
Abstract
Polyamines play an important role in cell growth, differentiation, and cancer development, and the biosynthetic pathway of polyamines is established as a drug target for the treatment of parasitic diseases, neoplasia, and cancer chemoprevention. The key enzyme in polyamine biosynthesis is ornithine decarboxylase (ODC). We report herein an analytical method for the continuous fluorescence monitoring of ODC activity based on the supramolecular receptor cucurbit[6]uril (CB6) and the fluorescent dye trans-4-[4-(dimethylamino)styryl]-1-methylpyridinium iodide (DSMI). CB6 has a significantly higher binding constant to the ODC product putrescine (>107 M−1) than to the substrate L-ornithine (340 M−1). This enables real-time monitoring of the enzymatic reaction through a continuous fluorescence change caused by dye displacement from the macrocycle by the formed product, which allowed a straightforward determination of enzyme kinetic parameters (kcat = 0.12 s−1 and KM = 24 µM) and inhibition constants of the two ODC inhibitors α-difluoromethylornithine (DFMO) and epigallocatechin gallate (EGCG). The potential for high-throughput screening (HTS) was demonstrated by excellent Z′ factors (>0.9) in a microplate reader format, and the sensitivity of the assay is comparable to or better than most established complementary methods, which invariably have the disadvantage of not being compatible with direct implementation and upscaling to HTS format in the drug discovery process.
Introduction
Enzymes play a crucial role in physiological processes, and their upregulated or downregulated activity has been associated with major diseases, including numerous types of cancer. 1 Moreover, many pathogens require specific enzymes for cellular entry or infection. Enzymes are thus considered important therapeutic targets for drug discovery by the pharmaceutical industry. Identification of novel, specific, and potent modulators of enzymatic function relies to a high degree on developing scalable in vitro assay methods and performing high-throughput screening (HTS) of collections of small molecules.2,3 The HTS paradigm in turn requires the careful design of enzyme assays, which are compatible with existing plate-based screening infrastructure and use of sensitive optically based readouts, such as fluorescence intensity measurements, as a measure of target function. 4
Polyamines are a class of compounds that are associated with the genesis and metastasization of various types of cancer, including breast, skin, colorectal, bladder, and cervical cancer.
5
For example, increased cell proliferation, decreased apoptosis, and increased expression levels of genes, which influence tumor invasion and metastasis, are accompanied by elevated polyamine levels. The rate-limiting enzyme in the biosynthesis of polyamines in mammals is ornithine decarboxylase (ODC), which catalyzes the decarboxylation of
Because of these promising avenues, new ODC inhibitors are currently being sought in academia and industry with the goal to either identify more potent inhibitors of human ODC with improved pharmacokinetic properties or find ODC inhibitors, which are selective toward pathogen-specific ODC variants.11,12 However, ODC inhibitors are difficult to identify, because of the lack of suitable ODC enzyme assays. The established methods for monitoring ODC activity include assays based on detection of radiolabeled putrescine, 13 chemiluminescence-based 14 or colorimetric detection of H2O2 produced by oxidation of putrescine by amine oxidases,8,15 electrochemical measurements, 16 and high-performance liquid chromatography (HPLC)–based methods.17–19 All these methods require several incubation or washing steps, for example, spotting of cellular extracts onto phosphocellulose, 14 differential extraction of picrylsulfonic acid adducts from water into 1-pentanol, 8 and postcolumn or precolumn derivatization for HPLC. 18 This renders these methods unfavorable for HTS such that no HTS-compatible enzyme assays for ODC are currently available, which would allow for systematic identification and exploration of large libraries of potential inhibitors.
