
Abstract
Mast cells (MCs) are known to regulate innate and adaptive immunity. MC activators have recently been described as safe and effective vaccine adjuvants. Many currently known MC activators are inadequate for in vivo applications, however, and research on identifying novel MC activators is limited. In this study, we identified novel MC activators by using high-throughput screening (HTS) assays using approximately 55,000 small molecules. Data sets obtained by the primary HTS assays were statistically evaluated using quality control rules and the B-score calculation, and compounds with B-scores of >3.0 were chosen as mast cell activators (hits). These hits were re-evaluated with secondary and tertiary HTS assays, followed by further statistical analysis. From these hits, we selected 15 compounds that caused degranulation in murine and human MCs, with potential for flexible chemical modification for further study. Among these 15 compounds, ST101036, ST029248, and ST026567 exhibited higher degranulation potency than other hit compounds in both human and mouse MCs. In addition, the 15 compounds identified promote de novo synthesis of cytokines and induce the release of eicosanoids from human and mouse MCs. HTS enabled us to identify small-molecule MC activators with unique properties that may be useful as vaccine adjuvants.
Introduction
Mast cells (MCs) are innate immune cells highly prevalent in the dermal region of the skin and in the respiratory, gastrointestinal, and urinary tract mucosa. 1 The strategic location at the host–environment interface and the expression of a wide range of pattern recognition receptors (PRRs) allow MCs to serve as immune surveillance cells, and these cells play a critical host-protective role as a first line of defense through innate immune responses. In addition, the expression of receptors on MCs recognizing activated complement, endogenous peptides (antimicrobial peptides, neuropeptides, etc.), exogenous venom peptides, and interleukin-33 (IL33) can amplify proinflammatory responses and modulate immune responses.1–5 The immune regulatory role of MCs is particularly supported by the fact that MCs have a capacity to pre-store proteases, cytokines, biogenic amines, and other mediators that can be instantly released at the recognition of trigger stimuli. In addition to pre-stored mediators, MCs produce and secrete a wide range of cytokines and chemokines through de novo synthesis. These cytokines and chemokines associated with MC activation are characteristic of a T-helper type 2 (Th2) response, and they may provide protection against extracellular infection.6–8 We recently demonstrated that intentional activation of MCs at local tissue sites could induce MCs to release pre-stored inflammatory mediators, resulting in the upregulation of adaptive immune responses such as the production of protective antibodies.9–12 The ability of MC activators to serve as vaccine adjuvants has been confirmed by other laboratories.13–15 MC activation can be achieved by various compounds such as Compound 48/80 (C48/80), and peptides such as mastoparan and β-defensin.3,5,6 These stimulators may act via triggering the Mas-related G-coupled receptor X2 (MRGPRX2), one of the major receptors that recognize these stimulators.16–18
In this study, we identified novel MC activators using high-throughput screening (HTS) of 55,198 compounds using MC degranulation as the readout. Primary screening using the mouse MC line MC/9 identified 1265 compounds with MC degranulation activity (2.29% hit rate). Secondary screening with a human MC line (ROSA) allowed downselection to 194 compounds exhibiting both mouse and human MC activation activity. Further downselection was accomplished through tertiary screening with primary human MCs. The 15 compounds selected from tertiary screen hits were then evaluated for their ability to induce MC secretion of select cytokines and arachidonic acid metabolites.
Materials and Methods
Compounds
Compound libraries were purchased from TimTec (TimTec LLC, Newark, DE). Compound libraries purchased included the ActiGlobe-50K library, a natural product library of 720 compounds (NPL-720), a cationic amphiphilic druglike library, compounds that matched the National Institutes of Health (NIH) clinical collection library, and cationic amphiphilic-known drug molecules. All 15 hit compounds are identified by their chemical name and the library name: 1-[6-(4-ethylphenyl)-4-(trifluoromethyl)pyrimidin-2-yl]-4-piperidylamine (ST101036), 2-(tert-butyl)-4-[5-(tert-butyl)-4-hydroxy-2-methylphenylthio]-5-methylphenol (ST029248), 4-ethyl-5-methoxy-2-(5-methyl-4-phenoxy-1H-pyrazol-3-yl)phenol (R529877), 5-chloro-10-[3-(trifluoromethyl)phenyl]-9-azatetracyclo[10.2.1.0<2,11>.0<3,8>]pentadeca-3,5,7-triene-7-carboxylic acid (ST086136), 2-(5-methyl-4-phenoxy-1H-pyrazol-3-yl)-5-propoxyphenol (R606278), 2,4-bis(1,1-dimethylpropyl)phenol (ST081379), 6-chloro-4-phenyl-2-quinolylamine (ST048871), 2-[(ethylsulfonyl)methyl]-5-nitrofuran (R127655), 4-(4-chlorophenoxy)-3-(trifluoromethyl)phenylamine (ST027688), 3-hydroxy-2-(1-methylpyrazol-4-yl)chromen-4-one (ST099914), (2E)-3-(4-hydroxyphenyl)-2-(6,7,8,9-tetrahydro-5H-[1,2,4]triazolo[4,3-a]azepin-3-yl)-2-propenenitrile (L147192), 1-(5-chloro-2-hydroxyphenyl)-3-phenylpropane-1,3-dione (ST026567), 5-[(4-hydroxyphenyl)methylene]-2-morpholin-4-yl-1,3-thiazolin-4-one (ST045940), 4-nitro-2-{[(1-phenyl-1H-tetraazol-5-yl)sulfanyl]methyl}phenol (L201863), and 4-(1,1-dimethylhexyl)phenol (ST029279) (the Small Molecule Synthesis Facility at Duke University, Durham, NC) were stored as 20 mM stocks in DMSO. All compounds were used at the indicated concentration.
Cell Lines and Cell Culture
Human and murine MC lines were used in this study: MC/9 [American Type Culture Collection (ATCC), Manassas, VA] is a murine cell line. ROSA and leukocyte adhesion deficiency-2 (LAD2) are human MC lines. MC/9 cells were cultured in Dulbecco’s Modified Eagle Medium (DMEM; Quality Biological, Gaithersburg, MD) containing 10% heat-inactivated fetal bovine serum (HI-FBS; GenClone, Genesee Scientific, San Diego, CA), 10% rat T-Stim supplement (Corning, Corning, NY), 100 U/mL penicillin, 0.1 mg/mL streptomycin, 25 mM 4-(2-hydroxyethyl)-1-piperazineethanesulfonic acid (HEPES), 2 mM L-glutamine, 1 mM sodium pyruvate, 1 mM nonessential amino acids, and 1 mM Minimum Essential Medium (MEM) amino acids (all from Quality Biological). ROSA cells (a kind gift of Dr. Michel Arock, Molecular Oncology and Pharmacology, Ecole Normale Supérieure de Cachan, Cachan, France) were cultured in Iscove’s Modified Dulbecco’s Medium (IMDM; Quality Biological) containing 10% HI-FBS, 100 ng/mL recombinant human stem cell factor (rhSCF; Biolegend, San Diego, CA), 2 mM L-glutamine, 100 U/mL penicillin, 0.1 mg/mL streptomycin, 1× MEM vitamin solution, 40 µg/mL human transferrin (MyBiosource, San Diego, CA), 10 µg/mL insulin (MyBiosource), and 75 µM β-mercaptoethanol (Sigma-Aldrich, St. Louis, MO). LAD2 cells (a kind gift of Dr. Dean Metcalfe, Laboratory of Allergic Disease, NIH, Bethesda, MD) were cultured in Stem Pro 32 media including supplements (Invitrogen, Thermo Fisher Scientific, Waltham, MA) with 80 ng/mL rhSCF, 100 U/mL penicillin, 100 µg/mL streptomycin, and 2 mM L-glutamine. All cells were cultured at 37 °C in a humidified water-jacketed incubator under 5% CO2/95% air atmosphere.
