, toxins, and pollutants, these strictly demarcated territories often fail to acknowledge the relationality of disease—that animal life and its pathogens have co-evolved and are wildly intertwined (Hinchliffe and Lavau, 2013; see Haraway, 2008; Lowe, 2014). As highlighted, biosecure spaces are implicated in the co-production of zoonoses. The walls erected to keep disease out also enclose it within, confining populations of birds and their pathogens within conditions that enable their rapid mutation (Hinchliffe et al., 2013). Once again these criticisms challenge narratives scapegoating wild birds, rather than finding issue with the apparently secured populations of factory-farms (Hinchliffe et al., 2013; Lowe, 2010).
Also scrutinised in these explorations of biosecurity is how these outbreaks have been used to justify new regimes of biopolitical power. Biosecurity is defined here as a political technology geared towards governing ‘the disorder of biological life,’ aiming to refigure its ongoing becoming along more manageable lines (Braun, 2007: 23). Following initial outbreaks of HPAI, these biopolitical interventions have caused a proliferation of monitoring activities designed to watch for eruptions of disease (see Keck, 2022). One element of this has reconfigured the knowledges and observations of birdwatchers and ornithologists as ‘a kind of border patrol, a first line of veterinary surveillance’ (Wilbert, 2006: 2). Analyses of this surveillance culture has highlighted again how scrutiny of wild birds deflects critical attention away from the real cause of concern, which is how food is produced and traded; ‘the harm being done—ecologically, socially, and to animals—by agribusiness (Wilbert, 2006: 3). Further, this invitation for members of the public to join collective regimes of surveillance enables a creeping governance of human and nonhuman lives to extend outwards, integrating even more elements of life into secure frameworks designed only to service and protect the health of human populations (Hinchliffe and Lavau, 2013; Wilbert, 2006).
Building on this, much criticism of biosecurity has sought to conceptualise a space in which the complexity of viral relations can be communicated beyond strict inside/outside dichotomies, and in ways that jettison desires ‘to govern the ‘global biological’’ (Braun, 2007: 21). Expression here is given to viral borderlands (Hinchliffe et al., 2013), to viral intra-active worlds (Wilbert, 2006), and to multispecies viral clouds (Lowe, 2010). These frameworks all speak in different ways to the fundamental relationality of disease; the fact that illness always emerges from and pulls us into connection with multiple others (Karesh et al., 2012). In voicing this relationality, these articulations are aligned with post-humanist attempts to explore how particular human lives and cultures are bound up with other entities (Bingham, 2006; Haraway, 2008; Hinchliffe, 2007; Kirksey and Helmreich, 2010; Whatmore, 2006).
Yet, emerging zoonotic diseases challenge these concerns—learning to live with nonhuman life becomes difficult with the prospect of a deadly pandemic emerging from those relations. In the context of HPAI, this has caused an early abandoning of more responsive engagements with non-human worlds in favour of approaches that reinforce a Foucauldian power over life (Hinchliffe and Lavau, 2013). Challenging this reversion, Hinchliffe et al. (2016) argue how differing forms of attention to viruses and their hosts open up multiple forms of political relation. Pointing to how the ‘gentle science of ornithology’ has been co-opted as a surveillance practice, they argue that the multisensory observational practices that underpin this practice are not just acts of indifferent scrutiny (Hinchliffe et al., 2016: 201). They emerge from a pre-existing inclination or sensitivity towards non-human others. Observation here is opposed to watchfulness, offering a softer form of ‘knowing around’ a subject. To observe birds is to be ‘in amongst things,’ responsive and open to the ‘generativity of relations’ (Hinchliffe et al., 2016: 205; see Barad, 2007; Serres, 2008)—a far cry from the distanced, unidirectional gaze implied by surveillance. Such knowing, I find, dovetails with Lorimer's (2010) aforementioned explorations of convergences of ethology and phenomenology, as well as my own birdwatching. Attention in these approaches is relational, and points to how the self, as a ‘nexus…[of] knowledge, feeling and memory’ is compelled to think, feel, and act differently’ by the presence of others (Brown, 2011: 165; Hinchliffe et al., 2016). For Lorimer (2010; see Despret, 2013), attentiveness to animal lives is both a form of witnessing and a process of learning how to be affected by the social interactions encountered amidst more-than-human profusions. Such approaches—ones in which ‘all parties are altered as relations take shape’ (Hinchliffe et al., 2016: 205; see Barad, 2007)—point towards a different sort of engagement with viral outbreaks.
