
Abstract
It is often recommended that patients at risk for gastroesophageal reflux (GER) should remain in a seated or upright position for 30 to 60 min after eating. However, there is no scientific justification for this time frame. The lower esophageal sphincter (LES) is important for preventing GER, and some gastrointestinal hormones are known to regulate the LES. To obtain morpho-functional evidence for the time during which one should assume such a position, we used a rat model to focus on gastrin and cholecystokinin (CCK), the hormones that contract and relax the LES, respectively, and examined the changes in the number of cells immunoreactive to these hormones before and after eating. The present study is based on the deduction that vigorous secretion decreases the intracellular contents of hormones to a level lower than that detectable by immunohistochemistry. After 23 h of fasting (PC-0), rats were fed for 3 min and sacrificed 10 min (PC-10) and 60 min (PC-60) later. The number of gastrin cells was decreased in the PC-10 group; however, in the PC-60 group, the number had recovered to that of the PC-0 group. The number of CCK cells was unchanged in the PC-10 group but was decreased in the PC-60 group. Residual gastric contents were found in all groups, indicating that the LES is relaxed despite these contents and suggesting that there are some risks of gastroesophageal reflux even 60 min after eating.
Introduction
It is often recommended that patients at risk for gastroesophageal reflux (GER) should remain in a seated or upright position for 30 to 60 min after eating (Stockert, 2009). However, there is no scientific justification for this time frame. GER is prevented by several anatomical mechanisms at the junction between the esophagus and stomach (Boeckxstaens, 2005; Castell, 1975; Hyun & Bak, 2011; Miller et al., 2007). Among them, the contribution of the lower esophageal sphincter (LES) is the most critical (Korn, Csendes, & Braghetto, 2011; Miller et al., 2007; Mittal, 1990; Patti, Gantert, & Way, 1997). The LES is regulated by at least two systems, one of which is the parasympathetic nervous system (including the vagus nerve; Boeckxstaens, 2005; Castell, 1975; Korn et al., 2011; Mittal & Balaban, 1997; Patti et al., 1997). The LES is relaxed by inhibitory myenteric neurons (nitric oxide–containing neurons) receiving a certain amount of output from the dorsal motor nucleus of the vagus. However, the LES is contracted by excitatory myenteric neurons (cholinergic neurons) receiving a certain amount of output from the dorsal motor nucleus of the vagus.
The hormones secreted by gastrointestinal endocrine cells also influence the process of contraction and relaxation (Allescher, Stoschus, Wünsch, Schusdziarra, & Classen, 1995; Castell, 1975; Clavé et al., 1998; Cohen & Lipshutz, 1971; Giles, Mason, Humphries, & Clark, 1969; Jennewein, Waldeck, Siewert, Weiser, & Thimm, 1973; Korn et al., 2011; Resin, Stern, Sturdevant, & Isenberg, 1973; Sturdevant & Kun, 1974; Tonini, De Giorgio, & De Ponti, 2004). Hormones such as secretin, cholecystokinin (CCK), and glucagon relax the LES, whereas hormones such as gastrin, motilin, pancreatic polypeptide, and bombesin contract it. Hormones are secreted by exocytosis; the membrane of the secretory granule fuses with the cell membrane, and the content (hormone) is delivered outside the endocrine cells (Yamada, Sugitani, Iwanaga, Yamashita, & Misu, 1980). Accordingly, active secretion decreases the number of secretory granules as well as the content of hormones contained in the cell, resulting in a decrease in the intracellular content of hormones to a level lower than that detectable by immunohistochemistry and a decrease in the number of detectable hormone-containing cells. Therefore, we hypothesize that the number of immunoreactive cells is a surrogate marker for hormone levels; for example, a decrease in the number of cells reflects decreased hormone levels after active secretion.
In the present study, we focused on gastrin and CCK, which are important gastrointestinal hormones with effects on the LES that have been widely investigated, and we deduced contraction/relaxation conditions of the LES by counting the number of gastrin and CCK cells with time after eating. From these results, we show morpho-functional evidence for the time needed to elevate the upper part of the patient’s body.
Materials and Methods
Experimental Animals
Seven-week-old male Sprague-Dawley rats (Japan CLEA Inc., Tokyo, Japan) were used. All animal experiments were approved by the Saga University Animal Care and Use Committee. Animals were housed under controlled conditions of 12 hr of light/12 hr of dark and had access to food (CE-2, a standard laboratory chow) and water ad libitum. During the experiment, animals were fasted but had free access to water for 23 hr from 20:00 p.m., the time when the light was turned off and animals actively moved. After fasting, animals were randomly separated into three groups. The first group (PC-0, n = 3) was sacrificed without having eaten, and the second (PC-10, n = 3) group and third (PC-60, n = 3) group were sacrificed 10 and 60 min after eating for 3 min, respectively. Animals were anesthetized with an intraperitoneal injection of a mixture consisting of 0.15 mg/kg medetomidine hydrochloride, 2 mg/kg midazolam, and 2.5 mg/kg butorphanol tartrate. Then, they were euthanized by perfusion of physiological saline through the ascending aorta to flush out the blood. The stomach was dissected out continuously with the esophagus and duodenum (each approximately 2 cm long). After washing the inside of the stomach with saline, approximately 10-mL buffered formalin was poured into the esophagus–stomach–duodenum tract, which was ligated at both the upper and lower ends using a syringe inserted into the esophagus. Then, tissue blocks from the pyloric region to the upper duodenum were trimmed and cut longitudinally. They were dehydrated with graded ethanol and embedded in paraffin. Six-micrometers-thick sections containing the pyloric region of the upper duodenum were consecutively processed for immunohistochemistry.