Generation of Primary Human MCs
Primary human MCs were differentiated from human CD34+ hematopoietic stem cells. 19 Human CD34+ hematopoietic stem cells were obtained from commercial sources (AllCells, Alameda, CA). Isolated CD34+ cells were suspended at 5 × 105 cells/mL in IMDM supplemented with 100 ng/mL rhSCF, 50 ng/mL recombinant human interleukin-6 (rhIL6), 1 ng/mL rhIL3 (all from Biolegend), 100 U/mL penicillin, and 0.1 mg/mL streptomycin, and they were cultured for 3 weeks, changing media weekly. 19 In weeks 3–6, the same culture medium was used except that rhIL3 was omitted. 19 In weeks 6–7, 10% fetal bovine serum was added to the medium, and matured primary human MCs were used in experiments at week 7. The maturation of primary human MCs was confirmed by evaluating granularity (toluidine blue staining) and receptor expression (flow cytometry on FcεRI and CD117 expression).
β-Hexosaminidase Assays
To measure the degranulation of MCs, the release of β-hexosaminidase was evaluated as previously described, with minor modifications. 20 MCs were incubated with compounds in Tyrode’s buffer (2 mM MgCl2, 5 mM KCl, 1.8 mM CaCl2, 135 mM NaCl, 5.6 mM glucose, 1 M HEPES, and pH 7.4) for 30 min at 37 °C. To obtain the value of maximum release of β-hexosaminidase from MCs, cells were treated with 0.1% Triton X-100, and the negative control was treated with vehicle. After 30 min of incubation, supernatants were collected after cells were spun down. The collected supernatants were mixed with 3.4 mg/mL 4-nitrophenyl N-acetyl-β-D-glucosaminide in citrate buffer (0.1 M sodium citrate and pH 4.5) at 37 °C to measure β-hexosaminidase activity. After 60 min, the reaction was stopped by mixing with 0.1 M carbonate buffer (pH 10), and the optical density was measured at 405 nm. The formula to assess the degree of MC degranulation is as follows:

Cytokine Multiplex Assay
To evaluate cytokine production from MCs, mouse bone marrow–derived mast cells (BMMCs) or human LAD2 cells were used. MCs were cultured at 2 × 105 cells/well (96-well plate) in culture media and treated with the 15 hit compounds (12.5 µM) or positive controls [mastoparan 7 (12.5 µM) or ionomycin + phorbol myristate acetate (PMA) (20.4 nM + 352.5 nM, respectively)]. After 20 h of incubation at 37 °C in a humidified water-jacketed incubator under 5% CO2/95% air atmosphere, cell-free supernatant was collected and tested for the presence of cytokines using the Bead-Based Multiplex Assay (EMD Millipore, Burlington, MA). In this assay, 50 µL of culture supernatant was mixed with a panel of beads that are covalently bound to an antibody that recognizes one of the following cytokines: human: chemokine (CC motif) ligand-2 {CCL2 [also called monocyte chemoattractant protein-1 (MCP1)]}, CCL3 [macrophage inflammatory protein-1α (MIP1α)], CCL4 (MIP1β), CCL5 (RANTES), chemokine (CXC motif) ligand-8 [CXCL8 (IL8)], basic fibroblast growth factor (FGF2), granulocyte colony-stimulating factor (G-CSF), granulocyte–macrophage colony-stimulating factor (GM-CSF), interferon-γ (IFNγ), IL1α, IL1 receptor agonist (IL1ra), IL1β, IL2, IL4, IL5, IL6, IL10, IL17, tumor necrosis factor-α (TNFα), and vascular endothelial growth factor (VEGF); and mouse: GM-CSF, IFNγ, IL1β, IL2, IL4, IL5, IL6, IL10, IL12 p70, IL17, keratinocyte-derived cytokine (KC), MCP1, MIP2, TNFα, and VEGF.
Eicosanoid Synthesis
To measure the release of eicosanoids from activated MCs, prostaglandin D2 (PGD2) and leukotriene C4 (LTC4) were measured. Mouse BMMCs or human LAD2 cells were cultured at 2 × 105 cells/well (96-well plate). MC activation occurred in culture media for the LTC4 assay (Cayman Chemical, Ann Arbor, MI) or in Tyrode’s buffer for the PGD2 assay (Cayman Chemical). MCs were treated with the 15 hit compounds (50 µM) or the positive controls [mastoparan 7 (12.5 µM) or ionomycin/PMA (20.4 nM/352.5 nM)]. After 2 h of incubation at 37 °C in a humidified, water-jacketed incubator under 5% CO2/95% air atmosphere, cell-free supernatant was collected and tested for the presence of LTC4 or PGD2.
B-Score, C-Score, and Z-Score Normalization
The normalization algorithm is described in detail elsewhere. 21
Results and Discussion
Assessment of Variability in the Screening Assay
An MC degranulation assay was evaluated for its ability to be used as an HTS assay to identify compounds that induce MC degranulation. MCs store presynthesized mediators in their granules, including β-hexosaminidase, which has lysosomal enzymatic activity. 22 On MC stimulation, β-hexosaminidase can be released instantly, and its enzymatic activity is stably retained, making β-hexosaminidase release a useful indicator of MC degranulation. 20 We performed HTS of commercially available compound libraries using an in vitro β-hexosaminidase release assay using the MC/9 line of murine MCs. The assay procedure used 384-well plates and automated liquid-handling systems to minimize bias and enhance reproducibility. These adaptations significantly reduced the potential for errors that are common with manual handling of cell culture reagents and compound libraries. The readout in these assays is the release of β-hexosaminidase, which can be detected by its ability to produce a colorimetric change [in optical density (OD)] due to its activity on its substrate, 4-nitrophenyl N-acetyl-β-D-glucosaminide. 20
Triton X-100 was used as a positive control to lyse the MCs to measure the maximum release of β-hexosaminidase. Tyrode’s buffer was used as the negative control. Intraplate variation was evaluated by measuring triplicates of negative and positive control wells in the 384-well plates. Interplate variation was evaluated by measuring the distribution of responses among different plates for the same concentration of control-treated wells. Figure 1A shows significant interplate variation of OD values from the Triton X-100 of 11 different 384-well assays performed on different days (batches). Noticeable batch effects were associated with MC/9 cell status and stability of the treatment reagent, necessitating the use of a batch normalization procedure.
