
Abstract
Background
Anxiety disorders are the most common neuropathologies worldwide, but the precise neuronal mechanisms that underlie these disorders remain unknown. The hippocampus plays a role in mediating anxiety-related responses, which can be modeled in rodents using behavioral assays, such as the elevated plus maze. Yet, the molecular markers that underlie affect-related behavior on the elevated plus maze are not well understood.
Methods
We used herpes simplex virus vector delivery to overexpress extracellular signal-regulated kinase-2, a signaling molecule known to be involved in depression and anxiety, within the dorsal hippocampus of adult Sprague-Dawley male rats. Three days post virus delivery, we assessed anxiety-like responses on the elevated plus maze or general locomotor activity on the open field test.
Results
When compared to controls, rats overexpressing extracellular signal-regulated kinase-2 in the dorsal hippocampus displayed an anxiolytic-like phenotype, per increases in time spent in the open arms, and less time in the closed arms, of the elevated plus maze. Furthermore, no changes in locomotor activity as a function of virus infusion were observed on the open field test between the experimental groups.
Conclusion
This investigation demonstrates that virus-mediated increases of extracellular signal-regulated kinase-2 signaling, within the hippocampus, plays a critical role in decreasing anxiogenic responses on the rat elevated plus maze. As such, our data provide construct validity, at least in part, to the molecular mechanisms that mediate anxiolytic-like behavior in rodent models for the study of anxiety.
Introduction
Anxiety disorders constitute the most common forms of psychiatric illnesses worldwide.1,2 These disorders are associated with increased health-care costs and exhibit high comorbidity with other mental diseases, such as depression and posttraumatic stress disorder.3,4 Despite their high prevalence, the specific neurobiological mechanisms underlying these affect-related neuropathologies remain to be fully understood.
Members of the mitogen-activated protein kinases (MAPKs) function by integrating extracellular signals received by membrane receptors and transferring them through a phosphorylation cascade into the nucleus, thus regulating gene expression. 5 Although they are ubiquitously expressed, MAPKs have been found to play critical functions in the mammalian brain, ranging from synaptic plasticity, long-term potentiation, and neuronal differentiation. 6 Within the MAPK superfamily, the actions of members of the subfamily of extracellular signal-regulated kinase (ERK), particularly the ERK1 and ERK2 isoforms, have been extensively characterized in the central nervous system, and it is becoming increasingly clear that they play a prominent role in several psychiatric diseases, including depression and anxiety disorders.7–10
Preclinical pharmacological experiments have been useful in establishing a link between anxiety-related behavior and the ERK signaling pathway.11–13 However, despite significant efforts, there are currently no pharmacological agents that directly target individual and/or specific ERK isoforms for activation or inhibition,14,15 which in turn, limit our understanding on the role that each MAPK isoform plays in the regulation of mood-related behavior. Addressing this issue, we have previously shown that the upregulation of ERK2, via viral vector delivery in the dorsal hippocampus, reduces despair-related behavior in male rats, as determined in the forced swim test. 16 Conversely, stress-induced decreases of ERK-related signaling within this brain region has been linked to enhanced anxiety-like behavior, 17 which can be successfully studied in rodents using the elevated plus maze (EPM)—a behavioral paradigm with high face and predictive validity.18–20 Yet, to date, the precise role that ERK2 plays in the regulation of anxiety-related behavior has not been clearly delineated. Therefore, in order to determine if the upregulation of hippocampal ERK2 signaling is involved in the modulation of anxiety-like behavior, we combined a targeted molecular approach of ERK2 overexpression in the rat dorsal hippocampus, with behavioral assessment using the EPM and open field tests.
Materials and Methods
Animals
Male Sprague-Dawley rats (250–275 g) were obtained from Charles River Laboratories (Hollister, CA). Rats were maintained in a humidity- and temperature-controlled animal facility with a 12-h light/dark cycle and were subjected to a one-week acclimation period before surgery and the start of experiments. Rats were housed in pairs in standard laboratory polycarbonate cages (Ancare, model R20, Bellmore, NY) along with sani-chip bedding (P.J. Murphey San-Chips®, Murville, NJ) and had access to food and water ad libitum. Animals were cared for in accordance with the National Institutes of Health Guide for the Care and Use of Laboratory Animals 21 and with approval from the Institutional Animal Care and Use Committee.
