
Abstract
Importance of the neuroendocrine brain for health and happiness has become clear since the 1960s. Foundations laid 100 years ago culminated in Geoffrey W Harris’s model of control by the brain of secretion of anterior and posterior pituitary gland hormones through, respectively, releasing factors secreted into the hypothalamic-hypophysial portal system, and directly from axon terminals into the systemic circulation. Confirmation, expansion and deepening of knowledge and understanding have followed increasingly sophisticated technology. This allowed chemical characterisation of the posterior pituitary hormones, oxytocin and vasopressin, the releasing factors, their receptors and genes, location of the neurosecretory neurons in the hypothalamus, and how their activity is controlled, including by neural and hormonal feedback, and how hormone rhythms are generated. Wider roles of these neurons and their peptides in the brain are now recognised: in reproductive and social behaviours, emotions and appetite. Plasticity and epigenetic programming of neuroendocrine systems have emerged as important features.
Keywords
What neuroendocrinology is: overview of neuroendocrine organisation and mechanisms now
Neuroendocrinology was initially just about studying the control by the brain, via the hypothalamus, of the secretion of hormones by the posterior pituitary gland (oxytocin, vasopressin), and by the anterior pituitary gland (adrenocorticotropic hormone (ACTH); the gonadotropins – follicle stimulating hormone (FSH) and luteinising hormone (LH); growth hormone; prolactin; thyroid stimulating hormone (TSH)), the adrenal medulla (adrenaline) and the pineal gland (melatonin). The neuroendocrine brain comprises discrete sets of hypothalamic neurons, and their rich neural inputs, that either project to the posterior pituitary (oxytocin and vasopressin neurons) or the median eminence (releasing factor neurons) where their axon terminals abut blood capillaries. The former secrete into the systemic circulation (so these magnocellular neurons have large cell bodies owing to their necessary large capacity for peptide synthesis and secretion) and the latter into the primary capillaries of the hypothalamic-hypophysial portal vessel system (these neurons are small, parvocellular, reflecting need for production of less stored peptide) (Figure 1). Geoffrey Harris first proposed and demonstrated the key and essential importance and function of this portal system in the regulation by the brain of the stimulation or inhibition of secretion by specific types of anterior pituitary cells into the systemic circulation (Harris, 1955). This proposal was supported by contemporaneous studies with neuropharmacology, electrical stimulation and anterior pituitary transplantation, showing important roles of the hypothalamus in controlling gonadal function (Everett and Sawyer, 1950; Markee et al., 1946; Nikitovitch-Winer and Everett, 1957); the demonstration of hypophysiotropic areas of the hypothalamus by using anterior pituitary implants and selective micro-surgical de-afferentation within the hypothalamus (Szentágothai et al., 1968); and finding that extracts of the hypothalamus had actions expected of the proposed releasing factors, and that these were peptides (Guillemin and Rosenberg, 1955; McCann, 1962). This set the scene for precise characterisation of hypothalamic releasing factors, beginning in the 1970s.

Neurosecretory neurons in the hypothalamus. (a) Location of oxytocin and vasopressin neurons. Main: one side of a coronal section of a rat brain. PVN: paraventricular nucleus; SON: supraoptic nucleus; V3: third ventricle; OC: optic chiasm. Oxytocin (OXT) and vasopressin (VP) neurons are identified by fluorescent immunocytochemistry: oxytocin neurons are red and vasopressin neurons are green (top inset); both types of neuron are in the PVN and SON. The SON comprises only magnocellular neurons, projecting to the posterior pituitary gland. In the PVN, most magnocellular OXT and VP neurons are grouped laterally; the medial PVN contains mostly parvocellular neurons (Figure 1b). Inset bottom: electron microscopy photomicrograph of dendrites in the ventral SON (dashed oval outline/white arrow) showing secretory granules (red arrow) and an exocytotic profile (yellow arrow). (b) Distribution of neurosecretory neurons in the paraventricular nucleus. The PVN contains multiple types of peptidergic neurons, predominantly neuroendocrine neurons projecting to the posterior pituitary (magnocellular, mne in key) or to the median eminence, secreting into the primary capillaries of the hypothalamic-hypophysial portal system (parvocellular, pne in key) and identified by axonal terminal uptake and retrograde transport of intravenously injected fast blue (from the median eminence or posterior pituitary respectively). Each coloured dot represents the cell body of different peptidergic types of neurosecretory neuron, identified by simultaneous multiple immunocytochemistry; numbers of each type in this plane of section are in the key. Key: TRH: thyrotropin releasing hormone, SS: somatostatin, TH: tyrosine hydroxylase (i.e. dopaminergic – prolactin inhibiting factor (PIF), most are in the arcuate nucleus); CRH: corticotropin releasing hormone; VP: vasopressin; OXT: oxytocin; GRH: growth hormone releasing hormone (none in this plane, most are adjacent to the arcuate nucleus in the ventral hypothalamus). The PVN also contains non-neuroendocrine (i.e. centrally projecting) neurons of each of these types, especially CRH, OXT, SS and TRH neurons. 26A: section level in Simmons and Swanson (2009).
In this epoch, the amino acid sequences and genes for the anterior pituitary hormones were established, which included showing that ACTH is produced from a precursor, pro-opiomelanocortin (POMC), which contains β-endorphin and α-melanocyte stimulating hormone (α-MSH; processed from ACTH) and other active peptides (Lowry, 2016), and importantly is expressed in the neuroendocrine brain.
Scope of neuroendocrinology
This embraces major body functions – reproduction, growth and metabolism, water and electrolyte balance, diurnal and seasonal rhythms and resilience to stress. Control by the brain has three elements – receiving external environmental signals, in all sensory modalities, and then organising neuroendocrine responses appropriate to meet challenges (stressors), to accommodate opportunities (to eat, drink, reproduce) and to harmonise with the natural day and seasons on planet earth; processing internal signals to maintain or restore homeostasis through neuroendocrine adjustments (osmoregulation, balancing energy supply and use, or coping with infection (Kentner and Pittman, 2010)); detecting hormonal signals from peripheral endocrine glands the brain regulates that mainly act as negative (but some can be positive) feedback signals which automatically adjust the neuroendocrine output from the brain to keep the output from the controlled endocrine gland within set limits. Feedback may be on the anterior pituitary as well as on neurons in the brain.
Boosting feedback signals
The neuroendocrine brain is not a passive receiver of peripheral endocrine signals: considering steroid and thyroid hormones, the brain has enzyme mechanisms for amplifying or transforming these signals. Thyroxine (T4), the classical thyroid hormone, is activated by partial deiodination to triiodothyronine (T3) in target tissues, and the brain also expresses the deiodinase enzymes; their expression is regulated by T3 as a way of enhancing the feedback signal. Enzymes that interconvert sex steroid hormones are expressed in the brain and are important in the early developing male brain by converting testosterone to 17β-estradiol, which is the form that induces masculinisation and defeminisation of key control networks. The brain can convert progesterone, and other steroids, to forms (e.g. allopregnanolone) that are neuroactive – acting on neuronal membranes to alter electrical excitability. Glucocorticoids (cortisol in humans, corticosterone in laboratory rodents) are metabolised and inactivated in the periphery, by 11β-hydroxysteroid dehydrogenase 2 (11β-HSD 2): the brain is among tissues that can reactivate the metabolite, via action of 11β-HSD 1 (Harris et al., 2001), which has been shown to have implications for understanding stress effects on the brain (Sarabdjitsingh et al., 2014).
Importance
Disordered regulation of neuroendocrine systems, owing to problems within the brain, or damage to the pituitary or a peripheral endocrine gland, or an endocrine tumour over-secreting a hormone, can have serious life-threatening consequences. Indeed, such conditions have provided impetus for investigating neuroendocrine systems, and in turn such understanding is key to diagnosis and treatment.
Rhythms
A feature of neuroendocrine systems is that they generate rhythms, related to external rhythms of day and night or season, or to induce ovulatory cycles in females, or as a consequence of the intrinsic behaviour of neuroendocrine neurons, or as an emergent feature of feedback mechanisms. An early finding after gonadotropin-releasing hormone (GnRH) was characterised was that GnRH secretion into portal blood is pulsatile and surges at the critical period for stimulation of ovulation (Sarkar et al., 1976). Secretion of hormones in pulses evidently avoids target desensitisation and permits effects to be efficiently achieved through transient peak levels of hormone. While neuroendocrine neurons generate rhythms via the patterning of their electrophysiological activity, feedback from rhythmic activity in target gland activity can complicate the rhythmicity generated by mechanisms in the brain.
Wider and wider
But neuroendocrinology is about more than all this. Beyond central actions of hormones that are produced peripherally to regulate secretion of these hormones, hormone actions on the brain include triggering behaviours and emotions (mood and desires) that are complementary to other actions of the hormone. Examples are the central actions of sex steroids necessary for sex behaviour in adulthood and in determining gender during critical periods in early life; actions of glucocorticoids secreted during stress on the hippocampus that aid memory, but may also be deleterious, and also anxiogenic actions on the amygdala that can be interpreted as evoking coping responses. Now included are the actions of several peptides from adipose tissue, the gastro-intestinal tract and pancreas, and immune cells, which signal aspects of metabolic state and affect appetite and neuroendocrine systems that regulate metabolism.
Central actions of hormones
As for the target cells of peripheral hormones, actions in the brain and anterior pituitary are mediated by specific hormone receptors. The lipophilic properties of steroid hormones mean they readily cross the blood-brain barrier, and there are transporters for thyroid hormones; then they enter cells, bind to a specific cytoplasmic receptor (these are essentially transcription factors), enter the nucleus and modify (stimulate or repress) expression of specific genes, eliciting the specific hormone action on the cells, including neuroendocrine neurons. Faster steroid hormone actions on electrical excitability of neurons can be through receptors expressed on the cell membrane. Peptide hormones act via specific membrane 7-transmembrane (7-TM) receptors, typically coupled to G-protein (GPCR) and an adenylyl cyclase, cAMP, pathway, or via a specific single transmembrane cytokine family receptor and the JAK-STAT pathway.