We now report a robust and convenient fluorescence-based assay for continuously monitoring ODC activity based on our previously reported supramolecular tandem enzyme assay principle ( Fig. 1 ).20–22 Tandem assays rely on reporter pairs composed of a macrocyclic receptor and a fluorescent dye, which bind to each other, affecting a change in the fluorescence spectroscopic properties of the dye. In tandem assays, the receptor additionally interacts with either the substrate or the product of the enzymatic reaction, leading to a displacement of the fluorescent dye. Upon enzymatic conversion of the substrate into the product, the overall propensity of the reaction mixture to displace the fluorescent dye from the macrocyclic host changes; that is, the enzymatic reaction either converts a weakly binding substrate into a strongly binding product or vice versa, which generates a detectable fluorescence change in any case. This affords an alternative, simple, inexpensive, sensitive, and label-free method to continuously monitor enzymatic reactions. 21
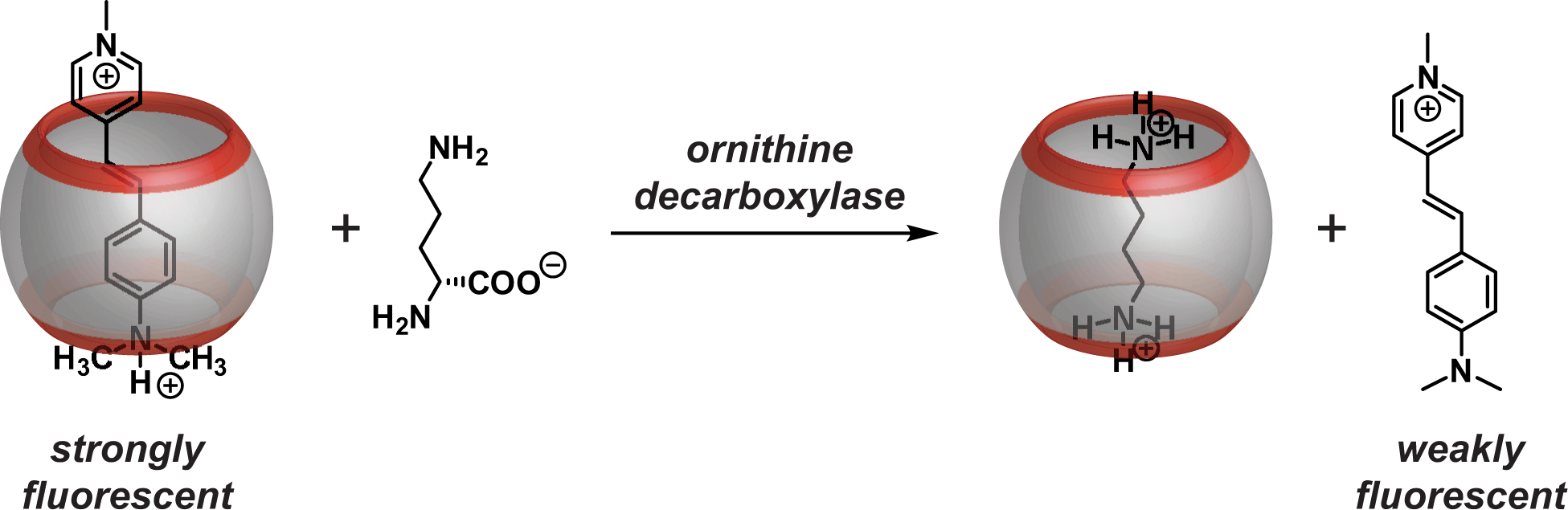
Working principle of the fluorescence-based supramolecular tandem enzyme assay for ODC.
To adapt the tandem assay principle to ODC ( Fig. 1 ), we selected trans-4-[4-(dimethylamino)styryl]-1-methylpyridinium iodide (DSMI) as the fluorescent dye and cucurbit[6]uril (CB6) as the macrocyclic receptor, which were recently reported to form a highly fluorescent inclusion complex.23,24 The substrate of ODC binds only very weakly to CB6, whereas the product putrescine is a strong competitor. As a consequence, ODC activity generates a strongly binding product, which continuously displaces the fluorescent dye with time and results in a concomitant decrease in fluorescence intensity. We demonstrate that the resulting product-selective tandem assay 21 can be used to determine enzyme kinetic parameters of ODC and for inhibitor screening with DFMO and (−)-epigallocatechin gallate (EGCG) as two exemplary known inhibitors.5,8,25,26 The performance of the assay was also evaluated in a microplate reader format, which gave excellent Z′ factors, revealing its potential for HTS. 27
Materials and Methods
Materials
All chemicals, including the fluorescent dye DSMI, were from Sigma-Aldrich (Steinheim, Germany) and used as received, except for CB6, which was received from Strem Chemicals (Kehl, Germany). Human ODC (0.702 µg/µL) expressed in HEK cells was received from Origene (Rockville, MD) and stored at −60 °C. The Tris buffer was prepared from Tris base, and the pH was adjusted by addition of HCl. Stock solutions of pyridoxal 5′-phosphate (PLP) and dithiothreitol (DTT) were prepared fresh on a daily basis and stored at 4 °C.
Spectroscopic Measurements
Absorbance measurements were performed with a Varian Cary 4000 spectrophotometer. Fluorescence was measured with a Varian Cary Eclipse spectrofluorometer equipped with a temperature controller. All spectroscopic measurements were performed in 0.5 or 3.5 mL quartz glass cuvettes (Hellma Analytics, Müllheim, Germany).
Binding Titrations
Fluorescence titrations were performed at ambient temperature by successive addition of known concentrations of the titrant. It was ensured that the concentrations of all other components were kept constant. Commonly, the change in fluorescence intensity in the spectral area of largest variation was plotted against the concentration of titrant and analyzed by nonlinear regression by using a 1:1 host–guest binding equation (for CB6 with DSMI) or a competitive binding equation (all others) implemented in OriginPro 8.5 (OriginLab Corporation, Northampton, MA).