Virus Vectors
Overexpression of ERK2 was achieved by microinjecting herpes simplex virus (HSV) vectors encoding green fluorescent protein (GFP)-alone or a GFP-wild-type ERK2 (wtERK2) in the dorsal hippocampus. Virus was packaged in a p1005 plasmid with expression of the target gene (i.e., ERK2) driven by an HSV IE 4/5 promoter, while GFP expression was driven by a CMV promoter. 22 All viruses were diluted in phosphate-buffered saline (PBS) + 10%sucrose and 20 mM HEPES (pH 7.3). For further specifics on vector design, construction, neurotropic effects, as well as in vitro and in vivo validation, see previously published work.23,24 Highest levels of transgene expression by the HSV vectors were reached three days after surgery. Therefore, behavioral experiments began after this time period (see Figure 1(a) for experimental design).25,26 Transgene expression was restricted to an area of ∼1 mm2 surrounding injection site, as previously demonstrated.27,28 Behavioral tests (described below) were recorded using a video tracking system (Ethovision XT, Noldus, Leesburg, VA).
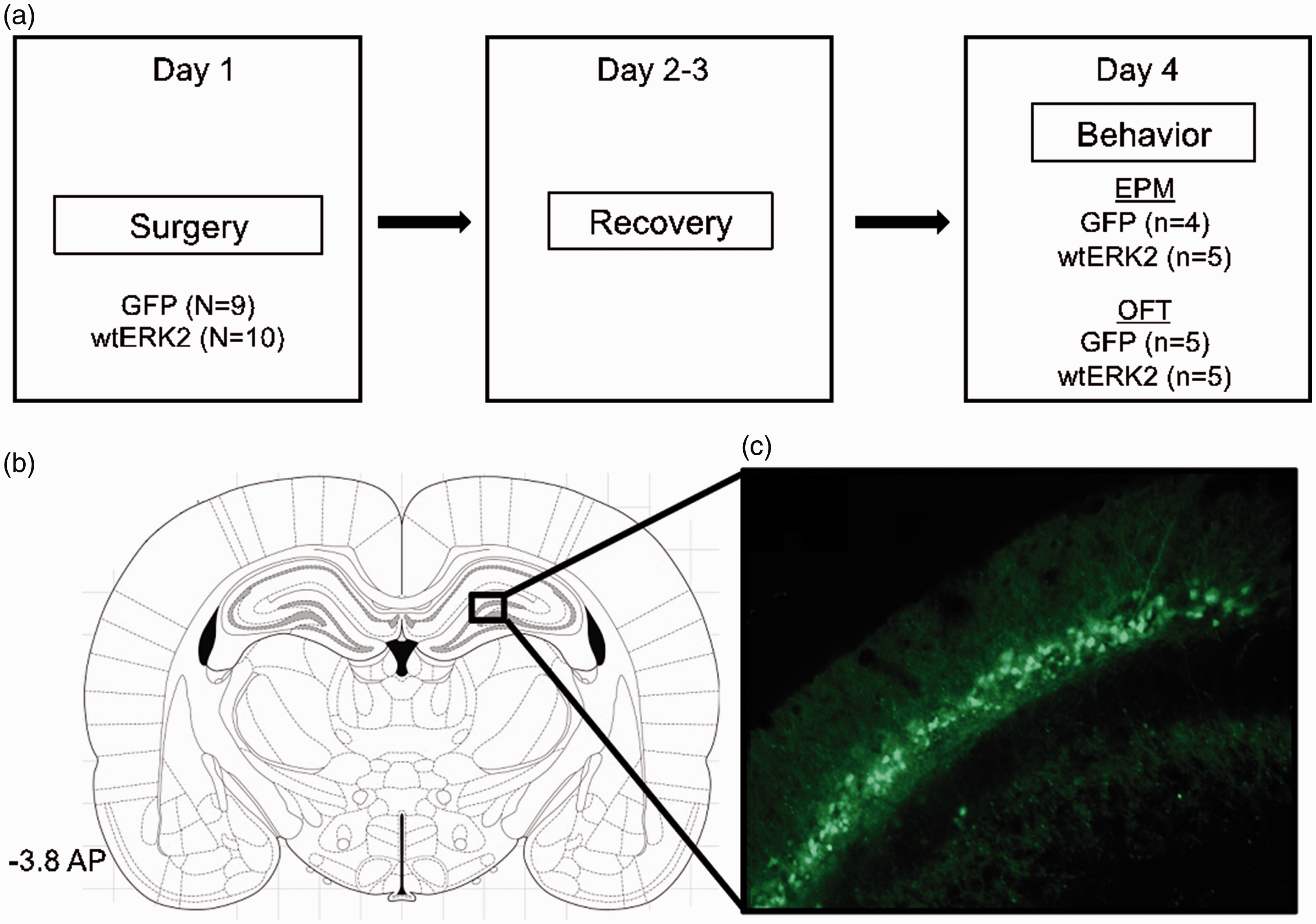
Experimental design and virus-mediated overexpression of ERK2 in the rat dorsal hippocampus. (a) Male rats were infused with HSV vectors overexpressing GFP-alone or a GFP-wtERK2 on day 1 (surgery) and allowed to rest for 48 h (days 2–3). On day 4, rats were tested on the EPM or the OFT. (b) Region of the hippocampus to which microinjections of HSV vectors was targeted (AP: −3.8, lateral: −2, dorsoventral: −3.2 mm below dura). Adapted from The Rat Brain in Stereotaxic Coordinates. 53 (c) Cells expressing GFP-wtERK2 (green, cyanine 2) fluorescence (magnification, ×400). GFP: green fluorescent protein; EPM: elevated plus maze; OFT: open field test; AP: anteroposterior; wtERK2: wild-type extracellular signal-regulated kinase-2.