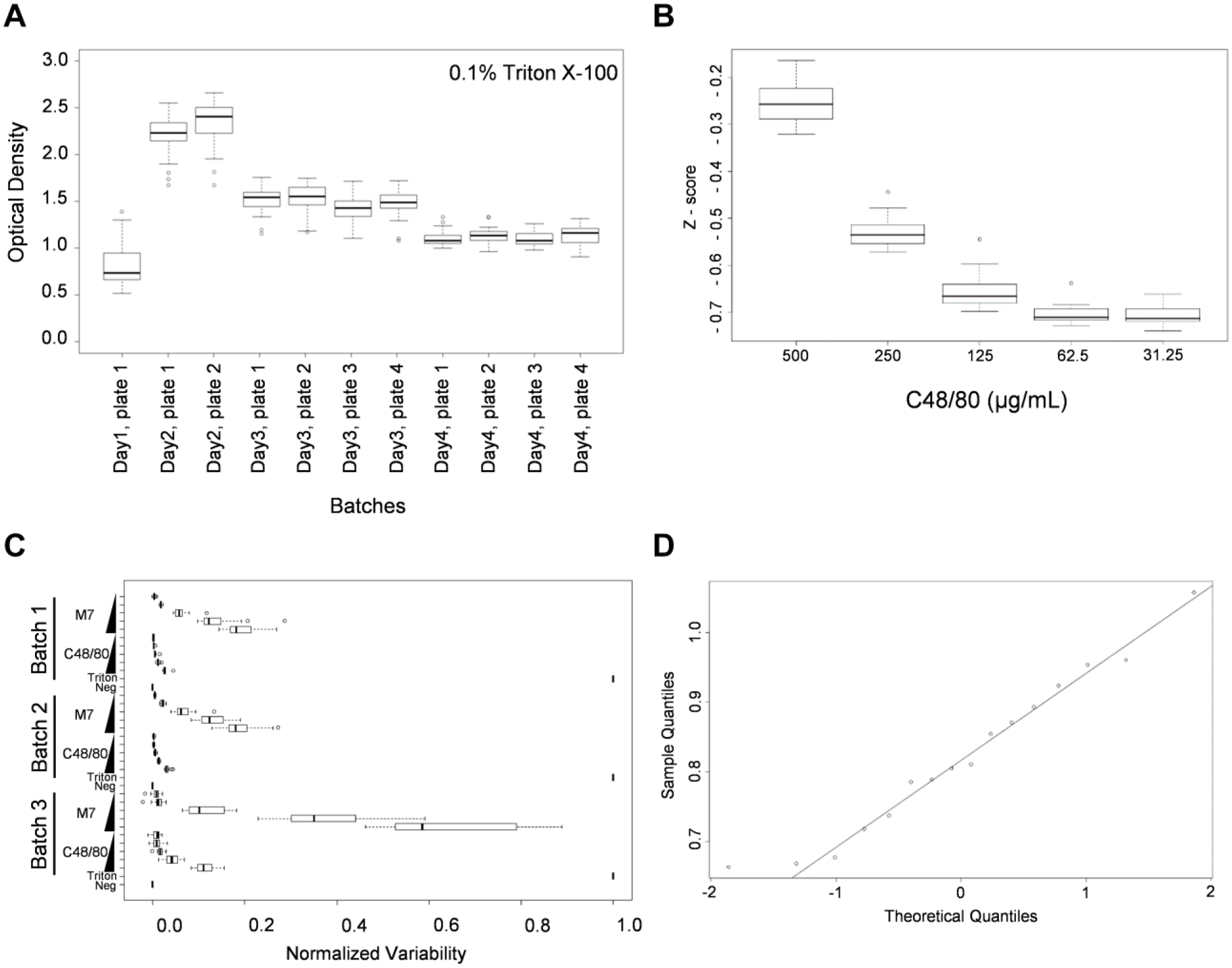
Variability evaluation by using a pilot screening assay. (
Next, we evaluated the MC degranulation responses at various concentrations of C48/80 (a well-known MC activator) to determine if the assay would generate the expected dose–response curve ( Fig. 1B ). Although the noise from the assays is expected to be constant, we noticed that the magnitude of response variance was proportional to the mean degranulation response. To evaluate whether the observed heteroscedasticity of the MC response is C48/80 specific, mastoparan-7 (M7, another well-known potent MC activator) was tested for MC degranulation at various concentrations. Figure 1C demonstrated that M7 also showed the heteroscedasticity of the MC degranulation response, consistent with the C48/80 response. Finally, we investigated the normality of treatment-specific variability to determine if the variability of each treatment replicate was approximately normally distributed. Based on the quantile–quantile (QQ) plot ( Fig. 1D ), we noticed right skewing, which suggests that assumptions of normality are violated and that a traditional Z-score measure would need care in interpretation; therefore, an alternate statistical method was used, as explained below.
B-Score Provides Robust Corrections for Variation in OD Values
Due to the observed data variance in the positive control experiments using 384-well plates for the HTS assays, the following experimental design considerations were developed for robust screening data analysis. First, to minimize the positional edge effects resulting in intraplate variation, the positive and negative controls were loaded into alternating wells in the first two and last two columns of each assay plate ( Fig. 2A ). Second, to reduce the degree of interplate variation, triplicates of each tested compound were plated among three 384-well plates. Third, we used the B-score normalized values to calculate the values to be used for compound downselection. The B-score is well-known to be a more robust normalization method for HTS screening experiments. B-score values are calculated using a median polish procedure, as described in Figure 2B . The B-score, which does not use controls, robustly corrects for both positional (via median polish) and interplate effects (via median absolute deviation scaling), and it does not make any parametric assumptions. In addition, the B-score is resistant to outliers and minimizes measurement bias. 23 Therefore, B-score normalized values provided the clearest distinction between potential hits. The use of control-based normalization [(test value − negative control value) / (positive control value − negative control value)], which we shall refer to as the C-score, and the Z-score methods have the disadvantages of assuming similar cross-compound variances and being sensitive to outliers. 23 Moreover, C-scores, which depend on positive and negative controls, are also sensitive to edge effects, since positive and negative controls are loaded in the left and right boundary wells. 23 Given these considerations, we used the B-score method to normalize the data and select hits from primary screening with MC/9 cells. While not used for determination of hits, that percentage of degranulation as measured by the C-score is still used for reporting due to its ease of interpretation.