Continuous Enzyme Assays
Continuous fluorescence assays were performed in 0.5 mL quartz glass cuvettes with 3 µM DSMI in 10 mM Tris-HCl, pH 7.5, at 37.0 ± 0.1 °C. The sample was allowed to equilibrate for at least 15 min, and the fluorescence intensity was subsequently continuously monitored (λexc = 450 nm, λem = 582 nm) as a function of time during the addition of 0.5 µM CB6 and varying amounts of ODC (typically 0.6 ng/µL) added from stock solutions containing EDTA, DTT, PLP, and Tween 80 to afford final concentrations of 0.1 mM EDTA, 2.5 mM DTT, 0.1 mM PLP, and 0.1% Tween 80. Reactions were initiated by addition of
Microplate Measurements
Microplate measurements were performed with a JASCO FP-8500 spectrofluorometer coupled with a JASCO FMP-825 microplate reader accessory in 96-well microplates and with an EnSpire multimode plate reader (PerkinElmer, Waltham, MA) in 384-well microplates. Both microplates were flat-bottom microplates with a nonbinding surface from Corning (New York, NY, product nos. 3820 and 3650).
For the enzymatic reactions in 96-well microplates, 234 µL of a solution containing 3 µM DSMI and 0.5 µM CB6 in 10 mM Tris-HCl, pH 7.5, were added to each well and equilibrated for >8 min. Then, 24 µL of 0.5 ng/µL ODC (added from a stock solution containing EDTA, DTT, PLP, and Tween 80 to afford final concentrations of 0.1 mM EDTA, 2.5 mM DTT, 0.1 mM PLP, and 0.1% Tween 80) was added and the reaction was initiated by addition of 2 µL of
For the enzymatic reactions in 384-well microplates, 6 µL of a solution containing 3 µM DSMI and 0.03–0.5 µM CB6 in 10 mM Tris-HCl, pH 7.5, was added to each well and equilibrated for >15 min. Then, 3 µL of 1 ng/µL ODC (added from a stock solution containing EDTA, DTT, PLP, and Tween 80 to afford final concentrations of 0.1 mM EDTA, 2.5 mM DTT, 0.1 mM PLP, and 0.1% Tween 80) was added and the reaction was initiated by addition of 1 µL of a 200 µM
After initiating the reactions, the microplate was immediately placed into the reader and the fluorescence intensity (λexc = 450 nm, λem = 582 nm) of each well was recorded at specific time intervals.
Results and Discussion
Characterization of the CB6/DSMI Reporter Pair
To date, various supramolecular tandem assays have been introduced, including supramolecular tandem assays for amino acid decarboxylases. 20 In these cases, two different supramolecular reporter pairs were used, calix[4]arene (CX4) with 2,3-diazabicyclo[2.2.2]oct-2-ene as a fluorescent dye and cucurbit[7]uril (CB7) with dapoxyl. Literature precedence, however, suggested that the smaller-homologue CB6 has a significantly higher binding constant to certain biogenic amines, in particular to the ODC product putrescine, 28 such that CB6 was considered superior to CB7 in an ODC assay and expected to afford an increased sensitivity. So far, the utility of CB6 was limited by the lack of suitable fluorescent dyes. While the cavity of CB7 is sufficiently large to accommodate various fluorescent dyes, for example, dapoxyl, acridine orange, and berberine chloride, the choice of dyes binding to CB6 had been limited to UV-absorbing dyes. 29 The recently reported DSMI lifted this limitation and shows an absorbance as well as fluorescence emission, at relatively long wavelengths when bound to CB6 (λabs = 450 nm, λem = 582 nm), accompanied by a strong increase in fluorescence intensity by a factor of ca. 270. 23
To ensure that these beneficial properties are retained under the conditions relevant for enzyme assays, titrations of DSMI with CB6 in 10 mM Tris-HCl at pH 7.5 were performed. In accordance with titrations performed in neat water, 23 addition of CB6 to DSMI produced a negligible decrease in absorbance, whereas the fluorescence increased strongly ( Fig. 2 ). A plot of the fluorescence intensity at maximum emission wavelength (582 nm) and analysis by a 1:1 host–guest titration curve gave a binding constant of Ka = 7.0 × 105 M−1, which is in good accordance with the literature value. 23 The combined results revealed DSMI as the dye of choice for supramolecular tandem enzyme assays with CB6. The reported standard buffer for ODC additionally contains the reducing agent DTT, EDTA, the cofactor PLP, and Tween 80.6,7 Therein, the apparent binding constant of DSMI to CB6 was slightly reduced (Ka = 1.3 × 105 M−1; see Suppl. Fig. S1 ), which is most likely due to competitive binding of the buffer components. Also noteworthy, the standard concentration of PLP in ODC buffer led to a relatively strong absorbance at the excitation wavelength of DSMI (ca. 0.4 at 0.1 mM PLP), which may have led to a nonlinear fluorescence response due to the inner filter effect, which was not explicitly corrected for.