Unlike steroid hormones, most peptide hormones cannot cross the blood-brain barrier, so their actions may be via circumventricular organs, which have fenestrated capillaries, hence some blood-borne peptides can act here (e.g. angiotensin, relaxin, interleukin-1β, ghrelin) if there are adjacent neurons expressing the appropriate receptors; for some other peptides (e.g. leptin, prolactin), there are also selective transport mechanisms in the choroid plexus that enable entry into the CSF. For yet others, their actions can be at the site of peripheral release, on receptors on afferent nerve terminals (e.g. cholecystokinin (CCK) and vagus nerve), or indirect through excitation of a peripheral target which in turn excites afferent innervation (e.g. oxytocin and myometrial contractions); clearly, such peripheral actions must act via central projections of the afferent neural pathway, which may be via brainstem monoaminergic and/or peptidergic systems.
Change means change
In general, every component of neuroendocrine systems can be a site of plasticity that affects activity and responsiveness; changes may involve refinements of circuitry in the neural networks in which neuroendocrine neurons are embedded. Plasticity may be part of a normal step at different stages of life (e.g. puberty, pregnancy) or be a consequence of a dangerous disturbance of homeostasis (stress) and its costly correction (allostasis; McEwen and Wingfield, 2003). This encompasses up- and down-regulation of receptor effectiveness, which may be due to ligand-induced desensitisation or altered expression; such plasticity is important in understanding normal and abnormal physiology. The specific signalling molecules that neuroendocrine neurons produce impart their secretory phenotype, but for a particular phenotype, this can change qualitatively and quantitatively in typical ways as a result of physiological state. Epigenetic mechanisms (DNA methylation, histone acetylation) may underlie such plasticity.
The neurohypophysis (magnocellular neurons + posterior pituitary) – an exemplary neuroendocrine system
Neurosecretion
The seed for neuroendocrinology was sown at the beginning of the last century with the discovery that the posterior pituitary gland, a midline ventral down-growth from the brain, contains active substances that stimulate uterine contractions and release of milk from the lactating mammary gland, and increase blood pressure (Dale, 1906; Ott and Scott, 1910), and excretion of concentrated urine (Farini, 1913; Heller, 1941; van der Velden, 1913). These activities were assigned to presence in the posterior pituitary of two hormones – oxytocin (acting on myometrium and mammary gland to cause ejection of fetus(es) and milk, respectively) and vasopressin (causes vasoconstriction, increasing blood pressure), also called antidiuretic hormone (ADH). There was initial confusion about presence in the posterior pituitary of diuretic as well as antidiuretic activities but the prime importance of ADH was resolved by Verney (1946a), who also demonstrated the role of osmoreceptors in the brain, supplied by the internal carotid arteries, that mediate stimulation of ADH secretion by increased blood osmolality (Verney, 1946b).
It took 50 years from the initial discoveries to confirm that oxytocic and vasopressor/antidiuretic activities are exerted by two distinct hormones in the posterior pituitary, through ground-breaking sequencing of the two peptides, each with nine amino acids, synthesising them, and confirming their distinctive biological activities (du Vigneaud et al., 1954a, 1954b): oxytocin was the first peptide hormone to be synthesised.
Comparative anatomical, histological and electron microscopy studies showed that the posterior pituitary is innervated by axons of neurons in the supraoptic nuclei (SON) and paraventricular nuclei (PVN) of the hypothalamus and that these relatively large, magnocellular, neurons contain abundant protein synthesising machinery and membrane bound granules like other secretory cells, including neurons in invertebrates (Scharrer and Scharrer, 1945). Hence, these neurons were designated as neurosecretory and deduced to produce oxytocin and vasopressin in their cell bodies and then to transport these hormones in their axons to the posterior pituitary and when electrically excited secrete the peptides by exocytosis from the thousands of axon terminals into fenestrated capillaries in the gland, thus entering the systemic circulation.
Oxytocin and vasopressin neurons
The magnocellular oxytocin and vasopressin neurons, found in the PVN and SON, are regarded as exemplar neuroendocrine cells with properties that other neurosecretory neurons should be expected to match (Figure 2): their cell bodies are in the hypothalamus; the neurons have excitatory and inhibitory synapses on their dendrites and soma, and appropriate specific receptors; the neurons produce and secrete peptides that mediate their functions (except that tubero-infundibular dopaminergic neurons use dopamine); their axon terminals are on fenestrated capillaries (in the posterior pituitary for magnocellular neurons; in the median eminence for releasing factors/ hormones); secretion is stimulated by arrival of action potentials that increase Ca2+ entry into the terminals, triggering exocytosis; frequency and pattern of action potentials generated in the cell bodies determine amount and pattern of peptide released; pre-terminal modulation of stimulus-secretion coupling is likely.

Features of a magnocellular oxytocin neuron (MCN): a model peptidergic neuron. Oxytocin is synthesised in the cell body, as a higher molecular weight precursor, translated from the mRNA transcribed from the OXT gene; precursor peptides are processed to oxytocin-neurophysin and packaged into secretory granules in the Golgi apparatus; secretory granules are moved by axonal transport to the axon terminals; some are transported into dendrites; precursor is processed further during transport; the axon terminals are on fenestrated capillaries in the posterior pituitary (some axon swellings release into portal capillaries in the median eminence, like releasing factors – oxytocin can stimulate prolactin secretion); a few magnocellular oxytocin neuron axons (dashed) project centrally (e.g. to amygdala). Secretion is via exocytosis, stimulated by action potentials (APs) that increase Ca2+ entry; frequency and pattern of action potentials generated in the cell bodies determine amount and pattern of peptide released; stimulus-secretion coupling can be modulated by pre-terminal action of κ-opioid peptide. Under most conditions, oxytocin neurons fire APs irregularly but during parturition or suckling they fire in brief high-frequency bursts, co-ordinated among the oxytocin neurons, resulting in secretion of a pulse of oxytocin after each burst. Major excitatory inputs use noradrenaline (NA) or glutamate (GLU); major inhibitory input is via GABA; allopregnanolone (AP; progesterone neurosteroid metabolite) potentiates GABA action via GABAA receptors; more than 20 neuropeptides have excitatory or inhibitory (e.g. opioid) actions on magnocellular oxytocin neurons. Dendritic secretion of oxytocin drives bursts – auto-stimulating endocannabinoid (CB) release, which presynaptically modulates inputs (GLU); placental leucine aminopeptidase (PLAP) from oxytocin dendrites limits local oxytocin action; ATP/adenosine (Ad) and nitric oxide (NO) from dendrites also have local inhibitory actions.
The hypothalamus and adenohypophysis
Harris’s hypothesis about how secretions of the adenohypophysis (anterior pituitary gland) are controlled by the brain through the secretion by specific neurons in the hypothalamus of releasing factors into the hypothalamic-hypophysial portal system has been amply confirmed by many studies since. Confirmation came especially from the first chemical characterisation of releasing factors/hormones (for TSH – TRH, and for FSH and LH – LHRH/GnRH) by Roger Guillemin and Andrew Schally, who used extracts of hundreds of thousands of sheep and porcine hypothalami from slaughterhouses to achieve this (Amoss et al., 1971; Burgus et al., 1970; Schally et al., 1971). The chemical identification of CRH (Vale et al., 1981), GHRIH (somatostatin; Brazeau et al., 1973) and GHRH (identified in a pancreatic tumour extract; Guillemin et al., 1982; Rivier et al., 1982) followed (McCann, 1992). Discovery of stimulatory and inhibitory factors regulating GH secretion and the patterning of their secretion into the portal system explained the periodic pattern of GH secretion as resulting from intermittent lifting of central inhibition of GHRH neurons by somatostatin (Plotsky and Vale, 1985).
Identification of the releasing factors enabled use of immunocytochemistry (ICC) and in situ hybridisation (ISH) to locate, and visualise the neurosecretory neurons that produce these neuropeptides (Figure 1b).
Further progress since the 1960s: novel technologies and new paradigms
Hormone characterisation
A basic hypothesis is that there can be a releasing hormone (RH or factor RF) and a release inhibiting factor (RIF or RIH) for each anterior pituitary hormone, with appropriate specific receptors on the respective anterior pituitary trophic cells. Most of these are produced by parvocellular neurons in the PVN (Figure 1b). Specific membrane receptors have been identified for each RH or RIH, but to date, only GH is clearly regulated by both a GHIH (somatostatin 14/28 a.a.) and by a GHRH (44 a.a. peptide); for ACTH/Corticotropin, CRH (a 41 a.a. peptide) and vasopressin (9 a.a.) stimulate ACTH secretion; LH and FSH (gonadotropins, Gn) are stimulated by the same peptide (GnRH = LHRH, decapeptide), and a proposed LHRIF (RF-amide related peptide-3, RFRP-3) (Clarke et al., 2016), has not yet been fully validated (George et al., 2017b); for TSH, TRH tripeptide (the first releasing factor to be characterised) stimulates secretion; prolactin secretion is inhibited by dopamine (acting as a PIF) and can be stimulated by oxytocin or prolactin releasing peptide (PrRP, 31 a.a. peptide; Hinuma et al., 1998), though PIF is predominant, and it is now clear that PrRP primarily functions within the brain, including in the context of energy homeostasis (Dodd and Luckman, 2013). For ACTH, gonadotropins and TSH the target gland hormones (steroids and thyroxine) act as inhibitory regulators.
An important finding was the discovery that continuous treatment with GnRH down-regulates the GnRH receptor on gonadotrophs, removing the drive to sex steroid secretion (Zilberstein et al., 1983): so GnRH agonist analogues are important in treatment of androgen-dependent cancer, and precocious puberty.
Protein and gene expression
As both oxytocin and vasopressin have a cysteine-cysteine ring, in vivo synthesis of the oxytocin and vasopressin precursor peptides and axonal transport and storage could be studied by local injection of S35-labelled cysteine, or other radio-labelled constituent amino acids, into the PVN or SON (Brownstein et al., 1980; Cross et al., 1975; Sachs and Takabatake, 1964). Availability of specific antibodies has enabled key questions about synthesis and distribution of the peptide neurohormones, at the cellular and sub-cellular level, with the techniques of immunocyto/histo-chemistry with light and electron microscopy, and at the brain region level with western blotting. Isolation of the mRNAs and sequencing of the genes encoding the posterior pituitary peptides and the releasing hormones and their receptors provided tools to identify with ISH the anatomical distribution of expression in the brain and anterior pituitary, and to investigate dynamics of changes with functional demand and the mechanisms regulating expression of the genes.