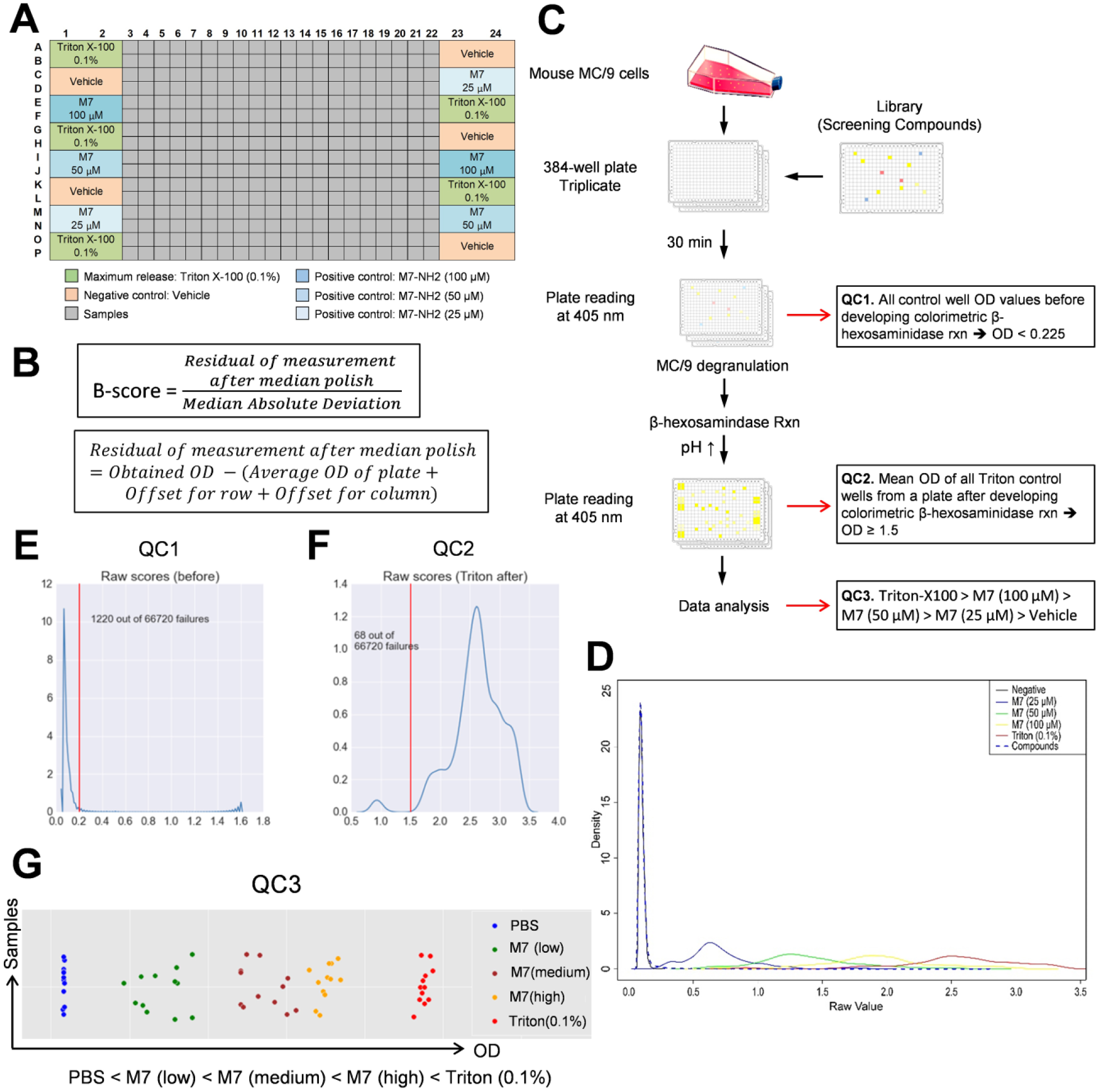
High-throughput screening (HTS) to identify mast cell (MC)-activating compounds. (
Quality Control (QC) Rules for Establishing Robust HTS
We performed the HTS degranulation assays as schematically described in Figure 2C . The assay raw data results were displayed as a histogram graph ( Fig. 2D ). When the assay raw data for the compounds screened was graphed with the raw data from assay positive and negative controls, the compound raw data values were distributed within the range of assay responses observed with the negative control [phosphate-buffered saline (PBS) treated] to positive control (Triton X-100 0.1%). In addition, different levels of degranulation could be demonstrated by the responses observed after treatment with low, medium, and high doses of M7. Due to technical or reagent inconsistencies and biological variability, however, HTS assays may provide variable results throughout the course of the entire time needed to complete screening of many compounds. Therefore, QC procedures are required to minimize these effects and provide reproducible assay results throughout the course of the entire screening procedures. We developed QC rules to minimize false-positive hits during HTS. We noticed that a number of the compounds exhibited intrinsic color in solution and inflated the colorimetric readout of the β-hexosaminidase release assay. QC rule 1 (QC1) functions to exclude those colored compounds from being identified as hits by passing only noncolored compounds based on the OD readout before addition of carbonate buffer (developer for the colorimetric β-hexosaminidase reaction). Based on the pilot assays performed using the positive and negative controls, which are intrinsically noncolored compounds, a threshold OD value of 0.225 was set and validated using all MC/9 assay data ( Fig. 2E ).
QC rule 2 (QC2) controls for the sensitivity of the β-hexosaminidase assay readout based on OD values of Triton X-100–treated wells. The Triton X-100 control serves as a positive control (maximum release of β-hexosaminidase) for detection of degranulation in the assay because close to 100% cell lysis should be achieved in the well, thus releasing 100% of β-hexosaminidase. This control allows identification of the upper limit of detection in the assay. Based on the results from pilot assays, an average OD value of ≤1.8 was expected from Triton X-100 control wells. Following completion of MC/9 screening assays, however, this threshold was revised to 1.5 based on distribution of data from all plates ( Fig. 2F ). Final analysis and scoring of MC/9 compound “hits” were performed using this revised threshold.
QC rule 3 (QC3) assures appropriate sensitivity of the assay based on values from control-treated wells. In each assay plate, three doses of M7 were used as a dose-dependent positive control in addition to the Triton X-100 control. These controls were used to assess the sensitivity of the assay by ensuring that a dose response was observed in correlation with Triton X-100 and the three M7 dilutions. The high dose of M7 was not expected to result in activation of 100% of the cells and should therefore have a lower OD value than that of the Triton X-100 control. PBS-only containing wells serve as a negative control to set the minimum level of detection in the assay. Therefore, the mean OD value after carbonate addition for each M7- or Triton X-100–treated well should rank in the following order: Triton-X100 > M7 (100 µM) > M7 (50 µM) > M7 (25 µM) > PBS only. Plates that failed to show the expected dose–response rankings were rejected. Ranking of assay control OD values from three representative plates is shown in Figure 2G .
These three QC rules enabled us to establish the robustness of assay performance, which ensured that assay performance was acceptable and reproducible prior to data analysis to identify compounds with MC-degranulating activity.
Primary HTS Using Murine MC/9 Cells
Various compound sources were used for the HTS, as shown in Table 1 . ActiGlobe-50K was the largest library used and comprised 51,040 compounds. We also included the cationic amphiphilic (CA) drug–like set library, because it includes known MC activators and other compounds exhibiting a cationic charge and ligands of MRGPRX2 such as C48/80. 17 For the primary HTS assay, all 55,198 compounds from the listed libraries were screened in a degranulation assay using mouse MC/9 cells, as described in Figure 3A . We screened the novel compounds at a concentration of 100 µM, which was based on our previous experience with MC/9 cells and compound screening. 20 The OD values obtained from the primary HTS were evaluated using the three QC rules and normalized using the B-score method described above. An arbitrary B-score threshold of 3 was used to identify compounds with MC-degranulating activity (“hits”) that would move to secondary screening in human MCs. In total, 1265 hits were identified for an overall hit rate of 2.3%. Table 1 shows the number of hits identified in each of the analyzed compound libraries. 1103 compounds from the ActiGlobe-50K library were identified as hits, whereas 68 compounds from the CA drug–like library and 88 compounds from NPL-720 were identified as hits.
List of Compound Libraries Screened for MC-Activating Compounds.