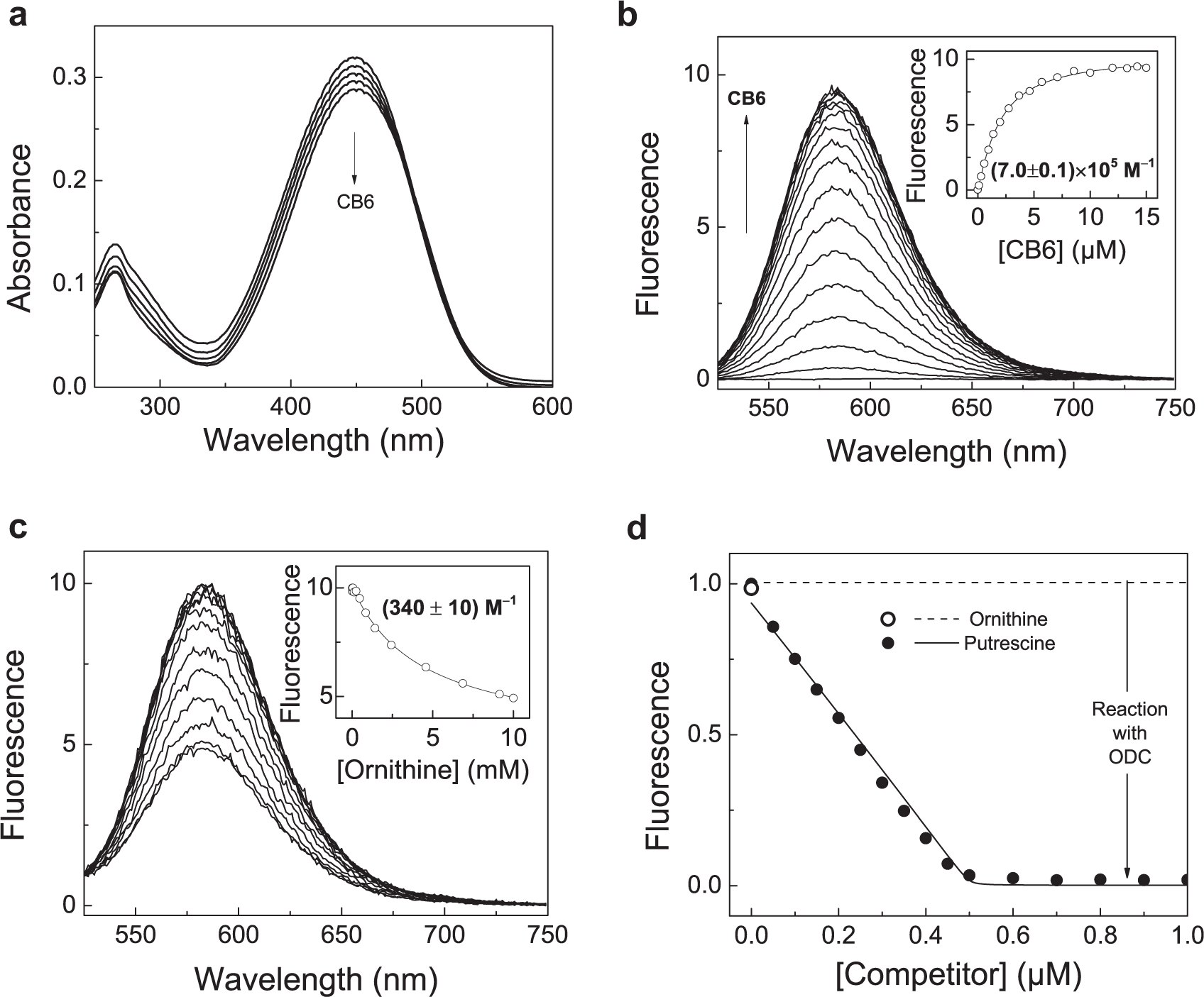
Determination of the binding constant of DSMI to CB6 and response of the reporter pair to the substrate
The response of the CB6/DSMI reporter pair toward the substrate and product of the enzymatic reaction was explored by competitive titrations, in which the substrate or product is successively added in small portions to a solution containing the reporter pair. This revealed that large quantities of ornithine (>1 mM) were required to efficiently displace DSMI, and fitting of the resulting titration curve by nonlinear regression gave a very small binding constant of (340 ± 10) M−1 ( Fig. 2c ). In contrast, the enzymatic reaction product putrescine efficiently displaced DSMI at nanomolar concentrations, yielding an almost linear dependence (R2 > 0.99 in the range of 0–0.5 µM) of the fluorescence intensity on putrescine concentration ( Fig. 2d ). This indicates that binding of putrescine to CB6 is quantitative under these conditions, pointing to a very high binding constant, Ka > 107 M−1. Interesting to note, the linear response also demonstrates that the high absorbance of PLP at the DSMI excitation wavelength (see above) is not further detrimental for a quantitative analysis of the fluorescence data. Presumably, the influence of the inner filter effect on the fluorescence intensity remains constant, because the absorbance of the additive, albeit significant, remains constant in the course of the titration.