New paradigm: hypothalamic-pituitary-thyroid (HPT) axis feedback
Tanycytes
Recent careful morphological and gene expression studies have indicated that the morphologically striking tanycytes lining the base of the third ventricle have a critical role in mediating negative feedback action of thyroid hormone on hypophysiotropic TRH neurons (Fekete and Lechan, 2014; Figure 3). They have long processes extending to the median eminence, arcuate nucleus, dorsomedial nucleus (DMN) or ventromedial nucleus (VMN); one group projects to the pars tuberalis of the pituitary. Tanycytes express thyroid hormone receptors, and TRH degrading enzyme, which is up-regulated by thyroid hormone, acting like a metabolic fire extinguisher, limiting TRH stimulation of TSH secretion and hence thyroid hormone secretion as part of negative feedback. They also have TSH receptors and TSH, from nearby pars tuberalis thyrotrophs, stimulates type 2 iodothyronine deiodinase (D2) expression, hence increasing local tri-iodothyronine (T3) concentrations (Bolborea et al., 2015). Tanycytes are considered to have a particular key role in seasonal and hibernating species, as they show increased D2 expression under long day conditions (Ebling, 2014). There is clearly more to learn about roles of tanycytes in metabolic regulation.

Tanycytes and thyroid hormone feedback to TRH neurons. Tanycytes (which stain for vimentin, green) line the base of the third ventricle (3V; shown here in a transverse section diagram), in 4 groups (α1 and 2, β1 and 2) sending processes into the hypothalamus, including the arcuate nucleus; β1 tanycytes project to the median eminence (ME) as far as the pars tuberalis. In the external ME zone, tanycyte processes are in close contact (small dashed oval) with pro-thyroid stimulating hormone releasing hormone (TRH) – immunopositive axon terminals (deep pink) abutting primary capillaries of the hypothalamic-pituitary portal vessel system, into which TRH diffuses after release from the axon terminals. The cell bodies of the TRH neurons are in the parvocellular paraventricular nucleus (pPVN), and receive several types of neural input. The large dashed oval details feedback interactions between tanycytes and TRH neurons: tanycytes are glucosensitive and have membrane transporters for thyroid hormone (T3 and T4, asterisks), and express type 2 iodothyronine deiodinase (D2) which converts T4 to T3 (active form of thyroid hormone), which is delivered to TRH neurons, either via CSF or retrograde axonal transport; tanycytes have cell surface pyroglutamyl peptidase II (PPII; red stars), inducible by T3, which degrades released TRH, attenuating the effects of stimulation of TRH neurons.
Hormone measurements
Measurements of activity of neuroendocrine systems were initially based on assessing outputs – changes in activity of anterior or posterior pituitary hormone targets, and careful, tedious bio-assays of hormone concentrations in peripheral blood, based on the physiological responses of target tissues, in vitro or in vivo, using international standard extracts or synthetic peptides to calibrate assays. Chemical measurements were developed to measure steroids and monoamines. In 1960, development of a radioimmunoassay method was announced (Yalow and Berson, 1960), which revolutionised measurement of specific peptide and steroid hormones. Refinement of the principle and commercial development has led to numerous types of assays based on the high affinity and specificity of antibodies for complex chemical moieties; though specificity is an issue, especially if measurements are attempted on un-extracted samples. The sensitivity and reproducibility of immunoassays meant that serial measurements in small samples of biological fluids became possible, providing information about dynamics of neuroendocrine responses. However, a limitation is that while hormones from the posterior and anterior pituitary and peripheral endocrine glands (including the gastro-intestinal tract, adipose tissue and immune cells) are present in the circulation at concentrations high enough for their actions and hence for immunoassay, hypothalamic releasing factors are at detectable concentrations only in the few µl of hypothalamic-hypophysial portal blood. Hence, surgical techniques were developed to sample portal blood to measure RH secretion (Clarke and Cummins, 1982; Sarkar et al., 1976), complemented by in vitro studies (Loxley et al., 1993).
Paradigm shift: peptide actions in the brain
The techniques for identifying neuropeptide production revealed that in addition to neurosecretory neurons projecting to the posterior pituitary and the median eminence, these or other neurons projecting within the brain produce and release the same peptides (Hökfelt et al., 1980; Landgraf and Neumann, 2004; Swanson and Sawchenko, 1980). Importantly, specific receptors for these neuropeptides were concurrently found in the brain. Thus, the known repertoire of neurotransmitters in the brain was vastly increased when neuropeptides were included. A working concept about the functions of these peptides in the brain is that they have actions in eliciting emotions and behaviours that are coherent with their neuroendocrine functions (e.g. oxytocin is essential for milk transfer and promotes maternal behaviour (Pedersen and Prange, 1979); CRH drives anterior pituitary-adrenocortical stress responses and is anxiogenic (Bale and Vale, 2004); gut and metabolic peptides alter appetite (Field et al., 2010). Routine screening of newly described neuropeptides for central actions on behaviours or neuroendocrine activity, selected on the basis of anatomical distribution in the brain, similarity to known peptides, and receptor binding properties is an important part of discovery (Wren et al., 2000).
Measuring activity of neuroendocrine neurons in real or historical time/time past
Real time in vivo
Geoffrey Harris and Barry Cross elegantly showed that electrical stimulation of the neural stalk of the posterior pituitary gland in vivo caused oxytocin secretion, as indicated by autobioassay with milk ejection the endpoint (Cross and Harris, 1952). The group led by Cross showed that magnocellular neurons, identified by antidromic stimulation of the neural stalk, discharge action potentials like other neurons and that they fire with an irregular or an asynchronous phasic pattern of activity (Cross et al., 1975). Functional tests showed a phasic pattern is typical of vasopressin neurons (Dreifuss et al., 1976); during suckling the type firing irregularly dramatically and synchronously discharged brief high-frequency bursts with a milk ejection following each burst, so these neurons were characterised as oxytocinergic (Wakerley and Lincoln, 1973; Figure 2). As the secretion of many hormones is rhythmic, the burst firing of oxytocin neurons provides a paradigm to explain neuroendocrine rhythms in general at the level of RH neurons. However, such neuronal rhythms have been difficult to show directly, but have been inferred from measuring rhythms in RH concentrations in portal blood (Sarkar et al., 1976). Detailed in vitro study of the mechanisms of secretion of posterior pituitary hormones showed that action potentials cause Ca2+ influx, triggering exocytosis, visualised with electron microscopy (Douglas and Poisner, 1964; Morris and Pow, 1988). Importantly, the rate of peptide release disproportionately increases as action potential frequency increases, indicating a mechanism at the axon terminals for facilitating secretion of a pulse of peptide (Leng et al., 2001; Shaw et al., 1984). Furthermore, in contrast, pre-terminal inhibition by endogenous opioid (revealed by the antagonist naloxone), and exogenous opiates, was shown, but this is a site of plasticity as the opioid inhibition of oxytocin secretion is lifted by the end of pregnancy (Douglas et al., 1993). Together, these studies on the posterior pituitary provided a paradigm for secretion by other peptidergic neurons.
Monitoring the electrophysiological activity of populations of neuroendocrine neurons is a challenge, but Ernst Knobil’s group found increased multi-unit activity in the mediobasal hypothalamus ahead of a pre-ovulatory LH surge in rhesus monkeys and deduced this to indicate excitation of GnRH neurons (Wilson et al., 1984). Such periodic activity of GnRH neurons will provide the pulsatile secretion of GnRH essential for preventing down-regulation of GnRH receptors on gonadotrophs (Wildt et al., 1981). Related studies showed that sex steroid positive feedback, essential for pre-ovulatory gonadotropin surges, only needed an intact mediobasal hypothalamus in the monkey (Krey et al., 1975), contrasting with rodent studies in which more rostral regions had been established to be essential (Rodgers and Schwartz, 1972).
Real time ex vivo
The electrophysiological properties of neuroendocrine neurons have been investigated with intracellular or patch-clamp electrode recording in hypothalamic brain slices or organotypic cultures (Armstrong et al., 2010; Hatton, 1983; Yamashita et al., 1983). Magno- and parvo-cellular neurosecretory neurons have been shown to have essentially similar properties to other neurons, with regard to membrane conductances, synapses and capacity for retrograde modulation (e.g. via nitric oxide, endocannabinoids; Figure 2) of synaptic input and auto-regulation. For vasopressin neurons, a property that explains direct osmosensitivity of these neurons is expression of a TRPV1-like non-selective stretch-inactivated cation channel, translated from a modified transcript of the trpv1 gene (Sharif Naeini et al., 2006).
Definition of these properties, and incorporating data about in vivo electrophysiological activity including from neuropharmacological studies, has enabled dynamic computer-based modelling of the electrical activity of both magnocellular oxytocin and vasopressin neurons that reproduces in vivo activity, including phasic firing of vasopressin neurons and the co-ordinated burst firing of oxytocin neurons that requires positive auto-feedback by dendritically released oxytocin as seen during suckling (Rossoni et al., 2008). In general, such models test validity of the assumptions about roles of each of many variables incorporated in the model, and inform and guide further experiments. For instance, termination of a vasopressin neuron phasic burst is best explained by including the experimental finding of auto-inhibition by dendritically released dynorphin (Brown et al., 2006), and burst firing of oxytocin neurons during suckling is expected to require inhibitory action of an as yet unidentified peptide (Rossoni et al., 2008).
Historical time/ Activity in time past
A highly productive (but post mortem) approach is to use changes in gene expression (actually in levels of hnRNA or mRNA by quantitative ISH) or expression of specific proteins (by ICC) in a sample of individual neurons of interest soon after a relevant experience to test hypotheses about neuronal function, or indeed about regulation of target genes. This approach is based on several findings: (1) expression level of Fos, a transcription factor, and of its gene, c-fos, is generally low but is rapidly stimulated when a neuron is excited, maximally with a delay of about 30 min (Figure 2); Fos is then readily visualised in the nucleus with ICC; combined with phenotype identification by ICC, the number of such neurons that have been excited can be counted; in addition, combination with retrograde labelling from a putative source of input further characterises the phenotype (Meddle et al., 2000); (2) some neuropeptide genes are rapidly activated by excitation of the neuron (e.g. parvocellular CRH neurons, but not magnocellular neurons – reflecting huge peptide stores in the latter); hence, in these parvocellular neurons, quantitative ISH for specific mRNAs has been used widely to compare responses under different conditions to address questions about the type of input the neurons respond to (Lightman and Young, 1988).