Reaching beyond attempts to merely police relations for a human good, attention to how the self is reshaped emotionally by confrontations with mass avian death enables the possibility of a different kind of ‘life politics’ in the viral borderlands produced by HPAI (Hinchliffe et al., 2016: 245). Emotional connection, such as that produced through birdwatching, forms a solidarity with nonhuman lives that offers an alternative to the creeping forms of securitisation and surveillance that protect and entrench the systems underlying HPAI's emergence (see Keck, 2022). Indeed, my focus on emotion here is important, revealing new kinds of experience, knowledge, and practice that alter how multispecies relations unfold in the shadow of viral outbreak. This is something that comes to the fore in the second empirical exploration of the virus's effects I detail here—the collecting of dead birds to try and alter the local trajectory of the virus's spread.
Viral atmospheres
But before I describe this encounter, I first need to outline the concept of viral atmospheres through which I explore the emotions engendered by immersion in these viral clouds. I define this by delving into conceptual debates around affect and emotion. However, tying this to the subject at hand, I undertake this alongside a discussion of Lowe's (2010) theorisations of viral outbreaks as multispecies clouds. This framework for understanding viral relations emerges from the fact that viruses, as infectious agents, enact their identities with others. Rather than existing as well-defined populations, viruses form quasi-species, which contain myriad variant genomes co-opted from the cells of their hosts. Quasi-species are complex becomings fuelled by ongoing mutations that arise as viral replication proceeds, and are often termed mutant spectra, swarms, or clouds (Domingo and Perales, 2019). Centred around her experiences in Indonesia as outbreaks of HPAI swept through the country in 2003, Lowe (2010) highlights how during the virus's spread multiple ontologies were altered through encounters with these clouds. Not only were the immune systems of myriad beings transformed, so too were the lives of millions of Indonesians, as well as the organisations and institutions tasked with responding to the outbreak. In this context Lowe (2010: 626) writes that amidst encounters with viruses ‘one can think… not only of quasi-species clouds but also of “multispecies clouds,” collections of species transforming together in both ordinary and surprising ways.’
Exploring how these multispecies clouds are experienced in place, Lowe notes how they formed a complex atmosphere, intermingling the meteorological, the epidemiological, and the emotional. Infectious viral particles—spread through droplets of spittle and other bodily fluids—lent the humid atmosphere of tropical Southeast Asia an affective, enveloping ambience that found emotional expression in a pervasive mist of uncertainty, fear, and ‘apocalyptic dreams’ (Lowe, 2010: 625). It is the effects of envelopment that I focus on here as I develop the conceptual framework for my own explorations of the emotional experience of living amidst circulations of deadly pathogens. To do so, I bring Lowe's multispecies clouds into conversation with geographical theories on atmospheres. In these, atmospheres, whether meteorological or otherwise, have been defined through their affectivity: as things dispersed yet palpable, which register on sensing bodies whilst remaining formless (McCormack, 2008). Here, envelopment is key to examining the relations between atmospheres and the entities contained within them. Atmospheres are forceful bodies that contribute to the ongoing emergence of feeling—and its transmission—across beings immersed inside them (Anderson, 2009; see Brennan, 2004). But atmospheres are also more than just spaces in which bodies are situated; they are emergent, dynamic processes that materialise from the ability all bodies have to affect one another (McCormack, 2018). To be enveloped is to be involved in the perpetuation or slow transformation of atmospheric mood, which itself acts forcefully on the enclosed body—a sort of affective feedback loop. Envelopment reveals a form of continuity between entities and the elemental conditions in which they participate, one in which all involved are better understood as ‘shifting fields of materiality’ rather than as fixed bodies (McCormack, 2008: 414).
Such transformations are visible in Lowe's (2010) descriptions of how the climate of fear surrounding HPAI's spread through Indonesia dies down. With time, the feeling of immediate personal danger she experiences diminishes, and a more relaxed curiosity about the situation emerges. Aware that poultry were the most significant casualties of the virus, and that backyard flocks had especially been targets of blame, she purchases some chickens and watches the birds, ‘taking field notes as an amateur ethologist’ as they build a nest in a tree (Lowe, 2010: 637). Over this period, she also notes how the atmosphere of relief amongst her neighbours and interviewees morphs into a sometimes-angry scepticism about curtailed freedoms as the predicted pandemic fails to emerge, leading to a proliferation of conspiracy theories. Visible here is a sense of how atmospheres act as a shared ground from which subjective states emerge. But these atmospheres shift, never achieving stability. They arise from currents of transpersonal affectivity that bloom and disperse as different bodies enter into relation with one another (Anderson, 2009). In the context of HPAI, currents of pre-personal affectivity coalesce into named emotions, which act as socio-cultural expressions of felt intensity stemming from multispecies viral relations—fear, relief, anger (McCormack, 2008; see Anderson, 2006; Anderson and Smith, 2001; Shouse, 2005).