Generally, supramolecular tandem assays can even be performed with moderate binding constants and comparably low selectivity for either substrate or product, but in these cases, a more elaborate data analysis is required, which involves several interrelated binding equilibria to extract reaction rates from the resulting nonlinear fluorescence response.20,21 The stark difference between the binding constants of the substrate ornithine and the product putrescine (>4 orders of magnitude) offers a simplified analysis with the added possibility to investigate a large working range of substrate concentrations. In detail, a simple linear dependence of the fluorescence intensity on the amount of formed product applies at all concentrations.
ODC Assay and Enzyme Kinetics
The ability to follow the reaction of ODC with the DSMI/CB6 reporter was demonstrated next ( Fig. 3 ). Here, the fluorescence intensity of a solution containing DSMI in 10 mM Tris-HCl, pH 7.5, was continuously monitored with time upon addition of CB6, ODC, and ornithine. As expected, addition of CB6 led to a strong increase in fluorescence due to complexation of DSMI by CB6. Subsequent addition of ODC in a buffer containing sufficient amounts of DTT, EDTA, PLP, and Tween 80 to give the desired final concentrations after dilution gave a ca. 2.7-fold decrease in fluorescence, due to a slightly reduced binding constant and quenching. Lastly, addition of the substrate ornithine resulted in a continuous, time-dependent decrease in fluorescence, which is indicative of the progress of the enzymatic reaction and which was absent when no ODC was present ( Fig. 3b ).
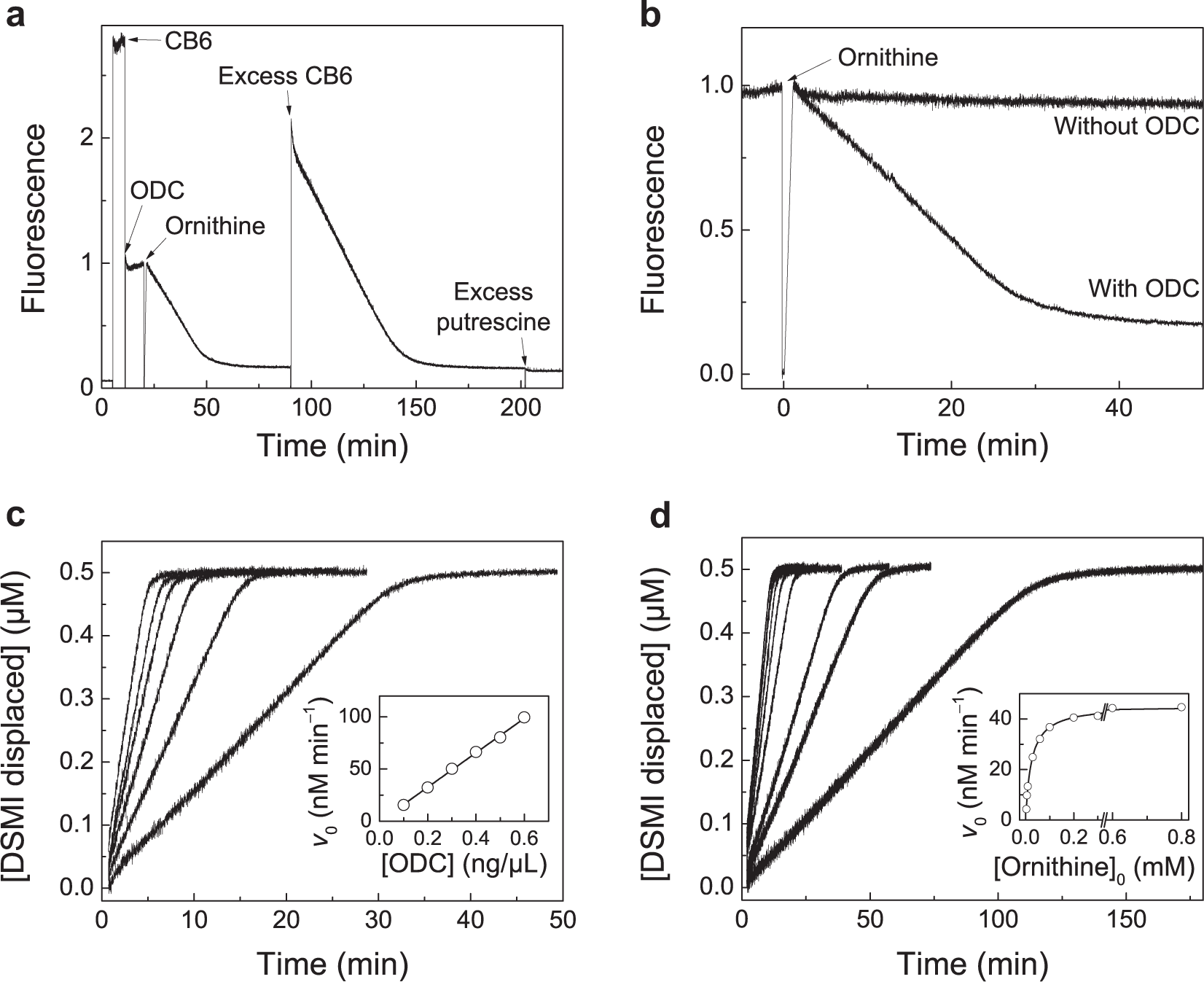
Continuous fluorescence enzyme assays with ODC. (
The time-dependent change in fluorescence progressed until a plateau of constant fluorescence was reached (
Fig. 3a
). Of note is that the fluorescence changed almost linearly, a response that differs from that of many conventional enzyme assays. Usually, the final plateau region indicates that all substrate has been converted into its product, and the overall reaction rate decreases as the enzymatic reaction proceeds due to substrate depletion. In the supramolecular tandem assays presented herein, however, the fluorescence reaches a plateau, because all CB6 is occupied by the product putrescine. In fact, the overall change in fluorescence in
Figure 3a
refers to only 0.25% substrate conversion (i.e., 200 µM
The peculiar response, which is desirable because it signals minute conversions and reflects the critical initial rates, could be further verified by addition of an excess of CB6 after reaching the plateau value, which regenerated the fluorescence and showed again a time-dependent decrease until a plateau region of the same final intensity was reached. This control experiment also demonstrated that the decrease in fluorescence is due to the conversion of ornithine to putrescine and not an artifact, for example, from thermal or photodecomposition. It also confirmed that the enzyme remained active, which was independently assessed for up to 36 h (see Suppl. Fig. S3 ). As an additional control experiment, an excess of putrescine was added at the end (after ca. 200 min), which had no influence on the final fluorescence intensity and confirmed that no undesirable interactions occur in this complex mixture.