Molecular genetics
Recognition and subsequent functional and molecular genetic analysis of spontaneous mutations in laboratory rodents and humans have profoundly altered understanding of the functioning of specific neuroendocrine systems and have led to revolutionary new paradigms, uncovering new neuroendocrine systems. Manipulating genotype, especially in mice, to link expression of a particular gene (e.g. a neuropeptide) to a fluorescent marker (e.g. green fluorescent protein, GFP) has enabled real-time identification of phenotype in electrophysiological studies; identification by RT-PCR of the phenotype of single neurons has also been used as a post hoc method.
Brattleboro rat
This strain was derived from a litter of Long-Evans hooded rats observed to drink excessively and to excrete copious dilute urine (diabetes insipidus), treatable with vasopressin injections. A recessive gene defect was identified, a single base deletion in the vasopressin gene, that does not affect vasopressin but alters the neurophysin protein, and the mRNA is not efficiently translated (Schmale et al., 1984). This natural gene ‘knockout’ strain has been used to show, for example, roles of vasopressin in the brain in stress responses
Hormone rhythms
Hypothalamic-pituitary-adrenal (HPA) axis and suprachiasmatic nucleus
By the 1960s, classical lesioning studies had revealed the distinct visual pathways critical for the well-described influences of circadian and seasonal rhythms on behaviours and neuroendocrine functions, especially of the pineal gland (Moore et al., 1967). Retinal input to the suprachiasmatic nucleus (SCN) had also been described, and for the distinct diurnal rhythm of the HPA axis, electrolytic SCN lesions were shown to suppress the diurnal peak in adrenal glucocorticoid content in rats (Moore & Eichler, 1972); glucocorticoid secretion peaks just before dark in nocturnal animals, and in the morning in humans, that is, there is a 180° phase difference in HPA activity between nocturnal and diurnal species. Importantly, this is not a result of phase difference in SCN neuron activity, as this is consistently greater in the light period (Schwartz and Gainer, 1977), but to different connections of SCN outputs. Thus, SCN neurons, especially vasopressin neurons in the shell of the SCN, project to the hypothalamus and indirectly to pPVN CRH neurons, and are active in the light phase, providing a basis for circadian rhythm in HPA axis function: but the effect of SCN vasopressin neurons is to inhibit CRH neurons indirectly via GABAergic neurons in the nocturnal species, and in diurnal species to excite CRH neurons indirectly, via glutamatergic neurons (Kalsbeek et al., 2015).
Ultradian HPA axis rhythms
Fine temporal resolution of changes in the circulating concentrations of ACTH and glucocorticoids revealed ultradian rhythmicity in their secretion, with peaks at ca. 60-min intervals; each peak in glucocorticoid concentration is preceded by a peak in ACTH secretion, which drives the former (Lightman and Conway-Campbell, 2010; Windle et al., 1998). The diurnal rhythm in ACTH and glucocorticoid secretion is superimposed on the ultradian pattern, such that the amplitude of the ultradian peaks is greatest in the nocturnal rat around the end of the light phase, as a result of increased CRH, and hence ACTH, secretion at this time (Lightman et al., 2008). However, the ultradian pattern does not involve the SCN or changes in CRH secretion, although basal CRH drive is needed (Lightman et al., 2008; Waite et al., 2012). Instead, the ultradian rhythms arise from delays in the feed-forward action of ACTH on glucocorticoid synthesis in the adrenal and the negative feedback action of glucocorticoid on the anterior pituitary corticotrophs (Rankin et al., 2012). Mathematical and computational modelling show these properties result in oscillations in ACTH and glucocorticoid secretion (Rankin et al., 2012), as seen in vivo (Windle et al., 1998).
Significance of glucocorticoid pulses
The functional significance of pituitary-adrenocortical ultradian rhythmicity is two-fold: firstly, sensitivity to stressful stimulation is greater at the start of an ultradian peak, and reduced after a peak (Windle et al., 1998), providing a mechanism to filter minor stimuli (Rankin et al., 2012); secondly, ultradian rhythmicity in glucocorticoid levels is measurable in the brain as well as peripherally (Droste et al., 2008), and this impacts on the molecular dynamics of actions of glucocorticoid on gene expression, mediated by glucocorticoid receptors (GR) rather than mineralocorticoid receptors (MR) (Stavreva et al., 2009). In the anterior pituitary, ultradian glucocorticoid peaks are followed by transient waves of GR briefly binding to regulatory elements of specific genes, with transient increases in gene expression, and production of quanta of newly synthesised mRNA; some genes (e.g. POMC in the anterior pituitary) are inhibited (George et al., 2017a; Stavreva et al., 2009). In the hippocampus, GR activation/deactivation follows the ultradian glucocorticoid cycle (Lightman et al., 2008), but in the prefrontal cortex an ultradian glucocorticoid pulse has longer-lasting effects on gene expression than in the anterior pituitary (George et al., 2017a). Evidently, for some genes, in some brain regions, but not for others, the effects of ultradian peaks of glucocorticoid on gene expression are smoothed. There are implications here for other steroid feedback mechanisms: for instance, those involved in regulation of ovarian cycles.
Paradigm shift: electrical and molecular clockwork
Electrical rhythm
Amazingly, studies on the SCN hub showed that after microsurgical isolation of the SCN in situ and in hypothalamic slices in vitro SCN neurons maintain circadian rhythms in their electrophysiological activity (Green and Gillette, 1982; Inouye and Kawamura, 1979; Weaver, 1998). Furthermore, SCN transplants substantially restore rhythm if transplanted into animals with SCN lesions (Lehman et al., 1987): hence the SCN was deduced to be the primary circadian pacemaker and to contain an intrinsic clock mechanism. Moreover, the SCN neurons, together with the pineal, respectively, synchronise the clocks now known to be widely distributed in cells in the brain and body (Buijs et al., 2016), profoundly influencing metabolism, and seasonal rhythms related to reproduction.
Individual SCN neurons have independent electrophysiological rhythms, with more frequent action potentials during the day, but need the company of other SCN neurons to optimise rhythmicity. Greater activity during the day is related to reduced membrane potential, with hyperpolarisation at night reducing activity (Figure 4(a) and (b)). The SCN contains several neuronal phenotypes that express neuropeptides (notably, vasoactive intestinal peptide (VIP) and gastrin releasing peptide (GRP) in neurons in the core of the SCN, which receives retinal input; vasopressin neurons in the shell, which is more strongly rhythmic and projects into the brain), and GABA and clock genes, showing circadian variation in expression and necessary for clock function (Evans, 2016). Such clocks consist of now well-defined molecular autoregulatory transcriptional/post-translational feedback loops (TTFL; Figure 4c).

Suprachiasmatic nucleus (SCN) circadian electrical and molecular clocks. (a), (b) Circadian rhythm in electrical activity. SCN neurons are more active electrically during the circadian day than the night. (a) The resting membrane potential (RMP) is more depolarised during the day (UP state), slowly increasing and then decreasing during the day, and vice versa during the night. During the day, the RMP changes reflect reduced activity of L-type Ca2+ channels, and Ca2+-activated K+ channels; at night, K+ channels open and the neurons are hyperpolarised (DOWN state). (b) UP state: Some neurons discharge action potentials (AP) at 4–6 spikes/s (b1), others with a high RMP show waves of depolarisation (b2), glutamate from light-activated retinal ganglion cells provides excitatory drive, and activates per1 (see Figure 4c); DOWN state: action potentials are produced at a lower rate, 0.1–3 spikes/s (b3). (c) Diagram of the core molecular components of the molecular clock. The core clock comprises transcriptional/post-translational feedback loops (TTFL) regulating expression of clock genes and proteins that take 24 h to complete a cycle. Clock and Bmal1 proteins (1), transcription factors, peak at the start of the circadian day, and act on E-boxes (2) to enhance expression of target genes, including the period (per1/ 2) and cryptochrome (cry) genes (3), translation of their mRNAs (4) leads to increased cytoplasmic levels of Per and Cry proteins by the end of the day (5), which enter the nucleus and auto-repress per and cry gene expression by interfering with Clock and Bmal1 actions at E-boxes (6; pink crosses X). Consequently, Per and Cry protein production stops, and remaining Per and Cry are degraded. This allows Clock and Bmal1 proteins to restart per and cry gene expression and a new cycle. The products of other genes contribute to feedback loops involving Clock and Bmal1, and oscillation in Bmal1 expression.
Clock genes
Studies of the molecular genetic basis of circadian rhythms in Drosophila uncovered a group of genes that interact as a molecular clockwork to generate cellular and behavioural rhythms (Reddy et al., 1984; Sehgal et al., 1995), and homologous genes were found in vertebrates (Shin et al., 1985). A molecular genetic basis of circadian rhythms in mammals was indicated by discovering a mutant Syrian hamster that showed an abnormally short circadian rhythm in constant dark (tau mutant; Ralph and Menaker, 1988), which was inherited and transplantable via SCN implants into normal hamsters with SCN lesions (Ralph et al., 1990). This was followed by finding in mice exposed to mutagens a phenotype with a longer than normal free-running period. The Clock gene responsible was cloned and the protein Clock identified as a transcription factor. Its function was subsequently explained by heterodimer formation with Bmal1 to drive per expression with Per in turn auto-inhibiting per (Gekakis et al., 1998); the cryptochrome (cry) gene was then shown to be important and regulated like per. The genes targeted by the proteins act via E-box enhancer elements: here Clock/Bmal1 activate and Per/Cry repress gene expression.