Animal Surgery
Stereotaxic surgeries were conducted for selective delivery of the HSV vectors. Rats were anesthetized with an intramuscular injection of ketamine/xylazine cocktail (80/10 mg/kg), with subcutaneous atropine (0.25 mg/kg) administered to reduce bronchial secretions; bilateral microinjections of either GFP or GFP-wtERK2 vectors were then administered (1 µl per hemisphere over a 10-min period) into the dorsal hippocampus (coordinates from Bregma: anteroposterior: −3.8, lateral: −2, dorsoventral: −3.2 mm below dura) using a 32-gauge Hamilton syringe.16,29 Postsurgical discomfort was minimized by applying the local anesthetic bupivacaine.
Elevated Plus Maze
The EPM was used to evaluate anxiety-like behavior. 30 The maze consisted of two perpendicular, intersecting runways (12 cm wide × 100 cm long): one with tall, closed arms (30 cm), and the other with open arms, connected by a central area. The maze was elevated 1 m from the floor in a behavioral testing room under controlled lighting (∼90 lux) conditions. Rats were placed initially in the central area, facing one of the open arms, and grooming counts, as well as the cumulative time spent in the closed and open arms was recorded for 5 min. 31
Locomotor Activity
Locomotor activity of GFP-alone or GFP-wtERK2 rats was assessed as distance traveled (cm) in an open field apparatus (63 × 63 × 26 cm) for 5 min.
Histology and Transgene Detection
The sites of HSV vector injection were confirmed in all rats. Animals were euthanized with an overdose of sodium pentobarbital administered 1 h after behavioral assessment, followed by transcardial perfusion with 0.9% saline and 4% paraformaldehyde (PFA). Brains were excised and postfixed by overnight immersion in 4% PFA and stored in 20% glycerol. Hippocampal coronal sections (45 µm) were obtained on a microtome and stored in 0.1 M sodium phosphate buffer containing 0.05% sodium azide. Tissue sections were used to examine GFP expression within the hippocampus as previously described. 32 Hippocampal free-floating coronal sections were blocked in 3% normal donkey serum (NDS), followed by overnight incubation with primary antibody (rabbit anti-GFP antibody, 1:1000; Abcam, Cambridge, MA) along with 0.3% Triton X-100 and 1% NDS. Tissue was then incubated with anti-rabbit secondary antibody (1:1000; Jackson ImmunoResearch, West Grove, PA) for 2 h at room temperature. Immunostained sections were mounted on microscope slides, dehydrated in ethanol and Citrosolv, and coverslipped with DPX (Sigma, St. Louis, MO). Slides were visualized and photographed using a confocal fluorescence microscope equipped with a digital camera.
Statistical Analysis
Rats were randomly assigned to receive either HSV-GFP-alone or HSV-GFP-wtERK2 vectors, three days prior to behavioral testing. Statistical testing was carried out using two-tailed Student’s t test. Data are expressed as mean ± standard error of the mean. A value of p < 0.05 was adopted to determine statistical significance.
Results
Figure 1(b) depicts the hippocampal region targeted for microinjection delivery of HSV vectors. As observed in Figure 1(c), immunofluorescence confocal microscopy revealed that ERK2 was successfully overexpressed in the dorsal hippocampus as previously shown.16,27
Using the EPM test to determine anxiety-like behavioral responses to GFP-wtERK2 overexpression in the dorsal hippocampus, we found that, when compared to their GFP-only counterparts (n = 4), GFP-wtERK2 (n = 5) rats spent significantly more time in the open arms (t7 = 2.78, p < 0.05; Figure 2(a)) and less time in the closed arms (t7 = 3.03, p < 0.05; Figure 2(b)) of the EPM. No changes were observed in grooming counts (t7 = 0.26, p > 0.05; Figure 2(c)). Lastly, Figure 2(d) shows that hippocampal overexpression of ERK2 (n = 5) did not influence locomotor activity as measured in the open field test when compared to GFP-only rats (n = 5; t8 = 1.12, p > 0.05).