Turning now more fully to the subject of emotion, the continuities between affect and emotion articulated here follow established theorisations of these categories in which differences between them are perceived in terms of intensity or degree, rather than quality or kind. Affects can acquire the complexity of emotion, and emotion can denature into affect (Ngai, 2005). For Ahmed (2014: 6; see Anderson, 2009; Stewart, 2007), affects are felt intensities that emerge as different bodies come into contact and leave ‘impressions’ on one another. Such impressions precede cognitive definition, causing inarticulable psychological shifts in response to particular situations (Brennan, 2004; Ladino, 2015). Emotions are often defined as the socio-cultural voicings of these felt intensities (McCormack, 2008), but ones that are still deeply personal and subjective (Ladino, 2015).
However, this is not to say that emotions are feelings that belong to individuals and are projected outwards, perhaps being reflected back to us as a sense of ‘fellow-feeling’ (Ahmed, 2014: 9). Neither do emotions emerge from a collective elsewhere and infect us like a contagion (Ahmed, 2014; Ladino, 2015; see Brennan, 2004). Rather, following Ahmed (2014: 10) emotions are impressions that are left on the surface of the ‘I’ or the ‘we’ by contact with others, producing the borders that allow the individual and the social to be delineated. Such surfaces emerge from the way that circulations of affect rarely register identically across individuals— ‘even when we feel we have the same feeling, we don’t necessarily have the same relationship to that feeling’ (Ahmed, 2014: 10). Affective experiences are felt differently, multiplying subjective worlds, and introducing variance into collective perceptions of events. Much like the viral clouds of mutant spectra described previously, clouds of shared feeling ‘saturated with affect’ clag around us, ‘like a thickness in the air, or an atmosphere,’ enacting and altering identities on an individual and social level (Ahmed, 2014: 10–11).
The viral atmospheres condensed from these theories frame my explorations of the emotional sociality of these outbreaks. Combined with my sensory immersion in the enveloping meteorological phenomenon of Orkney, they offer the means to explore the confines of this affective cloud of mutant spectra as it enveloped the archipelago. If Lowe's (2014) concept of multispecies clouds reveals how multiple beings are transformed by viral outbreak, then attention to the atmospheric conditions precipitated by these clouds—the different ways individuals immersed with them are altered by their affectivity, and how this shifts personal and communal identities—is key. Immersion in these atmospheres reveals how the virus's touch does not have to infect to be affective. Within these atmospheres, currents of affectivity caused by witnessing the virus's consequences for others coalesce into emotional experiences that reshape the individual, altering their relations with the more-than-human social realms they are a part of. Like a fissowy murr, a drizzling rain that floats in the air like smoke, they blur established boundaries between species. In the next section, I detail a further empirical encounter with the virus where these ideas are borne out. Then, in a final coda I reflect on how the affective atmosphere of this viral cloud and the empathy it precipitated sparked responses that sought to alter the local trajectories of the outbreak for avian others.
Carcass collection
As a place to dispel the gloom cast by dreary weather and viral threat, or at least temporarily drown it, the Papay pub offers all you could need. Holding unconventional premises compared to a traditional public house, it is a uniquely informal place. On Saturday evenings a cupboard in the corner of the community common room is opened to reveal a carousel of half-empty spirits, and a makeshift landlord is elected; drawn either from a revolving cadre of islanders, or just from whoever is standing near the bar at the time. One midsummer's night after my encounters on Westray with the solan geese I find myself there drinking whisky with the locals, whilst the prolonged brightness of the northern dusk slants through the window. Sometime after midnight, just as it finally manages to get dark and I’m thinking of heading home, an invitation comes my way for the following day— ‘come and scrape the dead birdies off the hill.’