Molecular clockwork
These interactions slowly develop each day: in the morning Clock/Bmal1 activates per and cry, so their transcripts accumulate, followed by their proteins, which oppose Clock/Bmal1 actions and Per/Cry production falls, and these proteins are degraded. A new cycle of oscillating feedback can now start (Hastings et al., 2014; Figure 4c). These rhythms are visualised as waves of activity across the SCN in vitro with bioluminescent and fluorescent reporters (Hastings et al., 2014). Multiple gene products are involved in regulating speed of the clock, other genes are involved in augmenting the cycle, and microRNAs (miR-219 and miR-132) are involved as clock regulators (Cheng et al., 2007). Light, acting via glutamate input from the retina, resets the clock, via SCN neurons producing VIP, which acts in a paracrine fashion and synaptically on other SCN neurons, via VPAC2 receptors, with subordinate roles for vasopressin and GRP, and a second messenger pathway to the per gene (Hastings et al., 2014). SCN neurons also receive synaptic input from within the brain, including input reflecting arousal state (signalled by the neuropeptide orexin). So far, uncertain is how the TTFL is linked reciprocally to electrical activity within SCN effector neurons, although an electrical membrane clock seems plausible, while interactions among SCN neurons are well-described (Belle, 2015; Evans, 2016).
Reproductive neuroendocrine rhythm
GnRH neurons
Identified immunocytochemically, GnRH neurons were shown to have a distinctive developmental origin – born in the olfactory placode, they migrate into the preoptic area, guided by multiple chemical signals (Schwanzel-Fukuda and Pfaff, 1989; Wierman et al., 2011; Wray et al., 1989). Here they are quite scattered; each is bipolar, with a rostral and a caudal dendrite, the latter up to 1 mm long and projecting to the median eminence where its axonal-like terminals abut the primary capillaries of the portal system. These dendrites have synaptic boutons all along their length, and the spike initiation site on the caudal process is unusually far (150 µm) from the cell body: the hybrid caudal process has thus been described as a dendron (Herde et al., 2013). Evidently, this feature of GnRH neurons gives huge potential for preterminal modulation of GnRH release, in which the entwinement of dendrons is likely important. Understanding how these neurons function to release ca. hourly GnRH pulses, and consequently trigger hourly LH pulses is not yet clear; individually they discharge brief intermittent bursts of action potentials. The co-ordinated burst-firing oxytocin neuron model may give clues, though GnRH neurons are older in evolution (preceding lactation) than oxytocin neurons (notwithstanding oxytocin neuron precursors in egg-layers).
Kisspeptin: a dramatic breakthrough in reproductive neuroendocrinology
Kisspeptin neurons
A now predominant idea that non-GnRH neurons are involved in GnRH rhythms followed the discovery in 2003 that hypogonadotropic hypogonadism (i.e. no puberty, infertility of central origin) in men and women is associated with a loss of function mutation in GPR54 (now called KISS1R; de Roux et al., 2003; Seminara et al., 2003). The ligand for this receptor is a 54 a.a. RF (arginine-phenylalanine) amide related peptide (RFRP), metastatin/kisspeptin, which like its gene KISS1 was already known in cancer biology. KISS1R is expressed by GnRH neurons, kisspeptin strongly stimulates GnRH neuron electrical activity and GnRH secretion, and is produced by kisspeptin neurons in the arcuate nucleus, and in the anteroventral periventricular nucleus (AVPV) in the preoptic area: close to the GnRH neurons and their dendrons (Figure 5).
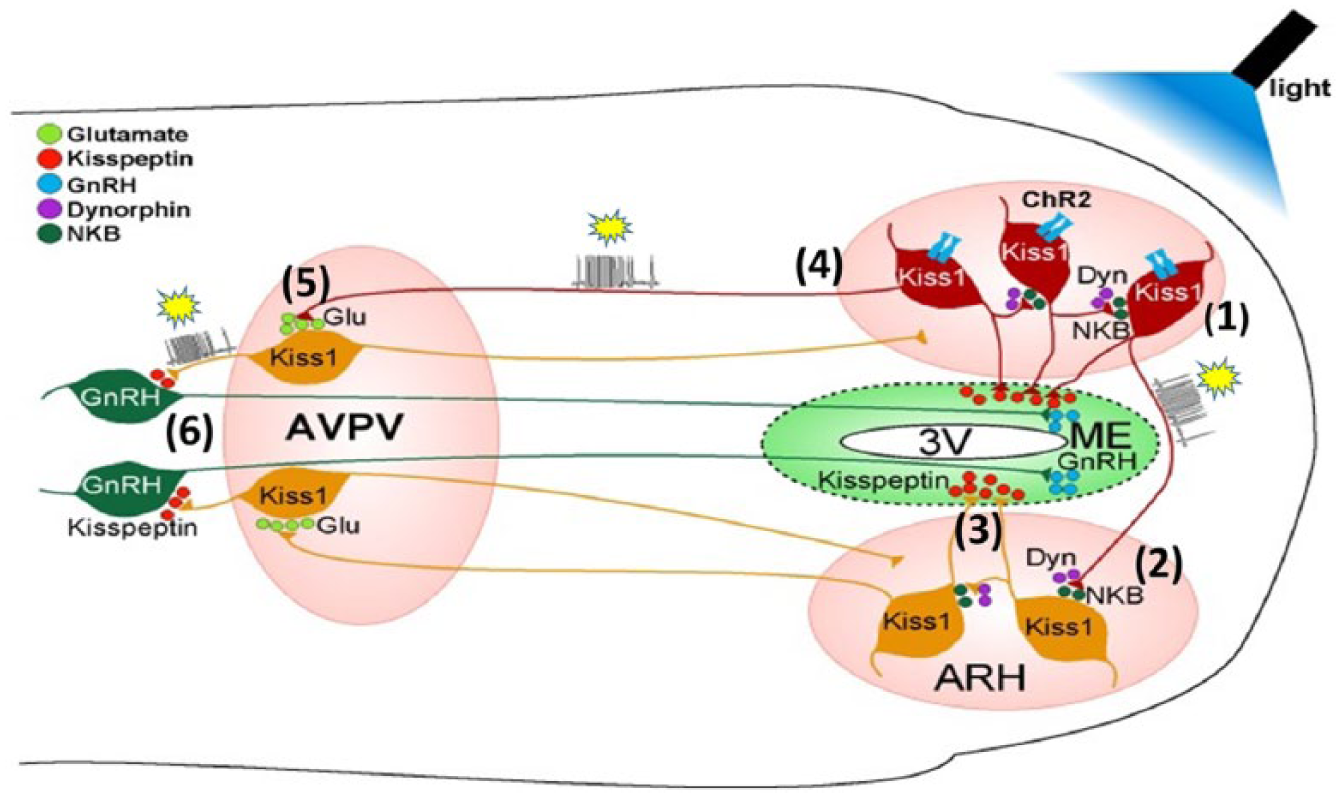
Micronet linking kisspeptin and gonadotropin releasing hormone (GnRH) neurons. Kisspeptin (Kiss 1) networks in the basal hypothalamus (arcuate nucleus, ARH) and anteroventral periventricular nucleus (AVPV), and synaptic connections with GnRH neurons in the preoptic area were studied with optogenetics, whole-cell electrophysiology, molecular pharmacology and single cell RT-PCR. Para-horizontal or coronal brain slices were used, from female mice, transgenically modified to express channelrhodopsin 2 (dark brown cells, ChR2; activated by blue light to cause depolarisation) and fluorescent protein markers. Arcuate nucleus kisspeptin (KNDY) neurons co-express dynorphin (Dyn; κ-opioid peptide) and neurokinin B (NKB). (1) High frequency photostimulation of Kiss 1 neurons (yellow stars: action potential bursts) excites adjacent and (2) contralateral Kiss 1 neurons, involving modulation by Dyn (inhibitory) and NKB (excitatory); hence KNDY neurons can be excited in a coordinated manner. (3) Kisspeptin released in the median eminence (ME) may promote GnRH release by acting on the long processes (dendrons) of GnRH neurons. (4) Arcuate KNDY neurons project to the AVPV Kiss 1 neurons; (5) when the KNDY neurons are stimulated in turn they excite Kiss 1 neurons via glutamate (Glu) release. (6) Trans-synaptic excitation of Kiss1 neurons projecting to GnRH neurons releases kisspeptin to excite GnRH neurons projecting to the median eminence (ME). GnRH released into the hypothalamic-hypophysial portal vessels will stimulate LH and FSH release by the anterior pituitary. The network described may operate as the ‘GnRH pulse generator’ and might explain pre-ovulatory surges of GnRH and LH.
KNDy neurons
Arcuate kisspeptin neurons also express neurokinin B (NKB; loss of function mutations of genes for this peptide and its receptor in humans have effects like GPR54 loss- no puberty, infertility) and dynorphin (hence K-N-Dy, KNDy neurons). The discovery of kiss-1 neurons has led to a model of local interactions with GnRH neurons which could generate the ca. hourly pulses of GnRH that cause the LH pulses known for more than 40 years. Unlike GnRH neurons, kiss-1 neurons express estrogen receptors, so they could mediate differential negative (arcuate KNDy neurons) and positive (preoptic kiss-1 neurons) feedback actions of estrogen on the GnRH neurons that also have been known for over 40 years to be key in triggering ovulatory LH secretion; except that primates, including women, evidently produce an ovulatory LH surge independently of a GnRH surge, although continual GnRH secretion is a requirement (Belchetz et al., 1978; Plant, 2015). Criteria have been satisfied for a key role of kisspeptin neurons as GnRH pulse generators: in vivo, in mice, photo-stimulation of arcuate kisspeptin neurons expressing channelrhodopsin induced pulses of LH secretion (Han et al., 2015); in vitro, in mouse brain slices, using multiple techniques mentioned above, excited KNDy neurons show synchronised activity and stimulate GnRH neurons (Qiu et al., 2016).
It remains to be seen whether these findings extend to the pre-ovulatory GnRH and LH surge, and explain positive and negative 17β-estradiol and progesterone feedback, and how the kiss-GnRH circuitry is activated for puberty.
Appetite control by peptides
Seeking a fat signal to the brain
Early brain lesioning and stimulation studies indicated that the ventromedial hypothalamus (VMH) and the lateral hypothalamus (LatH) organise satiety and hunger, respectively. Hervey (1959; Parameswaran et al., 1977) showed that in parabiotic pairs of rats, a VMH lesion or LatH electrical stimulation in one rat induced in that rat increased food intake and obesity, but had the opposite effect in the partner; Hervey postulated a humoral factor from adipose tissue acting centrally would feedback to control appetite, crossing into the parabiotic partner.