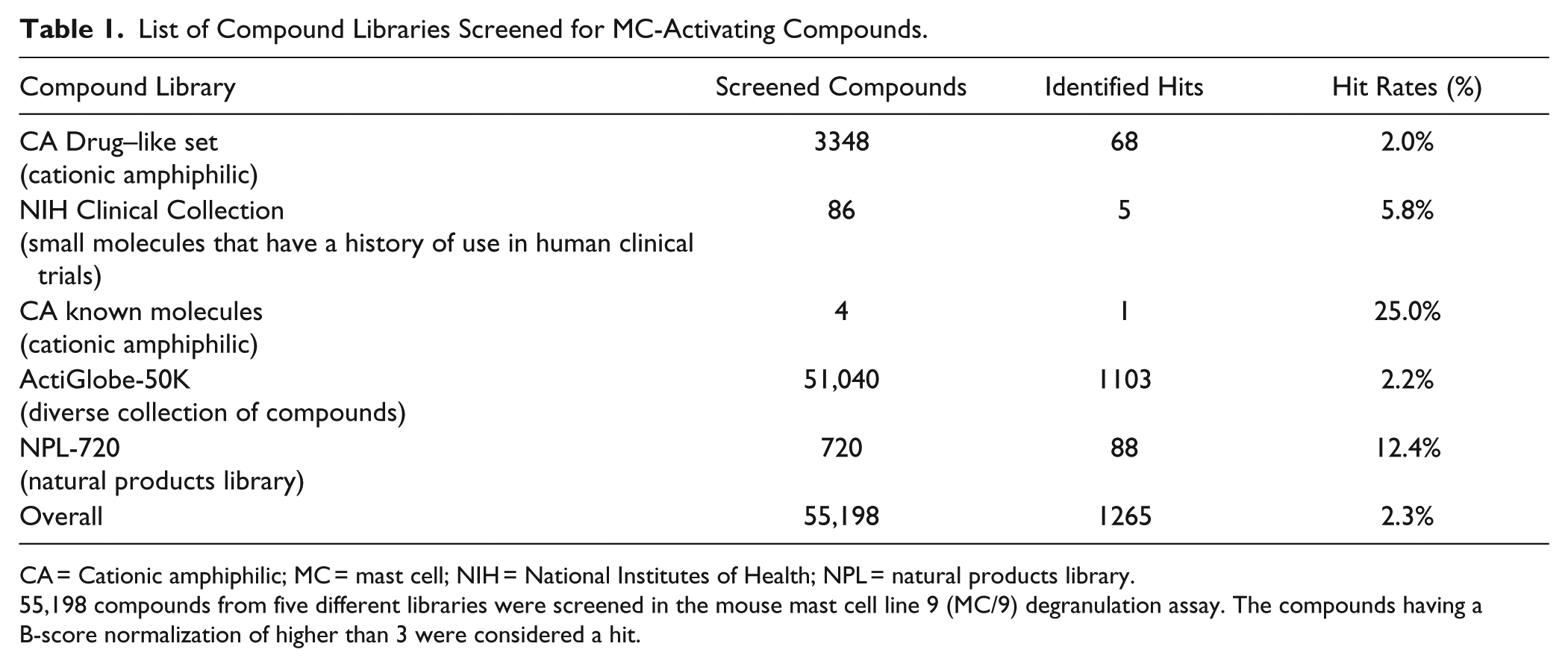
CA = Cationic amphiphilic; MC = mast cell; NIH = National Institutes of Health; NPL = natural products library.
55,198 compounds from five different libraries were screened in the mouse mast cell line 9 (MC/9) degranulation assay. The compounds having a B-score normalization of higher than 3 were considered a hit.
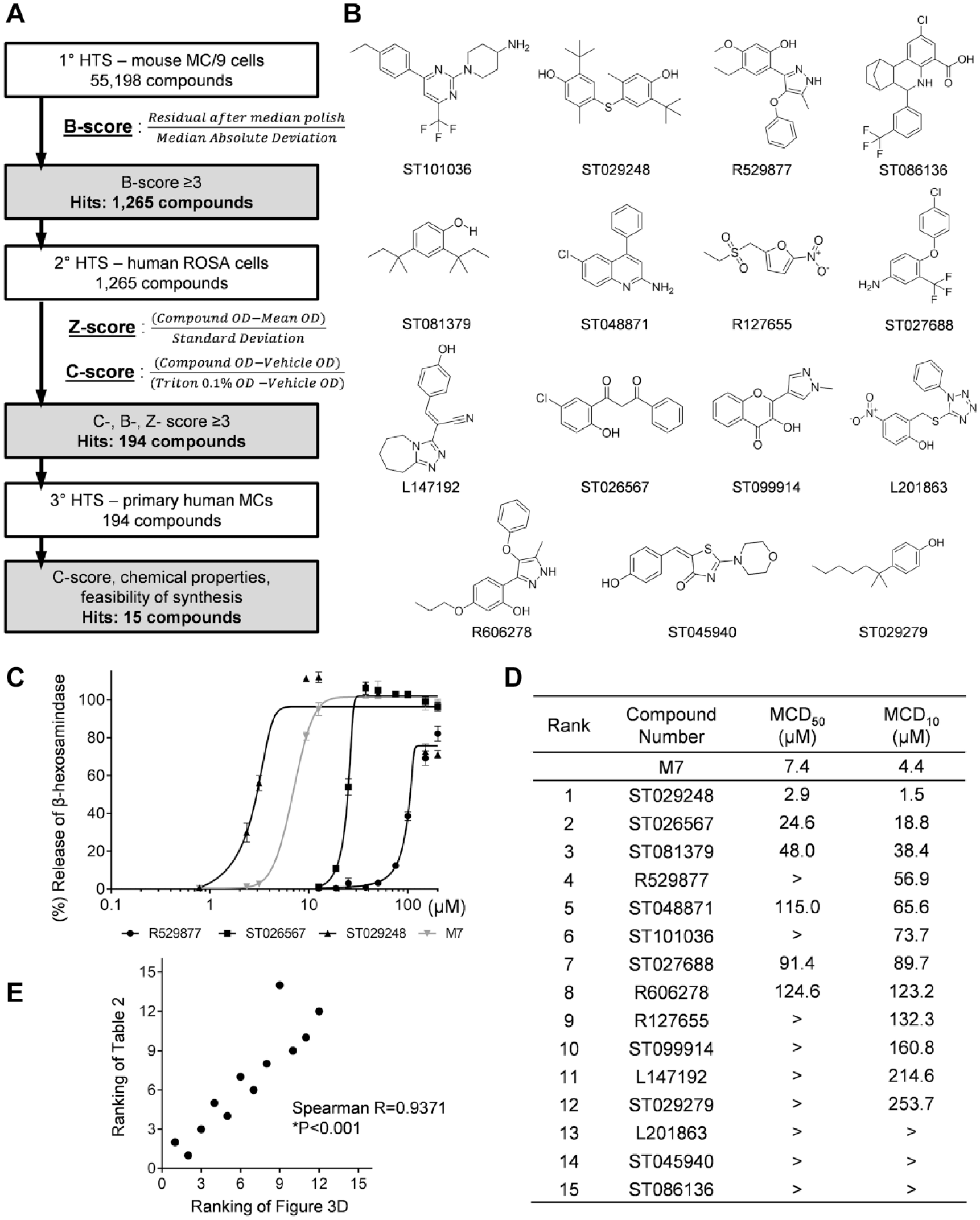
High-throughput screening (HTS) workflow and mast cell degranulation (MCD) parameters for ranking the 15 hit compounds. (
Retesting Hit Compounds on Human MCs
Ultimately, the goal of these studies is to identify compounds that can function as MC-activating adjuvants in the context of human vaccination. Because primary screening of the compounds was performed using a murine MC line, it was necessary to confirm the activity of hit compounds in human MCs. Secondary compound screening was performed using the human ROSA MC line, 24 followed by tertiary screening using primary human MCs. The ROSA cell line was used because it is easy to grow and the cells retain pre-stored MC mediators and are capable of de novo synthesis of cytokines. 24 During secondary screening, hit compounds identified in the primary screen were retested at four concentrations (100, 50, 25, and 12.5 µM) in a ROSA cell degranulation assay to determine dose-dependent responses. For tertiary screening, we used cultured primary MCs, which were matured from the human CD34+ peripheral blood mononuclear cells (PBMCs) of three healthy donors. The tertiary screening was also performed with the hit compounds at four concentrations (100, 50, 25, and 12.5 µM) to determine dose-dependent responses in a primary human MC degranulation assay. Figure 3A summarizes the process of this secondary and tertiary screening that was performed in this study, and it describes the normalization methods that were used for data analysis and compound hit selection following the primary (1°), secondary (2°), and tertiary (3°) screening assays. We obtained 1295 compound hits from the primary HTS and 194 compound hits from the secondary HTS, and we selected 15 compounds ( Fig. 3B ) after confirming the degranulation activity from the tertiary HTS along with consideration of the diversity in chemical properties, ease of chemical synthesis, and feasibility of structure–activity relationship (SAR) generation and medicinal chemistry optimization.