The hill referred to here is North Hill, an expanse of treeless maritime heath that dominates the northern third of Papay. It was once (and still is) common land used by islanders to graze livestock but is also now a nature reserve managed by the North Hill Grazing Committee in conjunction with RSPB Scotland and NatureScot. The habitat this reserve protects is important for its globally rare vegetal community and its many breeding birds, including arctic terns, bonxies, arctic skuas, curlew, and dunlin. The space itself is dominated by the island's highest point, Errival, a hill which slopes down to low sea-cliffs. But it is also dotted with numerous pools of water. During the Second World War, the northern part of the island was used by the Royal Navy as a firing range to practice sea-to-land bombardment in preparation for the 1944 D-Day landings, and the spaces of the reserve are still pockmarked with shell craters. Some of these are now water-filled, providing communal bathing spots and foraging spaces for the birds that breed here in the summer months.
The dead birds, I am told, are predominantly bonxies, a kind of seabird with webbed feet, sharp talons and a powerfully hooked beak. Bonxies are apex predators within the marine ecologies where they are present (Olsen and Larsson, 1997). They have broad diets: hunting fish by splash-diving onto shoals, foraging on discards from fishing vessels, and piratically harrying other birds to either steal their catch, or to predate them (Furness, 1987; Jakubas et al., 2018; Phillips et al., 1997). Indeed, the divide between parasitism and predation is often blurred, with assaults that begin as acts of piracy escalating into full blown attack whereby the victim is grabbed by the wingtip or tail and forced down into the water and drowned. Solan geese are a favourite target here, but Atlantic puffins and kittiwakes are also pursued, with the former often being killed in the process (Olsen and Larsson, 1997). Being opportunistic scavengers, they also resort to cannibalism when other food sources are scare (Jakubas et al., 2018).
Such revelations, I’ll admit, do not make these birds instantly loveable. They are, however, characterful animals; aggressive, confrontational, and often quite literally in-your-face as they rise to guard their territories from intruders. Such pre-emptive defence extends to human trespassers and follows a choreographed pattern: a running take-off and a menacing, highly direct flightpath culminates in repeated swooping bombardments (sometimes accompanied by faecal spraying), which persist until the intruder has retreated a safe distance. In my experience, these attacks are mostly aimed to miss, but are occasionally poorly calculated. I have read of burst eardrums arising from a misjudged dive colliding with a person's head (Jamie, 2022), and was also told—whilst walking through bonxie territory with the islander who witnessed it—about a fatal heart-attack arising from a hiker's attempted shortcut through an area occupied by the birds.
The morning of our foray into this space was grey and cold. The sky was dark with swept clouds, threatening a weet driv that never quite materialised. A strong wind rushed across the island, whining through fences and overhead wires. Wrapped up to varying degrees against the elements, a group of five figures, me included, gathered by one of the several entrances to North Hill. With the zoonotic threat of HPAI in mind, we don thorough but makeshift personal protective equipment (PPE). Alongside this hastily adapted protective gear, we are each given litter pickers and bin bags, which flap violently in the wind. The pickers are long sticks with soft grips at one end that mould to the shape of the hand. The squeeze of a trigger opens and closes a claw at the far end. Using these minimises our handling of the carcasses.
Responses to the outbreak amongst the various groups involved in the reserve's management differ. The RSPB for example, is not collecting corpses on most of its reserves due to the risks handling the deceased birds to its workforce (RSPB, n.d.). NatureScot likewise advises against carcass collection, unless they are on sites with high public access or if a large majority of local transmissions seem to be arising from the carcasses themselves. Such is the case here, with the spread of the disease through the bonxie population developing from the birds’ scavenging of their dead colony mates (Banyard et al., 2022). But many of the bodies also lie near the flooded shell holes that cover the hill. Studies have highlighted how low temperature bodies of water act as reservoirs of HPAI, allowing infectious particles to persist for long periods of time after contamination, aiding its spread (Ito et al., 1995; Zhang et al., 2014). As well as this, the virus can survive on soil, nesting material, vegetation, and other organic matter for several weeks (NatureScot, 2023). A sense here emerges of a toxic geography similar to the solan goose colony—one where the waterlogged ground oozes with viral threat and saturates our involvements in this space with anxiety, especially following recent experiences of Covid-19.