Breakthrough: ob/ob mouse and leptin
Subsequent genetic analysis of the ob/ob genetically obese mouse (a strain known since 1949) identified the defective gene, expression of the normal gene in adipose tissue and its protein product, a hormone, injections of which were shown to reverse the hyperphagic and obese phenotype in the ob/ob mouse (Halaas et al., 1995). This protein hormone, leptin (from the Greek, leptos, thin), is a cytokine, or adipokine as it is produced by adipocytes. Subsequently, the JAK2/STAT3-linked leptin receptor (ObRb) was characterised (via analysis of obese db/db mice, discovered in 1966, which have a defective leptin receptor gene, so are leptin insensitive), and shown to be expressed in the hypothalamus (Schwartz et al., 1996). However, treatment of obese people with leptin was unsuccessful (except for rare patients with a leptin gene mutation), indicating leptin resistance in obesity.
Paradigm shift
These findings necessitated building endocrine activity of adipose tissue into a neuroendocrine model of appetite and metabolic regulation. Producing this model has involved applying the full weight of a range of contemporary technology and collaborative effort. Leptin enters the brain via the choroid plexus and basal hypothalamus, by-passing the blood-brain barrier, and acts on leptin receptors (ObRb) expressed on neurons in the hypothalamic network and beyond. This still developing model describes complex neural networks with hubs and nodes containing several types of neuropeptide neuron in the arcuate nucleus, VMH, LatH, parvocellular (p) PVN and nucleus tractus solitarii (NTS), as proposed by Schwartz et al. (2000) (Figure 6). By virtue of the connections of the network neurons and actions of neuropeptide produced these are considered as orexigenic or anorectic, so the principle has emerged that appetite (measured as feeding) is suppressed by inhibition of the former and/or excitation of the latter, and appetite is increased by respective stimulation or inhibition of orexigenic and anorectic neurons. It is clear that leptin is but one circulating peptide that acts on the arcuate nucleus hub: insulin (signalling adequate energy) acts in the same direction as leptin; by contrast, circulating ghrelin (signalling an empty stomach) activates orexigenic neurons.

First order to second order metabolic signalling from arcuate nucleus (ARC). Diagram of transverse section through one half of hypothalamus. Leptin from the circulation signals adipose store, inhibiting first order (1°) NPY/AgRP (yellow) neurons (which increase food intake) and stimulating POMC/CART (purple) neurons (which decrease food intake) in the arcuate nucleus. AgRP: agouti-related peptide; CART: cocaine and amphetamine regulated transcript; NPY: neuropeptide Y; POMC: pro-opio-melanocortin; 3V: third ventricle. NPY and POMC arcuate neurons project into the hypothalamus including to the paraventricular nucleus (PVN; which also has reciprocal brainstem connections, not shown), perifornical area (PFA), lateral hypothalamic area (LHA; long established as a ‘hunger centre’) and ventromedial hypothalamus (VMH, not shown, a ‘satiety centre’). Candidate second order (2°) targets include, in the PVN, thyroid releasing hormone (TRH, light green), corticotropin releasing hormone (CRH, blue), oxytocin (OXY, pink) neurons – which have anorexic actions; and in the PFA, orexin (orange), and in the LHA, melanocyte concentrating hormone (MCH, dark green) neurons – which increase feeding.
Arcuate nucleus – further peptide complexity
The arcuate nucleus orexigenic and anorectic neurons each produce one or more peptides and project to other nodes or hubs (notably the pPVN and LatH), the NTS and to each other: in the arcuate nucleus, a GABA connection from orexigenic (neuropeptide Y/agouti-related peptide, NPY/AgRP) to anorectic (POMC/cocaine and amphetamine regulated transcript [CART]) neurons tonically favours appetite. Conversely, leptin activates POMC/CART neurons and inhibits NPY/AgRP neurons, which removes the GABA tone. NPY acts rapidly via Y1/Y5 receptors; actions of AgRP are slower, but prolonged; α-MSH is the active peptide released by arcuate POMC neurons, acting on melanocortin-4 (MC4) receptors (e.g. in the PVN and LatH), and AgRP opposes α-MSH actions directly, acting as an MC4R reverse agonist. Other candidate anorectic neuro-peptides have weighed in, for example, PrRP (from the NTS). Oxytocin has central anorectic actions, evidently via the VMN, and in vivo, α-MSH inhibits electrical activity of magnocellular oxytocin neurons, but stimulates dendritic release of oxytocin, hence indicating a possible anorectic mechanism for α-MSH (Sabatier et al., 2003). These oxytocin neurons are activated by a noradrenergic input from the A2 noradrenergic neurons in the NTS, which is stimulated by circulating CCK and secretin (indicators of a recent feed), acting via vagal afferents (Velmurugan et al., 2010); hence, centrally released oxytocin may contribute to satiety after a meal, as indicated by studies on oxytocin knockout mice.
Electrophysiological and functional pathway tracing studies
Evaluating and eventually dynamically modelling interactions among the closely associated neurons with different phenotypes within these clusters is an ongoing 21st-century challenge in neuroendocrinology. This challenge is being addressed by using contemporary techniques to map these networks and functions, founded on distribution of specific receptors: these include retrograde and anterograde tracing with micro-/nanoparticles or viruses (including trans-synaptic viruses) (Oldfield et al., 2016), manipulation in mice of specific peptidergic neurons by selective ablation, or inhibition and excitation by designer receptors exclusively activated by designer drugs (DREADDs), and optogenetic activation, coupled to electrophysiological recording and behavioural studies. This combination of techniques has been used to show in mice the wide penetration of arcuate nucleus AgRP projections (Sternson and Atasoy, 2014), to reveal a strong stimulatory connection from PVN TRH and pituitary adenylate cyclase-activating polypeptide (PACAP) neurons to arcuate nucleus AgRP neurons (Krashes et al., 2014; this is intriguing as TRH and PACAP have contrasting central anorectic actions), and that CCK-producing neurons in the NTS, which are activated by feeding, project to MC4R positive (α-MSH sensitive) neurons in the pPVN and excite some of these appetite-suppressing neurons (D’Agostino et al., 2016).
HPT axis
Thyroid hormone is a major regulator of metabolic rate and the TRH neurons in the pPVN are an intrinsic component of the network regulating appetite and metabolism. TRH neurons receive input from leptin sensitive arcuate nucleus components of the network such that they are stimulated by leptin acting via α-MSH/CART projections and inhibited via NPY/AgRP neurons (Figure 6). Hence, activity of the HPT axis is quite responsive to nutritional state (i.e. inhibited by fasting), interacting with negative feedback control of Trh gene expression by thyroid hormone.
Hunger
Hunger and satiety are powerful feelings, and the network regulating appetite and metabolism is connected with reward circuitry, via indirect or direct leptin actions on mesolimbic dopamine neurons, and involving ghrelin actions, and neurotensin neurons (Farooqi et al., 2007; Opland et al., 2013; Perello and Dickson, 2015).
Perspective
As more details of the appetite and metabolism-regulating networks in the brain’s connectome emerge, dynamical computer-based modelling of the power of each of the components involved should reveal targets for new drugs to alter settings within this neuroendocrine system in the management of obesity and anorexia.
Programming
Central leptin resistance can result from early life obesogenic conditions (pre- and post-natal) and persist into later life as a programming effect on development of the regulatory neuropeptide neurons leading to adult obesity (Croizier and Bouret, 2016).
Neuroendocrine system plasticity
Early life programming
There are several examples of early life (pre- or post-natal) programming of the activity of neuroendocrine systems as a result of exposure to natural biological transformative signals, or to a stressor, or particularly favourable or unfavourable nurturing, or extreme nutritional state. Programming resets or switches genetically determined activity in neuronal networks. Examples considered here are sexual differentiation of the brain, pre-natal and early life programming of neuroendocrine stress responses.
Reproductive behaviour and gonadal control: perinatal masculinisation and defeminisation
The potential for central neuroendocrine mechanisms to organise the cyclical secretion of gonadotropins for ovulation and to perform respective sexual behaviour is present in both genetically male and female developing brains. The organisational/activational concept of sexual differentiation of the brain arose from series of experiments begun in the 1950s showing that the central mechanisms organising sex-specific behaviours in males undergo defeminisation and masculinisation as a result of fetal testosterone secretion and action late in gestation and just after birth (McCarthy, 2010; Phoenix et al., 1959) (Figure 7). These principles of sexual differentiation of the brain with regard to gender and sexual orientation apply to humans, as indicated by detailed brain morphological studies in the context of clinical history (Bao and Swaab, 2011).

Sexual differentiation of the brain: epigenetic programming of organisation by 17β-estradiol. Above: During a critical period in late gestation and early post-partum, testosterone from the perinatal testes is converted into 17β-estradiol (E2) by aromatase expressed in genetic males in the nascent sexually differentiated hubs in the neuroendocrine brain. E2 binds to intracellular estrogen receptor α (ERα), a transcription factor, which enters neuronal nuclei, binds to DNA at specific sites and engages a transcription complex, which includes histone acetyl transferases (HATs), enabling access to DNA, and CREB-binding protein (CBP). DNA methyl transferase-3 (DNMT3) methylates cytosine in DNA at selected CpG islands, repressing gene expression, and importantly activated ERα evidently interferes with DNMT3 action in female neonates, reducing DNA methylation, which is essential for masculinisation and defeminisation of the brain in males. Below: Typical male sexual behaviour (black rat) is illustrated as the outcome in a genetically female neonate given E2 treatment and then when adult treated with testosterone to activate the masculinised and defeminised brain; testosterone does not have this effect in normal females.
Key role for 17β-estradiol
Testosterone acts after conversion to 17β-estradiol by aromatase expressed during the critical period in development in the brain target regions, in particular the medial preoptic area (mPOA), important for male sex behaviour, and the VMN, important for female sex behaviour. The effects of perinatal estrogen (i.e. in males) include, via estrogen receptors, the prevention of apoptosis in the sexually dimorphic nucleus (SDN, in the mPOA), increasing number of spines on mPOA neuron dendrites, loss of cells in the AVPV, in particular GABAergic and dopaminergic neurons (consistent with lack of support in males for the gonadotropin surge pattern stimulated by positive estrogen feedback for ovulation as in females), and defeminisation of the VMN, ablating female reproductive behaviour. Multiple mechanisms mediate these effects of estrogen, for example, prostaglandin E2 mediates estrogen action on mPOA dendrites, indirectly via glutamate and α-amino-3-hydroxy-5-methyl-4-isoxazolepropionic acid (AMPA) receptors (McCarthy, 2010).