Degranulation of Mouse and Human MCs by 15 Hit Compounds
Three stages of HTS screening allowed us to identify 15 compounds with MC degranulation activity that warranted further investigation. Because the compounds were identified by HTS using compounds in library plates, we confirmed the MC activation activities using new lots of the compounds in primary mouse BMMCs and the human LAD2 cell line. Unlike other human MC line cells, human LAD2 cells are a human MC line similar to primary human MCs in lacking any mutations in KIT and expressing FcεRI along with abundant production of MC mediators. 25 The drawback, however, of the slow growth rate (~2 weeks for doubling) hampered direct application of this cell line for HTS in this experiment. Table 2 shows the degranulation response of mouse BMMCs treated with the 12 hit compounds showing more than 10% degranulation at 100 µM. The order of individual compounds was determined based on the level of degranulation response to 100 µM concentration, in which ST101036, ST081379, R529877, and ST029248 exhibited more than 70% degranulation. The other compounds elicited a wide range of degranulation from 60% to 1%. Unexpectedly, ST086136 and ST027688 induced poor degranulation (less than 5%) of mouse BMMCs. We speculate that primary mouse BMMCs are less responsive than MC/9 cells to these compounds, because these compounds induced more than 10% degranulation activity during HTS using the MC/9 line (data not shown), along with diverse chemical properties.
Rank Summary of the 15 Hit Compounds Evaluated by Mouse and Human MC Degranulation Assays. (
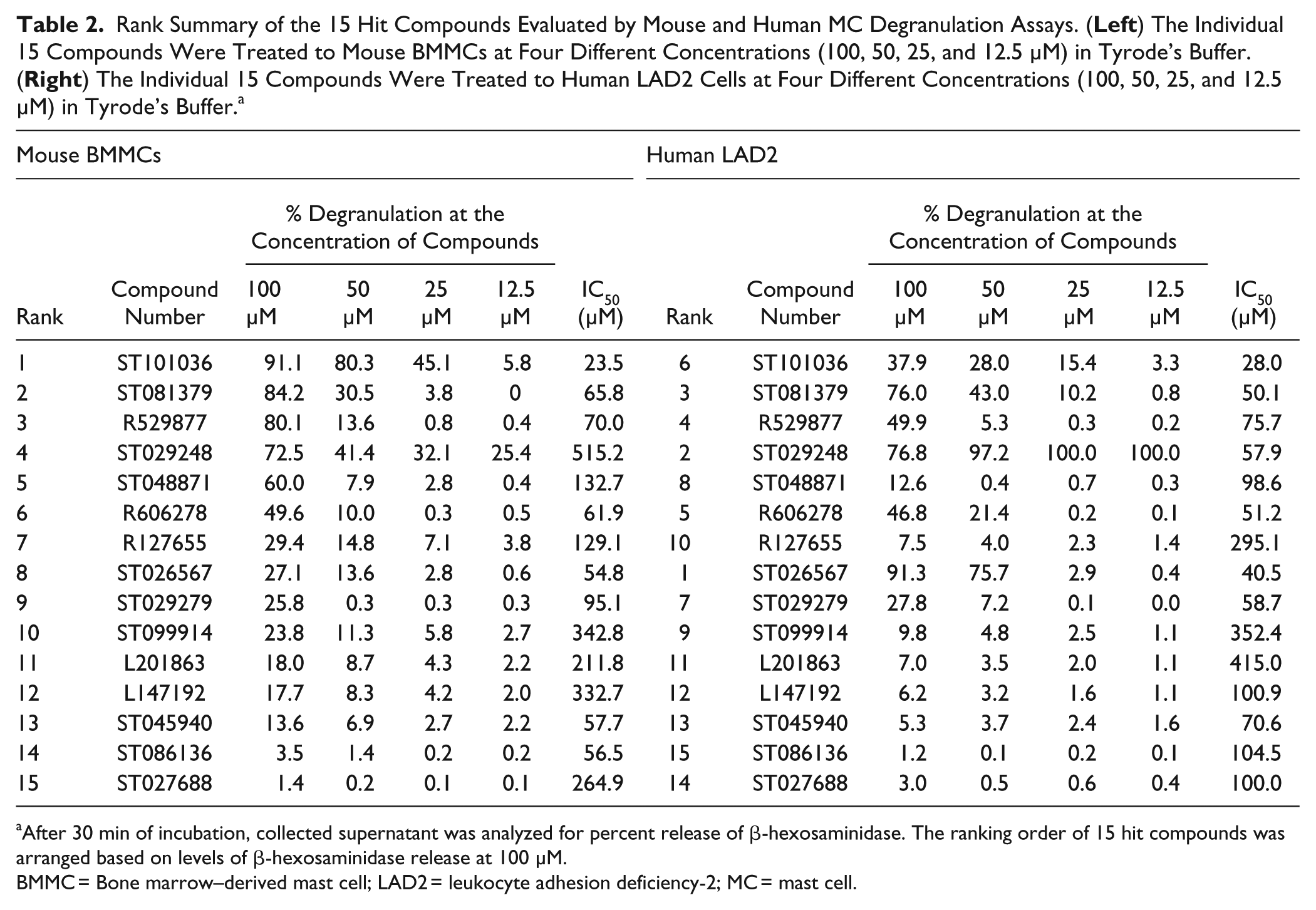
After 30 min of incubation, collected supernatant was analyzed for percent release of β-hexosaminidase. The ranking order of 15 hit compounds was arranged based on levels of β-hexosaminidase release at 100 µM.
BMMC = Bone marrow–derived mast cell; LAD2 = leukocyte adhesion deficiency-2; MC = mast cell.