But, with all of this weighed up, it has been decided by the island community that in this instance leaving deceased birds exacerbates the disease's spread through the population, and the resolution has been made to collect and dispose of them. The motivations of those involved in doing so, however, are deeply personal. Like for me, they stem from an emotional, empathetic connection to the birds borne out of long habits of watching them, and of witnessing the virus's effects on their populations. For some, a sense of helplessness amidst this deathly atmosphere has driven them to act in the face of risk, as well as a frustration at broader organisational and political inaction in the face of the outbreak (personal communication). For the majority of those present, the desire to reduce the threat of infection to other birds is primary. But this is a careful balancing-act. Intervention might spread disease by transferring the virus around on boots and clothes, as well as disturbing sick birds into flight, depleting their energy and further spreading the disease (NatureScot, 2023).
One of the group has plotted the birds’ territories this season and knows the location of the majority of the corpses, enabling us to minimise disturbance. Following their lead, we set off northwards into the reserve. Uneasy birds rise and wheel around us, or stumble away feigning injury, waving broken wings in our direction to draw us away from their eggs and chicks. A surviving bonxie swoops at us, its flight marked by slow wing beats and brief glides (Figure 3). It utters a mewing sound as it wheels overhead. The bird is brown, and stocky—almost barrel-like, with a short tail, and a thick, muscular boxer's neck. It is the colour of exposed moorland, but streaked with paler chestnut, and with eye-catching white flashes on its wings. I duck as it dives towards us, catching a glimpse of its dark hooked beak and black legs as it swoops overhead.

A surviving bonxie flying over North Hill. Image © the author.
It's not long before we come across the first pair of dead birds. Their eyes have already been scavenged. Each corpse is picked up and dropped into a bin liner held open around a bag hoop. The black plastic, which had been flapping noisily in the wind immediately quietens as it fills with the weight of the birds. We set off again, heading for the summit of Errival. As we reach the top I spot a crumpled object in the near distance. It is another corpse; its wings are contorted, and its head is pushed into the soft ground. It looks to have died horribly. Multiple reports of bonxies suffering from HPAI corroborate this, listing symptoms including head tilt, an inability to fly, incoordination, and a violent twitching of the wings and head as the birds become moribund (Banyard et al., 2022). I reach down with my litter picker and grasp the body around the neck, but as I try to lift it, the bird slips through the grip and drops back to the earth. Feathers plucked by the claw billow away in the wind. I nudge the carcass, trying to find a better angle to get a hold. Finally managing to lift it, I find the body is much heavier than I’d expected. I look up and see a binbag bearer heading towards me and lift the bird to deposit it through the hoop. It does not go in easily. Its wings catch on the edge, and the beak pierces the liner, tearing a long gash down the side.
More birds are added; one is packed with maggots that spill out as I raise it, causing me to retch involuntarily. Once the sixth one goes in the binbag is full. It is removed from the hoop, tied closed and placed in a thicker sack, which is then tied and sealed with tape. They will eventually be buried safely by a local farmer. A new bin liner is attached to the hoop. Someone hefts the sack full of dead birds and we set off again, descending the hill to the west. By the time we reach the coast path on the far side, four more bonxies have been added to the sack, as well as a smaller black-headed gull. We head north, following the coastal path. The bags of dead bonxies continue to fill and multiply, and silence begins to dominate. By the time we have navigated around the top of the island and turned for home each of us carries a full bag, with more birds left out on the hill to collect another day. There are close to 30, nearly a third of the island's breeding population. Alongside these observed mortalities, many more will result. If just one parent bird dies the chances of the remaining member of the pair successfully raising a chick are severely reduced (Banyard et al., 2022), often resulting in infanticide and cannibalism (RSPB, personal communication). The islands off northern Scotland are estimated to hold 60% of the world's population of these birds, and there are worries that the loss of such a large proportion may impact future generations (Banyard et al., 2022).
Near to the end of our circuit, the others stop and call me over, pointing at something on the ground. It is a dead chick, caught against a tussock. It is strange looking: patchily moulting its first feathers, with dark, fluffy down still visible protruding through the first quills. It takes me a while to work out what it is—an oystercatcher. I am confused as to why they want to show it to me. I reach for the unmoving chick with my litter picker, only to be held back. The chick, it turns out, is only feigning death, an instinctive response that enables them to avoid being eaten by predators. This is an unremarkable deception compared to other animal pretences that manipulate social situations to the deceiver's advantage, usually when there is a potential food source involved (Despret, 2016). It is a common ruse amongst the birds that hatch on the reserve and is usually easily seen-through, but this time I have fallen for it. The afternoon's work has left me expecting death, making such a display utterly convincing.