The large selectivity between substrate and product results in a linear response of the reporter pair on product concentration, whereas the substrate does not interfere in a wide concentration range. This allows an easy conversion of the fluorescence data into concentration changes by considering that at the time point of enzyme addition, no product has yet formed ([product] = 0 µM), while the plateau region indicates that all DSMI has been displaced from the available CB6 ([product] = [CB6]). The fluorescence traces were normalized with these considerations; that is, the fluorescence at the time point before initiating the reaction was subtracted from the time-dependent fluorescence data (= 0 µM), and subsequently, the fluorescence traces were multiplied with a factor to set the final plateau value to 0.5 µM (see Fig. 3a , b for original fluorescence traces and Fig. 3c , d for normalized traces). Linear fitting of the fluorescence progress curve thus allows a quantitative determination of the initial enzyme reaction rates, v0. Noteworthy, the exchange kinetics of supramolecular receptors are commonly much faster than milliseconds, and the response toward analytes, either added during a titration or generated during an enzymatic reaction, is immediate.21,24
The dependence of the v0 values on varying enzyme and substrate concentrations could thus be conveniently investigated, and showed the expected linear dependence of the reaction rate on the ODC concentration (inset of Fig. 3c ) and the expected Michaelis–Menten behavior, including saturation at high substrate concentrations (inset of Fig. 3d ). Analysis of the Cornish–Bowden diagram with the Michaelis–Menten equation afforded the catalytic turnover number (kcat = [0.12 ± 0.01] s−1) and the Michaelis–Menten constant (KM = 24 ± 1 µM) for human ODC expressed in HEK cells. Considering differences in reaction conditions, assay types, and enzyme sources, the obtained values are in satisfactory agreement with previously reported literature values (e.g., kcat = 3.3 s−1 and KM = 80 µM for a human ODC expressed with the pUC vector in Escherichia coli). 7
Determination of Inhibition Constants
ODC is a potential target enzyme for treating certain types of cancer, as well as parasitic infections,6–8 and consequently, a simple fluorescence-based enzyme assay would be desirable to identify additional ODC inhibitors in the future.11,12 To demonstrate the utility of our supramolecular tandem assay in identifying ODC inhibitors, we selected DFMO and EGCG as two established inhibitors (see Fig. 4 ).25,26 DFMO is the most widely studied prototypical suicide inhibitor of ODC. It forms after decarboxylation a covalent bond with a cysteine residue in the active site of ODC, and thereby irreversibly suppresses ODC activity.19,26 DFMO is currently in clinical trials as a promising chemopreventive agent in the therapy of colorectal cancer, nonmelanoma cutaneous cancer, bladder cancer, cervical cancer, and neuroblastoma.5,9,10 EGCG is the most abundant polyphenol in green tea, and may be associated with the claimed beneficial effects of green tea. It has been shown that EGCG inhibits various kinds of protein kinases, thus interfering with their signal transduction pathway, and EGCG is a known inhibitor of ODC. 25
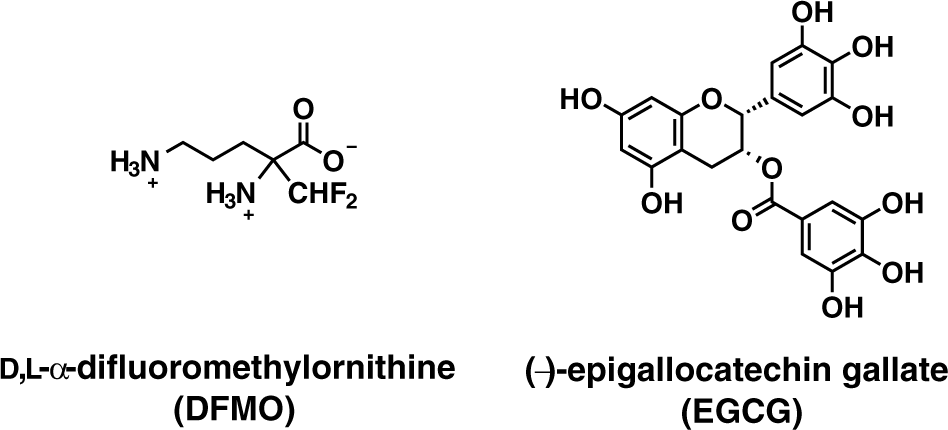
Structures of inhibitors.