Herpes simplex virus overexpression of ERK2 (wtERK2) mediates an anxiolytic-like effect in the elevated plus maze. When compared to controls (GFP, n = 4), male rats overexpressing wtERK2 (n = 5) in the dorsal hippocampus spent higher time in the open arms (a), along with decreased time in the closed arms (b), of the maze. No differences in grooming counts (c) or distance traveled (d, n = 5 per group) were noted between the experimental groups. *p < 0.05. GFP: green fluorescent protein; wtERK2: wild-type extracellular signal-regulated kinase-2.
Discussion
In this study, we aimed to determine the behavioral effect of viral vector-mediated ERK2 overexpression in the dorsal hippocampus of adult male Sprague-Dawley rats, using the EPM as a tool to assess anxiety-related behavior. 30 Our results indicate that rats overexpressing GFP-wtERK2 in the dorsal hippocampus spent more time in the open arms and less time in the closed arms of the EPM, without overall changes in locomotor activity. These behavioral responses are consistent with an anxiolytic effect, 18 a behavioral outcome that is commonly reported after acute benzodiazepine (BDZ) exposure,33,34 or chronic administration of selective serotonin reuptake inhibitors (SSRIs).31,35 Of note, these two classes of drugs, which are used clinically to treat anxiety-related disorders, 36 have been reported to mediate their therapeutic effects via neuronal ERK signaling regulation. Specifically, treatment with the BDZ drug diazepam has been shown to increase the levels of ERK1/2 in the rat hippocampus, 37 and the administration of the SSRI fluoxetine similarly enhances ERK1/2 signaling across different brain regions, 38 including the hippocampus 39 ; brain circuitry that is activated during both stress and anxiety responses. 40 Likewise, activation of the ERK pathway via treatment with brain-derived neurotrophic factor results in elevated levels of activated ERK2 as well as reduced anxiety-like behavior in mice. 41 Conversely, pharmacological inhibition of hippocampal ERK signaling in rats has been shown to induce anxiety-like behavior in the EPM. 11 However, we must note that the role of ERKs, as a molecular link between stress and anxiety, is equivocal across the literature.42–45
While previous work has suggested that BZD and SSRI medications mediate their therapeutic effects via increases in hippocampal ERK1/2 signaling, our experimental approach confirms that the upregulation of ERK2 specifically, in the stress-naïve rat’s dorsal hippocampus, is a contributor in the modulation of anxiety (Figure 2) and despair-like behavior. 16 Importantly, we do so by using a genetically targeted approach, providing construct and mechanistic validity to the male rat EPM test. Furthermore, our findings expand on recent work indicating that the ventral hippocampus also modulates behavioral responses in the EPM46,47—however, whether anxiolytic behavior in this paradigm is ERK2-dependent, within the ventral portion of the hippocampus, remains to be explored. Unfortunately, a limitation of the current investigation is that we did not include female rats in our experimental design, reducing the interpretability of our findings to the clinical setting, where anxiety-related disorders are more frequently diagnosed in the female, versus male, population. 48 Lastly, given that ERK1 and ERK2 share over 80% homology, it will be important to evaluate how selective regulation of ERK1 influences responses on the EPM, since ERK1-alone has been found to modulate behavior differentially in other tasks.49–51
Conclusion
The neurobiological correlates between stress and anxiety are not well understood. Previous animal and human postmortem studies have shown that ERKs, across different brain regions,10,16,23,52 play a crucial role in modulating affect-related responses. Here, we expand this work to include a role for ERK2 specifically, within the dorsal hippocampus of rats, in mediating anxiolytic behavior when assessed on the EPM—a classic model used to screen for anxiolytic or anxiogenic compounds. Indeed, virus-mediated increases of ERK2 resulted in animals spending more time in the open arms, and lower time in the closed arms, of the EPM; an anxiolytic response mimicking the effects of traditional pharmaceutical agents used for the management of anxiety (i.e., SSRIs and BDZs). As such, our data may prove insight for the development of pharmaceutical agents, or alternative strategies, to treat/prevent the development of anxiety-related disorders.
Footnotes
Acknowledgment
The authors thank Dr. Eric J. Nestler for donating the virus vectors used in this investigation.
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This work was supported by a grant from the National Institute of General Medical Sciences (SC3GM130467) awarded to S. D. I. Additionally, J. A. S.-F. was supported through a postdoctoral training contract from the National Institute on Drug Abuse (HHSN271201600057C). F. J. F.-R. was supported by a Summer Research Grant from the Graduate School at The University of Texas at El Paso (UTEP). I. G.-C. was funded by the LSAMP (Louis Stokes Alliance for Minority Participation) Bridge to Doctorate support program at UTEP.