Next, human LAD2 MCs were treated with the 15 hit compounds to measure the degree of human MC response ( Table 2 ). The compounds ST026567, ST029248, and ST081379 exhibited more than 70% degranulation at 100 µM. We noticed that degranulation of human and mouse MCs was strongly induced by both ST029248 and ST081379. Specifically, when human LAD2 cells were treated with ST029248 at four different doses, the levels of degranulation were maintained at more than 90%, while treatment with other compounds at different doses showed dose-dependent MC degranulation levels. ST101036, which was the most potent inducer of mouse BMMC degranulation (91%), exhibited 38% degranulation of human LAD2 cells, whereas ST026567, which was a mild inducer of mouse BMMC degranulation (27.1%), exhibited 91% degranulation of human LAD2 cells. Although ST027688 and ST086136 were shown to be poor activators of mouse BMMC degranulation (~1% degranulation) and of the human LAD2 line, these two compounds provided more than 10% degranulation activity in primary human MCs (data not shown). We suspect that different sources of MCs provide various sensitivities to specific compounds. We kept these compounds in our hit list, however, to retain broad chemical structural diversity in our selected compounds. In summary, these results indicate that ST101036 and ST026567 are potent MC activators that exhibit species specificity, but ST029248 is a potent MC activator regardless of tested species (mouse or human).
Dose–Response Degranulation of Human LAD2 Cells on Treatment with the 15 Hit Compounds
As described above, certain compounds such as ST029248 exhibited near-maximal degranulation activities over the tested compound concentrations (100, 50, 25, and 12.5 µM). Therefore, we evaluated degranulation response using a broader range of compound concentrations (200 to 0.78 µM) and calculated the concentration of each compound that induced 50% MC degranulation (MCD50) and 10% MC degranulation (MCD10) ( Fig. 3 ). Figure 3C shows sigmoidal dose–response curves based on the degranulation of the human LAD2 MC line in response to three representative compounds (ST026567, ST029248, and R529877) compared to M7. MCD50 and MCD10 values of all 15 hit compounds were interpolated from the calculated dose–response sigmoidal curves of individual compounds. The complete list of MCD50 and MCD10 values for all 15 hit compounds in LAD2 cells is shown in Figure 3D . When we compared the values of all tested compounds, ST029248 showed the lowest MCD50 value (2.9 µM) and MCD10 value (1.5 µM), which demonstrated approximately threefold higher efficacy than the positive control, M7 (MCD50 = 7.0 µM, MCD10 = 4.4 µM). The MCD50 and MCD10 values for the remaining compounds were higher than those for M7. When we compared the ranked list of Figure 3D (MCD10 values) to that of Table 2 (ranking order from the degranulation response at 100 µM), the ranking of MC-activating potency was significantly correlated ( Fig. 3E ). Altogether, the calculated MCD10 parameter enabled the degranulation efficacy of the compounds to be compared, especially when the degranulation response reached saturating levels.
Cytokine Production by MCs on Stimulation with MC-Degranulating Compounds
MCs can produce a plethora of biological mediators, including proinflammatory cytokines, through either degranulation or de novo synthesis. 26 The cytokines released from activated MCs are known to regulate a rapid response as innate immunity and generate a long-term response as adaptive immunity. 1 Traditional enzyme-linked immunosorbent assays (ELISAs) have limitations such as measuring only one cytokine at a time and the difficulty to investigate a full profile of cytokines, technically requiring a large sample volume to measure multiple cytokines. Therefore, we performed Luminex-based multiplex profiling to measure multiple secreted cytokines, chemokines, and growth factors released to the culture media during MC activation. In the assays, after mouse BMMCs or human LAD2 cells were treated with individual positive controls (M7 and ionomycin/PMA) or the 15 hit compounds at 12.5 µM for 20 h, we measured 20 human or 15 mouse cytokines. We chose 12.5 µM to induce the secretion of de novo synthesized cytokines, because compound treatment was for a longer period (20 h) for the de novo synthesis assay as compared to a short period (30 min) of treatment for inducing degranulation at 100 µM. Figure 4A shows the patterns of cytokine secretion from mouse BMMCs, and Figure 4B shows the same for human LAD2 cells. Ionomycin + PMA treatment of mouse BMMCs induced the widest range of cytokines. Although M7 induced mouse BMMCs to secrete only two cytokines, compounds ST029248, ST101036, ST081379, ST026567, and ST027688 induced mouse BMMCs to secrete more than three cytokines. ST029248 caused mouse BMMCs to secrete the most diverse cytokines, including chemokines and growth factors. MCP1, MIP2, and IL6 are the most frequently detected cytokines after treatment with hit compounds. Unexpectedly, R127655, ST045940, and R606278 failed to induce detectable cytokine secretion at 12.5 µM, although these compounds generated close to 50% degranulation at a 100 µM concentration, indicating that MC degranulation and induction of de novo synthesis are not necessarily strongly coupled.
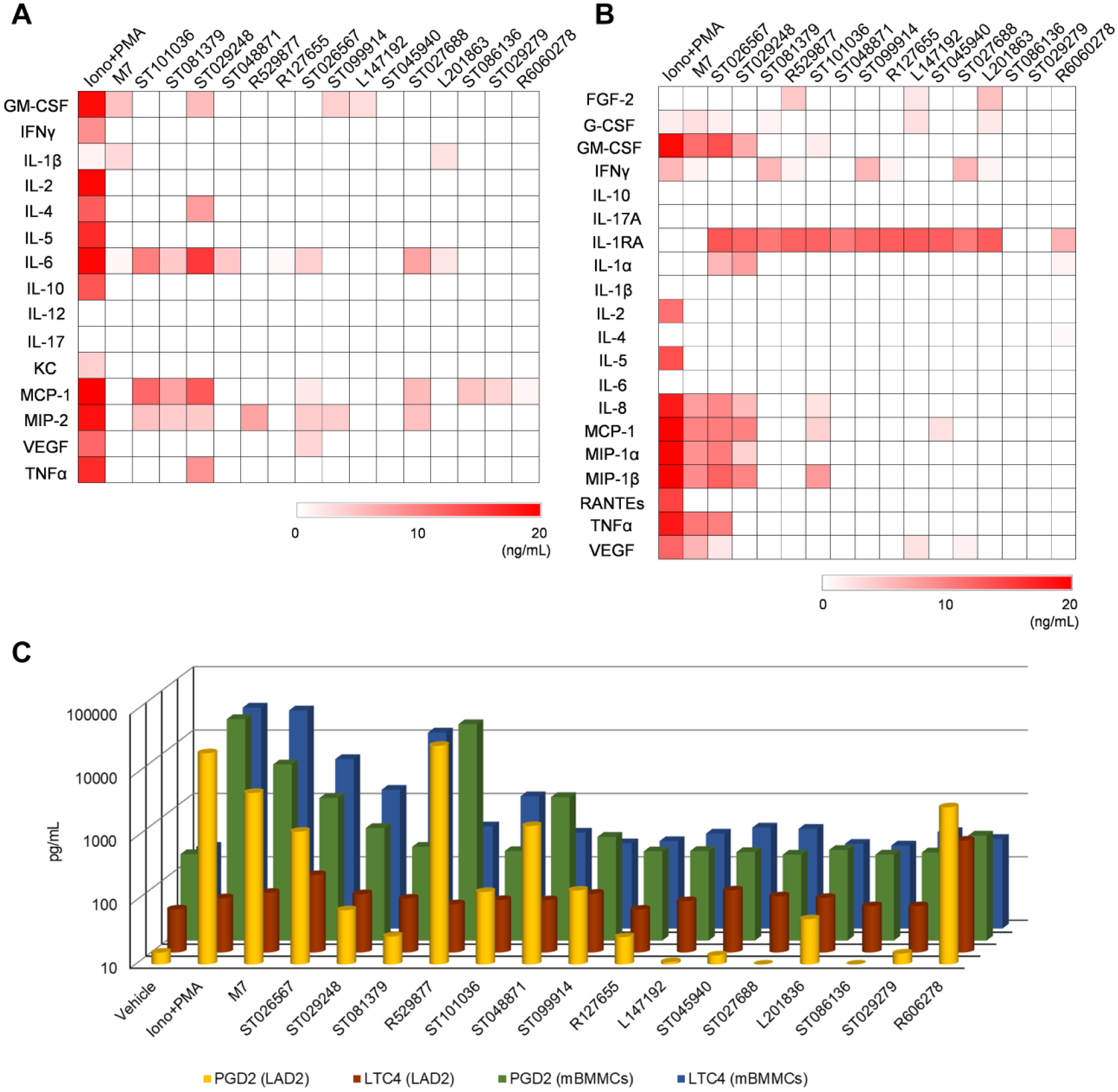
Secretion profiles of mast cell (MC) mediators on hit compounds treatment. (
When we analyzed the secretion of cytokines from human LAD2 cells at a concentration of 12.5 µM for 20 h of treatment, treatment with the positive controls, ionomycin + PMA or M7, induced a wide range of cytokine secretion, as expected ( Fig. 4B ). ST026567, which was a mild cytokine inducer in mouse BMMCs, had the broadest profile of cytokine secretion (as many as nine cytokines) among the 15 hit compounds. ST029248, which was the most potent cytokine secretor in mouse BMMCs, still exhibited potent activity in human LAD2 cells with secretion of seven cytokines. ST101036, L147192, and L201863 exhibited secretion of four to five cytokines from human LAD2 cells. The remaining hit compounds, however, induced minor or undetectable cytokine secretion. Summarizing, ST029248, ST026567, and ST101036 induced secretion of multiple cytokines from both human and mouse MCs, and ST029248 showed the strongest activity among the three compounds.