To study the inhibition of ODC by DFMO and EGCG with our supramolecular tandem assay involving the CB6/DSMI reporter pair, the time-dependent fluorescence changes in the presence of varying inhibitor concentrations were recorded and normalized in the same manner as described above for the determination of its enzyme kinetics ( Fig. 5a , c ). Importantly, both inhibitors did not significantly bind to CB6 (Ka < 500 M−1; see Suppl. Fig. S2 ), which would otherwise need to be accounted for (but which is generally only likely for polycationic molecules or residues with sufficiently low molecular weight to become encapsulated into the cavity of CB6). The initial enzyme reaction rate obtained by linear fitting of the fluorescence progress curve was then plotted against the inhibitor concentration and analyzed with the common Hill equation ( Fig. 5b , d ). This yielded the IC50 of both inhibitors, that is, the inhibitor concentrations needed to reduce the enzyme activity by 50%, which approximates to the inhibition constant Ki, when the actual concentration of the enzyme falls far below the IC50 value, which is fulfilled.
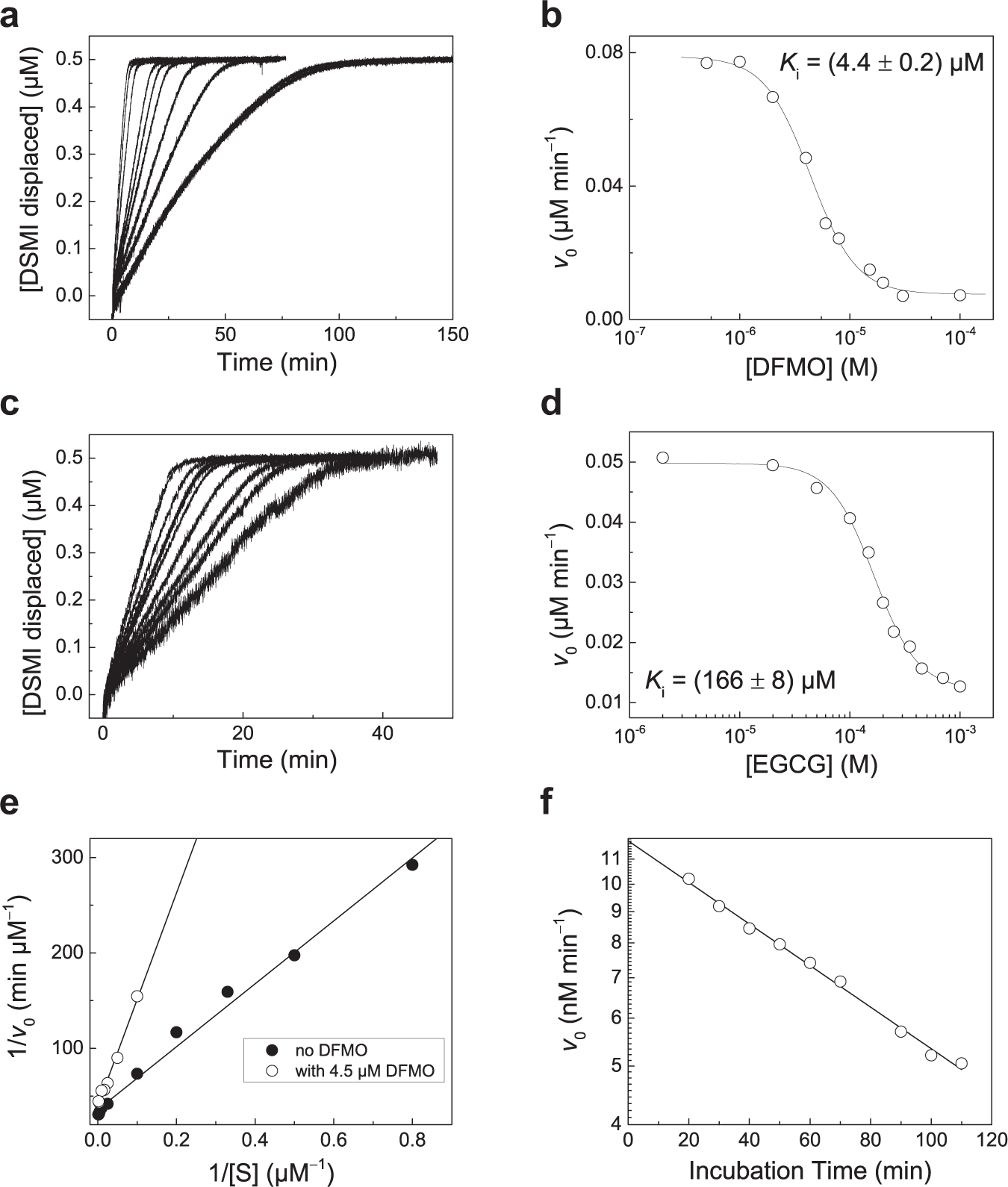
Determination of inhibition constants of DFMO (
The inhibition constants obtained with our supramolecular tandem assay were Ki = 4.4 ± 0.2 µM for DFMO and Ki = 166 ± 8 µM for EGCG. The value for DMFO is in good agreement with a literature value of 2.2 µM for the racemic mixture, 19 and Ki > 100 µM was previously deduced for EGCG. 30 This demonstrates that our supramolecular tandem assay for ODC can be used to reliably determine inhibition constants, even for weakly inhibiting compounds such as EGCG.