Epigenetics
It has been anticipated that the programming effects of 17β-estradiol derived from testosterone in the perinatal male brain would include epigenetic changes. There is evidence for epigenetic marks, DNA methylation at CpG sites (generally indicating gene repression) by DNA methyl transferases (DNMT). Female neonates have higher DNMT activity in the POA than males, with periods of expression in development essential for feminisation. Methylation can be dynamic with multiple mechanisms for demethylation, which is essential for masculinisation: the critical period for masculinisation is closed by DNA methylation, but can be kept open by treatment with a DNMT inhibitor. Histone de-acetylation (increasing gene expression) is involved in masculinisation, affecting multiple genes, so inhibition of de-acetylation impairs masculinisation of reproductive behaviour, but it is not clear which genes are critical (McCarthy and Nugent, 2013).
Maternal brain adaptations: neurosteroids and stress resilience in pregnancy
Steroid milieu
Pregnancy involves massive changes in physiology and exposure of the brain to increased circulating concentrations of several hormones, from the ovaries, fetus and placenta, which elicit multiple changes in neuroendocrine mechanisms essential for the pregnancy and subsequent motherhood (Brunton and Russell, 2015). Progesterone is produced in large amounts in pregnancy and importantly, this can be converted by sequential actions of 5α-reductase and 3α-hydroxysteroid dehydrogenase (3α-HSD) in the brain into allopregnanolone, a neurosteroid allosteric enhancer at GABAA receptors.
Neuroendocrine hyporesponsiveness to stress
Among the neuroendocrine changes uncovered around the start of the 21st century is attenuation of HPA axis responses to stress, which is considered to give some protection to fetuses from adverse programming by glucocorticoid (Johnstone et al., 2000). This attenuation is seen in responses to physical and emotional stressors (e.g. interleukin-1β, IL-1β injection, as an infection mimic; aggression from a con-specific – social stress). Without pregnancy, IL-1β activates a noradrenergic pathway from the NTS to the CRH neurons in the pPVN, but in late pregnancy while IL-1β activates Fos in the NTS, it does not increase noradrenaline release or CRH mRNA expression in the PVN, and ACTH and corticosterone secretion are barely increased.
Allopregnanolone-opioid mechanism
Activation in pregnant rats of an endogenous opioid mechanism (PENK-A mRNA expression in the NTS is increased) is involved in reducing HPA axis stress responses as the opioid receptor antagonist naloxone, given before IL-1β, restores noradrenaline release and HPA axis responses. In turn, the opioid mechanism is activated by the high levels of allopregnanolone, locally produced in the NTS (Brunton et al., 2009) (Figure 8). HPA axis responses to stress return after parturition, but are reduced by the suckling stimulus in lactation. Whether the changes in NTS noradrenergic neurons in pregnancy involve temporary allopregnanolone-induced epigenetic changes is not known. The same allopregnanolone-induced opioid mechanism is also responsible for attenuated magnocellular oxytocin neuron responses to IL-1β in late pregnancy, which is seen as a mechanism preventing premature birth.

Neurosteroid and opioid suppression of pPVN CRH neuron activity in pregnancy. In late pregnancy, hyporesponsiveness of the hypothalamic-pituitary-adrenocortical (HPA) axis to stressors is explained by reduced activation of corticotropin releasing hormone (CRH) neurons in the medial parvocellular paraventricular nucleus (mpPVN). Without pregnancy, intravenous interleukin-1β (IL-1β) strongly activates the HPA axis, acting via A2 noradrenergic neurons in the nucleus tractus solitarii (NTS) that project to the pPVN, releasing noradrenaline (NA) to excite CRH neurons via α1 receptors. In late pregnancy, NA release in the PVN, CRH neuron responses (marked by increased CRH mRNA expression) to IL-1β, and consequently ACTH and corticosteroid responses, are all suppressed. There are two connected mechanisms: (1) Naloxone given before IL-1β reverses the changes, indicating activation of an endogenous opioid (EOP) mechanism in pregnancy; pro-enkephalin-A (PENK-A) and μ-opioid receptor (MOR) mRNA expression in the NTS is increased in pregnancy, providing a mechanism to explain presynaptically inhibited NA release in the PVN. (2) Allopregnanolone (AP) levels in the brain increase in pregnancy, as a result of conversion from high progesterone levels by 5α-reductase (5αR) and 3α-hydroxysteroid dehydrogenase (3αHSD); 5αR and 3αHSD mRNA expression and activity in the NTS is increased, indicating AP synthesis here. AP is an allosteric modifier of GABAA receptors (prolonging open time for Cl- entry), a possible action in pregnancy on CRH neurons, while blocking 5αR with finasteride reverses the EOP inhibition of the HPA axis, indicating that AP induces this EOP inhibition in pregnancy.
Adversely programmed offspring phenotype
Emotional stressors activate rostral limbic brain circuitry controlling the HPA axis, and HPA responses in pregnancy to social stress are attenuated, but not fully suppressed, giving an opportunity for programming by glucocorticoid transferred across the placenta. Adult offspring of mothers exposed to social stress in pregnancy show exaggerated HPA axis responses to IL-1β and, in males, anxious behaviour (Brunton, 2013) (Figure 9).
Allopregnanolone deficit
Expression of 5α-reductase mRNA in the NTS is reduced in such prenatally stressed offspring, and repairing this using adenovirus-mediated gene transfer, or giving allopregnanolone, normalises HPA axis responses (Brunton et al., 2015). Prenatally stressed offspring show underlying changes in expression of multiple mRNAs in brain regions organising HPA axis stress responses and anxiety, with similarities to models of post-natal stress (Maccari et al., 2014) (Figure 9). The involvement of epigenetic mechanisms in the gene expression changes in the offspring brain in the prenatal social stress model is yet to be determined, though some other studies have found methylation of stress-related genes (Mueller and Bale, 2008).

Prenatal social stress changes mRNA phenotype of stress processing networks in the offspring brain. Pregnant rats were exposed to social defeat by a lactating rat for 10 min each day in the last week of pregnancy. Male offspring were compared with control males as adults for measurement by in situ hybridisation of basal expression of mRNAs in coronal brain sections (Basal, left), and acute changes after i.v. IL-1β (Stress, right). mRNAs assessed were in neurons driving the HPA axis (parvocellular paraventricular nucleus (pPVN), arginine vasopressin (AVP) and corticotropin releasing hormone (CRH) under basal (1) and stress conditions (2). Further measurements were under basal conditions on mRNAs for glucocorticoid (GR) and mineralocorticoid (MR) receptors in hippocampus (3), as indicators of dynamic and basal feedback, respectively; GR, CRH and CRH-R1 (receptor 1) in central nucleus of amygdala (4; CeA), as indicators of activity of anxiogenic genes; CRH-R1 and CRH-R2 in three other amygdala regions (5; basolateral, BLA, basomedial, BMA, Medial, MeA) as indicators of relative expression of anxiogenic (CRH-R1) and anxiolytic (CRH-R2) genes; overall, the ratio of CRH-R1 mRNA: CRH-R2 mRNA was increased in the prenatally stressed rats. ↑, Greater expression; ↓, lower expression and ↔, no difference in mRNA expression vs. controls. The data are consistent with observed increased HPA axis secretory responses to i.v. IL-1β, anxious behaviour, and cognitive impairment are similar to other studies of experimental prenatal stress on rats and pigs. OT: optic tract.
Post-natal programming by the mother
Nursing style
Mother rats show variability in the style of nursing (+/- arched-back nursing (ABN)) and amount of tactile stimulation given to the young – high or low licking and grooming (LG). Offspring of high LG-ABN mothers have reduced HPA axis activation and fearfulness (anxiety) upon stress exposure; these features are not genetically transmitted so this programming of the offspring is epigenetic. The high LG-ABN offspring have increased glucocorticoid feedback sensitivity, and increased GR mRNA expression in the hippocampus, evidently mediated by 5HT7 receptors and nerve growth factor inducible protein-A (NGFI-A), via a promoter adjacent to exon I7 (Meaney and Szyf, 2005).
Paradigm shift
Michael Meaney and colleagues went on to test the hypothesis that the impact of maternal behaviour on GR mRNA expression in the hippocampus involves DNA methylation. They found that the exon I7 site shows much more DNA methylation in all newborns, and in low LG-ABN offspring this persisted, which blocks access to the transcription factor NGFI-A; but the methylation is reversed by cross-fostering to a high LG-ABN mother (Weaver et al., 2004). Hence, more attentive mothering promotes demethylation of the GR gene and thus increases GR expression and glucocorticoid feedback (Meaney and Szyf, 2005) (Figure 10). However, expression of hundreds of genes in the brain is altered by the quality of maternal care via stable, but reversible, epigenetic mechanisms (Weaver et al., 2006). These elegant studies have triggered effort to translate this discovery to understanding the impact of early life experiences in humans on later mental health (Turecki and Meaney, 2016).