MC Secretion of Arachidonic Acid Metabolites on Stimulation with MC-Degranulating Compounds
In addition to cytokine and chemokine production, activated MCs release various lipid mediators, causing vasodilation and inflammation. These lipid mediators have been shown to be involved in host protection against infection and the regulation of adaptive immunity. Arachidonic acid, as a precursor, is processed by phospholipases into prostaglandins and leukotrienes, which bind directly to the receptors on various cells nearby. 27 In this study, we measured the release of arachidonic acid metabolites from human and mouse MCs, including PGD2 and LTC4. Human LAD2 cells or mouse BMMCs were treated with the 15 hit compounds (50 µM) or positive controls (ionomycin + PMA and M7) for 2 h, and the culture supernatants were tested for the secretion of PGD2 or LTC4. Because treatment was for only 2 h, we increased the concentration to 50 µM, as compared to the lower concentration (12.5 µM) used for a longer period of incubation (20 h) when measuring de novo synthesized cytokines. Figure 4C illustrates that treatment with M7 and ionomycin + PMA induced human LAD2 cells to secrete high levels of PGD2 (yellow bars), as expected. Compound R529877 induced the highest levels of PGD2 secretion among the compounds tested. ST026567, ST048871, and ST099914 also induced the secretion of PGD2 from LAD2 cells, with levels as high as 1 ng/mL. When we quantified the secretion of PGD2 from mouse BMMCs, R529877 induced the highest levels of PGD2 secretion among the compounds tested, which is consistent with what was observed in LAD2 cells ( Fig. 4C , green bars). ST026567, ST048871, and ST099914 induced similar levels of secretion of PGD2 from mBMMCs, with levels as high as 5 ng/mL. The remaining compounds, which exhibited minimal dose-dependent PGD2 secretion, failed to secrete PGD2 from mBMMCs.
Production of LTC4 was also monitored. Figure 4C (brown bars) showed that treatment of LAD2 cells with positive controls (ionomycin + PMA and M7) induced minimal levels or no secretion of LTC4 (~100 pg/mL). ST026567 induced higher levels of LTC4 secretion (~200 pg/mL) from LAD2 cells. Next, we quantified the secretion of the LTC4 from mouse BMMCs, as we did for human LAD2 cells ( Fig. 4C , blue bars). The positive controls induced mBMMCs to secrete LTC4 as high as ~10 ng/mL. Compound ST026567, which was the strongest inducer of LTC4 secretion in LAD2 cells, induced a similar level of LTC4 secretion (~10 ng/mL) from mBMMCs compared to the positive control treatments. Although different cell types generated different levels of lipid mediators, R529877 and ST026567 induced the most potent PGD2 and LTC4 secretion among all cell types.
Because MCs have been implicated in the regulation of immune responses through activating both innate and adaptive immunity, MC-activating compounds may be useful for a variety of therapeutic applications in which activation of the immune system is beneficial. Supporting this notion is the observation that MC activators provide potent vaccine adjuvant activity that boosts pathogen-specific humoral immune responses. In addition, MC activation was shown to be efficacious in generating cell-mediated immunity and used as an adjuvant for cancer immunotherapy. 13 Our study identified 15 MC activators by using HTS assays. These novel MC activators were confirmed to degranulate mouse and human MCs, and even to induce secretion of various cytokines and lipid mediators that are associated with proinflammatory responses. These 15 novel MC activators may be useful in therapeutic applications that may benefit from MC activation, including use as vaccine adjuvants.
Supplemental Material
DS_DISC834699 – Supplemental material for Identification of Novel Mast Cell Activators Using Cell-Based High-Throughput Screening
Supplemental material, DS_DISC834699 for Identification of Novel Mast Cell Activators Using Cell-Based High-Throughput Screening by Hae Woong Choi, Cliburn Chan, Ivo D. Shterev, Heather E. Lynch, Taylor J. Robinette, Brandi T. Johnson-Weaver, Jianling Shi, Gregory D. Sempowski, So Young Kim, John K. Dickson, David M. Gooden, Soman N. Abraham and Herman F. Staats in SLAS Discovery
Footnotes
Acknowledgements
We acknowledge the valuable assistance of Duke Regional Biocontainment Laboratory staff member K. Riebe. We would also like to acknowledge the Duke Functional Genomics Shared Resource for technical support with assays.
Supplemental material is available online with this article.
Declaration of Conflicting Interests
The authors declared the following potential conflicts of interest with respect to the research, authorship, and/or publication of this article: S.N.A. is the cofounder and Chief Scientific Officer for Mastezellen Bio Inc. H.F.S. is the cofounder and Chief Executive Officer of Mastezellen Bio Inc. None of the work described in this manuscript was performed by Mastezellen.
Funding
The authors disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: Multiplex analysis was performed in the Immunology Unit of the Duke Regional Biocontainment Laboratory, which received partial support for construction from the National Institutes of Health, National Institute of Allergy and Infectious Diseases (UC6-AI058607). S.N.A. is the cofounder and chief scientific officer for Mastezellen Bio Inc. H.F.S is the cofounder and chief executive officer of Mastezellen Bio Inc. This work was funded by NIH contract HHSN272201400054C.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.