To demonstrate the possibility to also determine the mode of inhibition, DFMO was investigated in more detail ( Fig. 5e , f ). At short incubation times (<20 min), the Lineweaver–Burk plots in absence and presence of DFMO revealed a competitive inhibition mode ( Fig. 5e ), which is in accordance with reversible binding to the active site before the enzyme is irreversibly inactivated. With increasing incubation times of DFMO with ODC, the activity of ODC continued to decrease as expected for suicide inhibitors. These results are in accordance with the literature on this well-investigated inhibitor. 19
Microplate Measurements
To demonstrate the potential of our ODC assay in HTS applications, we have also miniaturized the detection system and performed measurements in microtiter plates, which would allow the screening of several inhibitors or activators in parallel. This allows us to judge the statistical variability of the assay by the Z′ factor. 27 Here, the fluorescence signal in the absence of enzyme was taken as control and the signal of the plateau region after addition of enzyme was taken as the sample. This gave a Z′ factor for this assay of 0.96 in 96-well microplates and of 0.90 in 384-well microplates, which is considered excellent, as a Z′ value of 1.0 represents an ideal HTS assay, and even a value of 0.5 is normally taken as an acceptable HTS assay. We could also determine the inhibition constant of DFMO in a 384-well microplate format, which gave a value of 4.9 ± 0.2 µM, in excellent agreement with our cuvette-based measurements (see above).
Importantly, only 130 pmol putrescine is needed to afford a complete dye displacement, and thus a maximal fluorescence response in 96-well microplates with our supramolecular ODC tandem assay. This value is significantly lower than the reported limits of detection for colorimetric and electrochemical detection ( Table 1 ). Using 384-well microplates allowed further miniaturization, to a working volume of 10 µL. A maximal fluorescence response under these conditions is achieved with only 5 pmol putrescine (Z′ = 0.90), an amount that could even be lowered to 0.5 pmol (Z′ = 0.56, which is still sufficient for HTS). This places our supramolecular tandem assay among the most sensitive methods reported so far. Furthermore, the assay easily tolerates the presence of 1% DMSO, which is the common solvent for the addition of compounds to be tested ( Suppl. Figs. S4 and S5 ).
Limits of Detection of Various ODC Assays and Comparison of Desirable Performance Features of Functional and HTS Upscalable Assays. a

LOD, limit of detection; LC/MS, liquid chromatography–mass spectrometry.
Defined as the minimum detectable amount of the enzymatic reaction product putrescine.
n.d. = no limit of detection has not been reported.
384-well format.
In conclusion, we have introduced a label-free method for the real-time, continuous monitoring of ODC activity by fluorescence spectroscopy. The assay requires simply the addition of the commercially available host molecule CB6 and the fluorescent dye DSMI to the reaction mixture. We have shown that the resulting supramolecular tandem assay is suitable for determination of enzyme kinetic parameters and inhibition constants. In addition, we have demonstrated the potential of the assay in HTS applications and its sensitivity by using a 384-well microplate format. The novel supramolecular tandem assay for ODC thus presents a rapid, simple, and affordable method to allow its routine application, for example, for screening of drug candidates in HTS or for medical diagnosis of ODC levels as a disease marker. In direct comparison with other established assays, the supramolecular tandem assay presented herein ranks among the most sensitive methods; its simplicity is unmet, and it is the only ODC assay suitable for HTS ( Table 1 ). We believe that this assay may even be transferable to live cells.
Footnotes
Supplementary material is available online with this article.
Declaration of Conflicting Interests
The authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The authors disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This work was in part supported under the EURhythDia collaborative project funded by the European Commission (FP-7, grant agreement no. 278397) and by the DFG (HE 5967/4-1 and NA 686/11-1).
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.