Vulnerability and care amidst the viral cloud
The next day is equally overcast, and a mist-like rain known in parts of Orkney as a muggero-fue settles over the island. The mizzle is thick and enveloping, and the island's neighbours Westray, Eday, and distant Sanday, recede from view. Feeling disturbed by the previous day's events I pull on my raincoat and step out into the damp, hoping movement will help me process recent events. Following the path through the marshy, orchid-filled field, a curlew—upset by my presence—lifts its head and utters a cry; a haunting, evocative, repeated ‘crooo-ee, crooo-ee.’ Invisible in the mist overhead, a snipe is drumming, diving obliquely with its tail fanned so the air rushes through it, making a vibrating noise. The sound is eerie, deepened by the atmospheric conditions, a mysterious ‘huhuhuhuhu.’ I drop down onto the shore and proceed southwards towards the village. As I walk I see the corpses of birds everywhere. Solan geese jut from the sand, half buried. Sometimes just the tops of their heads protrude, the feathers moving slightly as the wind scrapes the ground, rippling the beach away into streams of matter. Still other birds lie discarded amongst the seaweed on the tideline: guillemots, fulmars, gulls. Tattered feathers are strewn throughout. The mortality is devastating.
Turning attention to how the atmospheres of the viral cloud envelop allows one to apprehend something of their affective spacetimes. Experience of these global and genetic drifts of mutant spectra is always haunted by the way that they exceed the immediate. The sensing of an atmosphere is always partial, it is never fully visible to something immersed within it. Like Morton's (2013; see McCormack, 2018) hyperobjects, they evoke something withdrawn, or ungraspable that evades complete understanding. Such elusiveness makes viral/atmospheric assemblages ideal for understanding something of the vast, diffuse nature of this global outbreak of HPAI and its slow, drifting movements along migration routes and between avian populations. However, it is through envelopment that these abstract assemblages are made palpable. They are apprehended through local experience, though this limits visibility. Enclosed in the viral cloud in Orkney I see and feel the corporeal effects of the virus on wild bird populations here, including emotionally, but lose sight of broader scales and injustices—the deaths of multitudes of wild birds, poultry, and now mammals elsewhere. 6
Envelopment also focuses my attention on vulnerability. To be shrouded in this and similar viral/atmospheric assemblages is to be blanketed in a contact zone where I am brushed unwantedly and invisibly by exposure to both the virus itself, and the swirling clouds of affectivity caused by its local effects. To be enveloped is to be touched, with such interactions acting like felt points of contact. But the haptic interactions at work here are not physical ones, they are indications of a more fundamental all-encompassing immersion in circulations of affect emergent from multispecies relations, the influence of which ‘is there all the time’ (Puig de la Bellacasa, 2017: 99). For Ahmed (2014), a key constituent of this amorphous, invisible sensory milieu is an emotional sociality that circulates between bodies and is shaped by contact between them. It is my embodied immersion in these precipitations of affect that enables the virus to disturb my being without infecting it. Infection is not needed to feel undone by proximity to HPAI. Envelopment is transformative. My emotions are an expression of the touch of the virus as it reshapes the boundaries between the individual and the collective.
But this focus on my own emotions in this context troubles me. At the heart of this situation is not a shared experience of exposure, but an unequal one. Despite the potential risk of human infection, the birds are much more susceptible to the virus, and more vulnerable to it. I may feel devastated by witnessing its consequences locally, but that means little to the thousands of birds dying whilst engulfed in a viral cloud of anthropogenic origins. In several works exploring the subject of vulnerability, Butler (2009, 2004) has incorporated this disparity into an ethico-political theory. For Butler (2004: 28) life is, at its heart, relational; we become attached to others, ‘given over, beyond ourselves, implicated in lives that are not our own.’ The individual is always open to this fundamental sociality, and is thus exposed—becoming vulnerable; emotionally, existentially, or otherwise. This vulnerability precedes the formation of the ‘I’ and cannot be avoided. Such interdependence, and the vulnerability it engenders, forms a complex political order, simultaneously opening us to hurt, but also producing an ethical responsibility towards the other (Butler, 2004; see Puig de la Bellacasa, 2017).