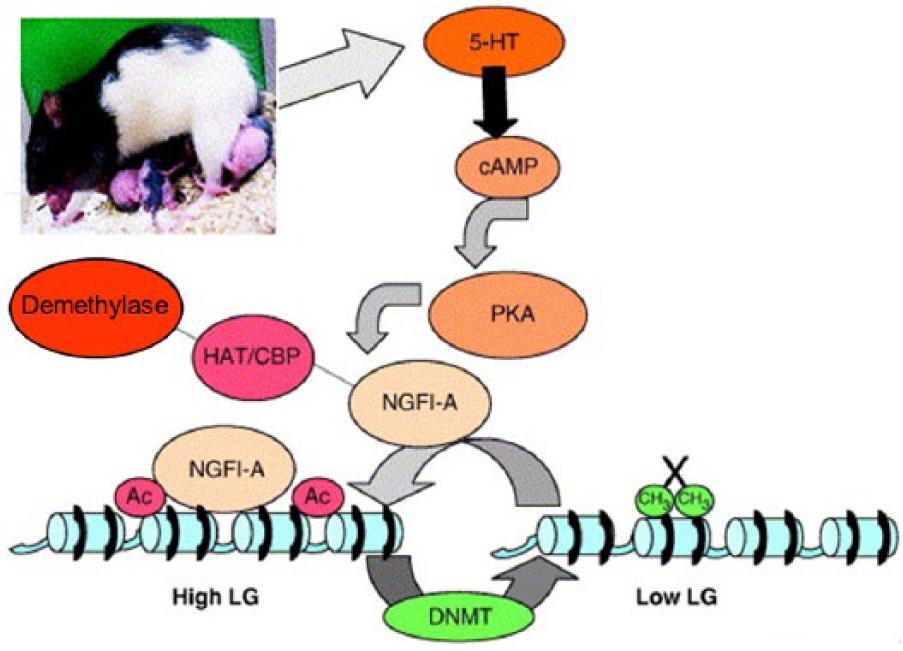
Maternal behaviour programs epigenetic state of glucocorticoid receptor (GR) in offspring brain. Model of mechanism for programming increased expression of hippocampal GR by mother rats showing high levels of pup licking and grooming and arch-back nursing (LG-ABN). In these offspring, compared with those of mothers not showing this style of maternal behaviour, maternal LG behaviour enhances central release of serotonin (5-hydroxytryptamine, 5-HT), which binds to the 5-HT7 receptor which acts via cAMP-protein kinase A (PKA) and CREB-binding protein (CBP) to activate NGFI-A expression. Binding of NGFI-A to the GR promoter recruits histone acetyltransferases (HATS, e.g. CBP) which acetylate histones (Ac), enabling access for demethylase; also the DNA binding site for nerve growth factor inducible-A (NGFI-A) on the GR gene is demethylated, enabling access. Consequently, GR expression is greater, enhancing negative feedback control of the hypothalamic-pituitary-adrenal (HPA) axis by glucocorticoid. Activation of DNA methyltransferase (DNMT) and hence methylation (CH3) would be a mechanism for reducing affinity for NGFI-A and hence GR expression (X).
Caution
Human studies have involved seeking epigenetic marks (DNA methylation, histone acetylation) on specific genes of interest in post mortem brain samples or circulating leukocytes, and correlating findings with early life experience and history of mental disorder (Jawahar et al., 2015). These studies on brain DNA seem to be problematic as methylation of genes of interest cannot be assumed to be stable through life (Witzmann et al., 2012), while the rationale for assuming that leukocyte DNA marks necessarily report about epigenetic modification of the same genes expressed in the brain is unclear and needs robust testing.
Social affiliation: A tale of two voles
Bonding via brain oxytocin and vasopressin receptors
Understanding roles of neuropeptides in the brain in social behaviour has come from studies on two species of North American voles. In a lesson from natural history, the majority of male and female prairie voles were observed to pair for life (more or less), the bonded pairs remain in close contact and are biparental; in contrast, montane and meadow voles are solitary, except for mating, and polygamous. These distinct differences are evaluated in experiments testing preference for a familiar versus an unfamiliar con-specific. As activation of oxytocin release in the mother’s brain after giving birth had been shown to be important in ensuring the rapid onset of maternal behaviour (in rats and sheep; Kendrick et al., 1987; Pedersen and Prange 1979), supposing a role for oxytocin in establishing social relationships was a logical step. Studies in the 1990s showed roles for oxytocin and vasopressin in the brain in social memory and recognition in several species. Prairie voles have been found to have, compared with montane voles, greater densities and much wider distribution of both oxytocin and vasopressin V1a receptors in the brain, assessed by radioligand binding, mRNA and protein expression (Figure 11). Pharmacological studies with site-specific injections of oxytocin and vasopressin and receptor antagonists, and transgenic studies expressing in mice the V1a receptor driven by the prairie vole 5ʹ regulatory region, all indicated the importance of central oxytocin actions in females, and of vasopressin in males in the bi-directional process of pair-bonding in prairie voles (Nair and Young, 2006). The prairie vole has two copies of the avpr1a gene, and the montane vole only one, with interspecies differences in upstream sequences, which seem important in determining anatomical distribution (Nair and Young, 2006). In prairie voles, variability in non-coding regions of the OXTR gene are associated with more intense expression in the nucleus accumbens than in montane voles; hence, a genetic mechanism determines regional differences in oxytocin receptor expression, and bonding behaviour (Young et al., 2001). In prairie voles, a cis-regulatory element is responsible for variation among individuals for the intensity of expression in the nucleus accumbens, which correlates with bonding behaviour (King et al., 2016).

Social affiliation and oxytocin and vasopressin 1a receptors in brain. Autoradiographs of transverse brain sections from, left, prairie (monogamous) and, right, montane (non-monogamous, promiscuous) voles. Sections were radiolabelled with ligands for A, B, oxytocin receptors, or C, D, vasopressin 1a (V1a) receptors. Oxytocin receptor binding is prominent in the nucleus accumbens (NAcc) in the prairie vole (A), and lacking in the NAcc in the montane vole (B); oxytocin receptor binding in the prelimbic cortex (PLC) is similar in the two species. V1a receptor binding is prominent in the ventral pallidum (VP) in the prairie vole (C), and much less dense in the VP in the montane vole (D); V1a receptor binding is similar in the lateral septum (LS) in the two species.
Translation to humans?
From the studies on the importance of differences in oxytocin receptor expression on bonding in voles, an inference was made that in humans actions of oxytocin in the brain on differentially expressed oxytocin receptors might be important in social function, through regulating rewarding aspects of social stimuli (King et al., 2016; Walum et al., 2016). The clear implications for better understanding autism spectrum disorder (ASD) and schizophrenia have prompted much research to test this idea. Approaches used include testing effects of administering oxytocin, measuring circulating oxytocin concentrations and molecular genetics.
Limitations and cautions
Oxytocin administration
The aim of such studies has been to seek effects of exogenous oxytocin on various behavioural measures related to a social challenge, to test hypotheses about oxytocin actions in the brain. Many such studies have used intranasal spray administration of oxytocin solution, in the expectation that the oxytocin will enter the brain. There is no firm evidence that it does; on the contrary, the blood-brain barrier prevents such entry, and oxytocin is not known to have actions on any circumventricular organ. Intranasal administration of oxytocin as an aerosol, but not by spray, to rhesus macaques increases CSF oxytocin concentration, but the efficiency of ‘transfer’ was extremely low (Leng and Ludwig, 2016; Modi et al., 2014); in one human study, an increase in oxytocin concentration in CSF was claimed at 75 min after intranasal oxytocin (n = 3), but with only one placebo control participant (Striepens et al., 2013). Hence, any changes in behaviour after intranasal oxytocin, at least by spray, are likely to be a consequence of peripheral effects following entry into blood; if so, these should be blocked by a peptide oxytocin antagonist, as this will not enter the brain, but evidently this has not yet been tested (Leng and Russell, 2016).
Measuring circulating oxytocin
There are two problems. First is a presumption that concentration of oxytocin in blood indicates oxytocin release in the brain: it will reflect the activity of magnocellular oxytocin neurons (of which only a few might be centrally projecting), but not parvocellular centrally projecting oxytocin neurons. Conversely, as magnocellular oxytocin neurons can be induced to release oxytocin from dendrites without excitation of secretion from the posterior pituitary, lack of increase in circulating oxytocin does not mean lack of central release (Sabatier et al., 2003). Second is that some of the assays used require extraction of peptide, and in many studies this has not been done, giving falsely very high (e.g. 60-fold vs. extracted) readings, as surveyed recently (Brown et al., 2016; Leng and Ludwig, 2016; Leng and Sabatier, 2016; McCullough et al., 2013). Some studies report measurements of salivary oxytocin concentrations; however, it is not clear how this peptide can enter saliva, and if it does, how any changes in rate of saliva secretion are accounted for (Jong et al., 2015).
Molecular genetics: Epigenetic studies
An initial study combining use of post mortem brain tissue and blood cell analyses found that blood lymphocytes and temporal cortex samples from ASD cases showed increased methylation of CpG islands in OXTR regulatory regions (Gregory et al., 2009); the origin of the hypermethylation is not known. However, studies in rats have shown that offspring reared by mothers showing low levels of licking and grooming (low LG) had less OXTR methylation in blood but not in brain (Beery et al., 2016); hence, measurement of epigenetic changes in blood or buccal cells does not reliably reflect changes in the brain. Moreover, OXTR methylation in blood cells changes acutely with stress (Unternaehrer et al., 2012). For these reasons, such studies on the important issue of impact of OXTR methylation in the brain have to be regarded with caution.
Genetic variations
Meta-analysis of studies of genetic variations in the OXTR gene indicates an association of the gene with ASD, and in particular implicates four single-nucleotide polymorphisms (SNPs); the functional consequences of the SNPs are not known. Further studies are needed to address several limitations of the datasets (LoParo and Waldman, 2015). Perhaps, CRISPR/Cas gene editing in model animals, that is, the laboratory rat (Meek et al., 2017), will allow elucidation of the impact of minor changes in a target gene on neuroendocrine function and complex behaviour.
Conclusion
Overall, human studies on the translation of studies of central oxytocin functions in animal models have been embarked on, and inferences often drawn on a background of inadequate information or critical appreciation of neuroendocrine mechanisms, despite the extensive basic biomedical literature (Grinevich et al., 2016). Nonetheless, there is more potential in this area for contributions to social neuroscience, especially in understanding aberrant social functioning, from deep basic neuroendocrine studies (Insel, 2010).
Footnotes
Acknowledgements
I am grateful for stimulation of my interest in neuroendocrinology by Emeritus Professor L. M. Pickford FRS (Mary died on her 100th birthday in 2002), and for guidance in critical thinking from Professor W. E. Watson, my PhD supervisor, who died in 2000. I acknowledge many productive and enjoyable collaborations with undergraduate and postgraduate students, post-doctoral fellows and senior colleagues in the past not quite 50 years.
Declaration of conflicting interests
The author declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author’s research cited here was supported by the Biotechnology and Biological Sciences Research Council, UK.