Vulnerability, then, is something held in common, even if it is experienced differently. But despite this mutual precarity, the origins of HPAI still prick at my conscience. The emotion I feel is shaped as much by my participation (however remotely) in the systems that have caused its emergence as it is by my sensing of the virus's consequences for those more vulnerable, and my receptive orientation towards them and their suffering. For Butler (2004), acknowledging shared vulnerability acts as a mode of resistance, engaging with that complicity by drawing us into connection with those whose lives are perceived to matter less. Though the violences of the viral cloud are felt differently across the lifeworlds they engulf, through attending to this ‘shared-but-different vulnerability’ (Green and Ginn, 2014: 152) empathy develops. To be moved by, and thus care about the plight of ecological others is an example of ethics—it is a way of reaching towards another being, and its vulnerability, even if we are complicit in its precarity (Green and Ginn, 2014; see Haraway, 2008). What is important here is the way that personal practices connect the individual to the collective (Puig de la Bellacasa, 2017). By exposing ourselves, making ourselves vulnerable—either emotionally, or risking infection—we engage in an ethical practice by sharing in the vulnerability felt by others. As Ahmed (2014) argues, emotions are relational. To feel such emotion indicates an attentiveness towards the experience of others. To respond to those emotions carefully reshapes one's orientation towards the more-than-human crowd. Emotions matter for politics, including biopolitics. Feelings open bodies to others. They have the power to shape emergent worlds.
Here, the corporeal concerns of carcass collection offer a way of altering the atmospheric milieu; of pushing back against the viral cloud and shifting the mood. It is an act of care that seeks to respond to the effects of a distant uncaring—the vast violences of industrial poultry production. In her work on the significance of care for thinking and living in more-than-human worlds, Puig de la Bellacasa (2017) explores the problems, possibilities, and ambiguities of touch as an involved form of caring relation. As she highlights, touch often indicates concern. It removes the distance and detachment between individuals. Though, as we have seen, touch in the context of the viral cloud is problematic and risks spreading the disease, or even causing infection. Touch, here, involves cohabiting with risk; it ‘makes us confront the fact of our own mortality,’ as well as ‘our need for each other’ (Dumm, 2008: 156; see Puig de la Bellacasa, 2017). Despite the shattering emotion involved in carcass collection, such acts of care provide a way of responding to the situation. It is an act of empathy that reaches outwards into the surrounding more-than-human sociality and seeks to makes an intervention. The deaths caused by these viral outbreaks will not be total, though these seabird populations are also suffering from many other stresses. Pockets of resistance to the disease within populations will eventually develop into a sort of immunity. Any action that might reduce its spread makes the conditions for living with such outbreaks easier as this immunity increases.
As I write reports come in detailing the further spread of the outbreak, including amongst dense populations of breeding birds in Antarctica, populations which have never been exposed to the virus before (Weston, 2023b). These talk variously of the emotional toll of this care work—of collecting corpses—on those in proximity to these populations who care for these birds, but also of how resistance is slowly emerging. Studies of returning gannets on Bass Rock in the Firth of Forth have noted a strange phenomenon. The irises of a proportion of the birds have changed colour. Previously startlingly blue, they have now turned black. Samples taken from these birds have revealed HPAI antibodies, indicating they had survived previous infection (Lane et al., 2024). In the presence of these viral circulations, this is, for the time being, a darker present; one marked by mass deaths. But as this article has shown, attending to the sociality of emotions offers a way of navigating the affective turbulences brought about by of mass mortality, highlighting how a reorientation towards the more-than-human along more careful grounds is possible.
Highlights
Offers a key addendum to understandings of HPAI, emphasising its various corporeal consequences.
Synthesises something of the global drift of these viral clouds and how they intersect with place.
Explores how emotion might be drawn into understandings of multispecies relationality
Provides an example of more-than-human research practice.
Footnotes
Acknowledgements
I would like to thank Merle Patchett and Franklin Ginn for providing feedback on various iterations of this manuscript, as well as their wider support. Thanks also to Alice Would, John Morgan and Mark Paterson for their readings, and to Katie Nudd and Krithika Srinivasan for their patience and editorial support. I am grateful to my friends on and associated with Papay for their warm welcome, and for sharing their company, insights and homes with me over the course of my multiple stays on the island. I would also like to acknowledge the feedback of the two anonymous reviewers who helped improve this article.
Ethical approval
The School of Geographical Sciences Research Ethics Committee at the University of Bristol approved this research (Approval: 11299) on 10th May 2022.
Funding
The author disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This work was supported by an AHRC (Arts and Humanities Research Council) doctoral studentship and an additional research grant from the South, West and Wales Doctoral Training Partnership.