Immunohistochemistry
The rabbit polyclonal antibody against eight amino acids (DYMGWMDF) from the 96 to 103 amino acid sequence of CCK (Cambridge Research Biochemicals, Cambridge, England) was used in the present study. According to the manufacturer’s data sheet, this antibody recognizes not only CCK but also gastrin; five out of eight amino acid sequences (GWMDF) of this antigen correspond to the 99 to 103 amino acid sequence of gastrin. Therefore, we used this CCK antibody for detection of both CCK and gastrin.
Deparaffinized sections were subsequently incubated with 2% normal donkey serum to block nonspecific reactions for 1 hr, rabbit antibody to CCK (diluted 1:10,000) for 2 hr at room temperature and overnight at 4℃, biotinylated donkey anti-rabbit IgG (diluted 1:200; Jackson Immunoresearch Laboratories, West Grove, PA), and avidin–biotin–peroxidase complex (diluted 1:200; Vectastain ABC kit; Vector Laboratories, Burlingame, CA) for 1 hr. Between each step, sections were rinsed with phosphate-buffered saline three times for 5 min each. Then, sections were treated with 0.02% diaminobenzidine tetrahydrochloride (DAB; Dojindo, Kumamoto, Japan) in 0.05 M Tris buffer (pH 7.6) containing 0.005% H2O2, rinsed with Tris buffer and distilled water, and covered.
Observation and Photomicrography
Photomicrographs of gastrin cells in the pyloric regions and of CCK cells in the duodenum were taken using an objective lens with a magnification of 10× under an Olympus BX51 light microscope equipped with an Olympus DP71 digital camera (Tokyo, Japan). The number of gastrin cells was counted on 10 randomly photographed fields per rat and that of CCK was counted on 5 to 10 serial fields. A public domain Java image-processing program (ImageJ v. 1.49e from National Institutes of Health, Bethesda, MD; http://rsb.info.nih.gov/ij/) was used to count cell numbers and to measure the length of the lamina muscularis mucosae. Image-processing software (Adobe Photoshop) was used to make minor adjustments to brightness and contrast of digitally captured microscopic images.
Statistical Analysis
The number of gastrin cells and the number of CCK cells per 1 mm of lamina muscularis mucosae from rats in the PC-0, PC-10, and PC-60 groups were calculated. When counting, only the cells with a visible nucleus were approved. All data were expressed as mean ± standard error of the mean (SEM). Comparisons were performed using Student’s t test. p < .05 was considered statistically significant.
Results
Although the rats bit chow only once or twice during the 3-min meal time after fasting, residual gastric content was found, even in the PC-60 group.
Gastrin-secreting cells were found at the base of the pyloric gland (Figure 1). For each 1 mm of lamina muscularis mucosae of the PC-0, PC-10, and PC-60 groups, there were 64.2 ± 3.3, 39.5 ± 1.2, and 114.5 ± 23.0 (mean ± SEM) gastrin cells, respectively (Table 1). The number of cells in the PC-10 group was significantly decreased compared with that of the PC-0 group (p = .002) and the PC-60 group (p = .031). The numbers found in the PC-0 and PC-60 groups were not significantly changed. There was a tendency for the gastrin cells of the PC-10 group to be light in color in the brown benzidine (Figure 1).
Distribution of gastrin cells in the pyloric gland before and after eating following 23 hr of fasting. Sections are from an animal that did not eat (A; PC-0 group), an animal 10 min after eating (B; PC-10 group), and an animal 60 min after eating (C; PC-60 group). Note that the number of gastrin cells (brown) in the PC-10 group is lower than that in the PC-0 and PC-60 groups. Hematoxylin counterstaining. Bar = 100 µm. The Numbers of Gastrin and Cholecystokinin Cells Before and After Eating following 23 hr of Fasting. p < 0.01 compared with PC-0. **p < 0.05 compared with PC-10.

The CCK-secreting cells were found in the epithelium of the intestinal villus and crypt (Figure 2). The numbers of CCK cells per 1 mm of lamina muscularis mucosae of the PC-0, PC-10, and PC-60 groups were 8.5 ± 0.5/mm, 8.2 ± 1.8/mm, and 3.9 ± 0.8/mm, respectively (Table 1). The number of cells in the PC-60 group was significantly decreased compared with that in the PC-0 group (p = .008), although no significant change was observed between the PC-0 and PC-10 groups.
Distribution of cholecystokinin cells in the duodenum before and after eating following 23 hr of fasting. Sections are from an animal that did not eat (A; PC-0 group), an animal 10 min after eating (B; PC-10 group), and an animal 60 min after eating (C; PC-60 group). Note that the number of cholecystokinin cells (arrows) in the PC-60 group is lower than that in the PC-0 and PC-10 groups. Hematoxylin counterstaining. Bar = 100 µm.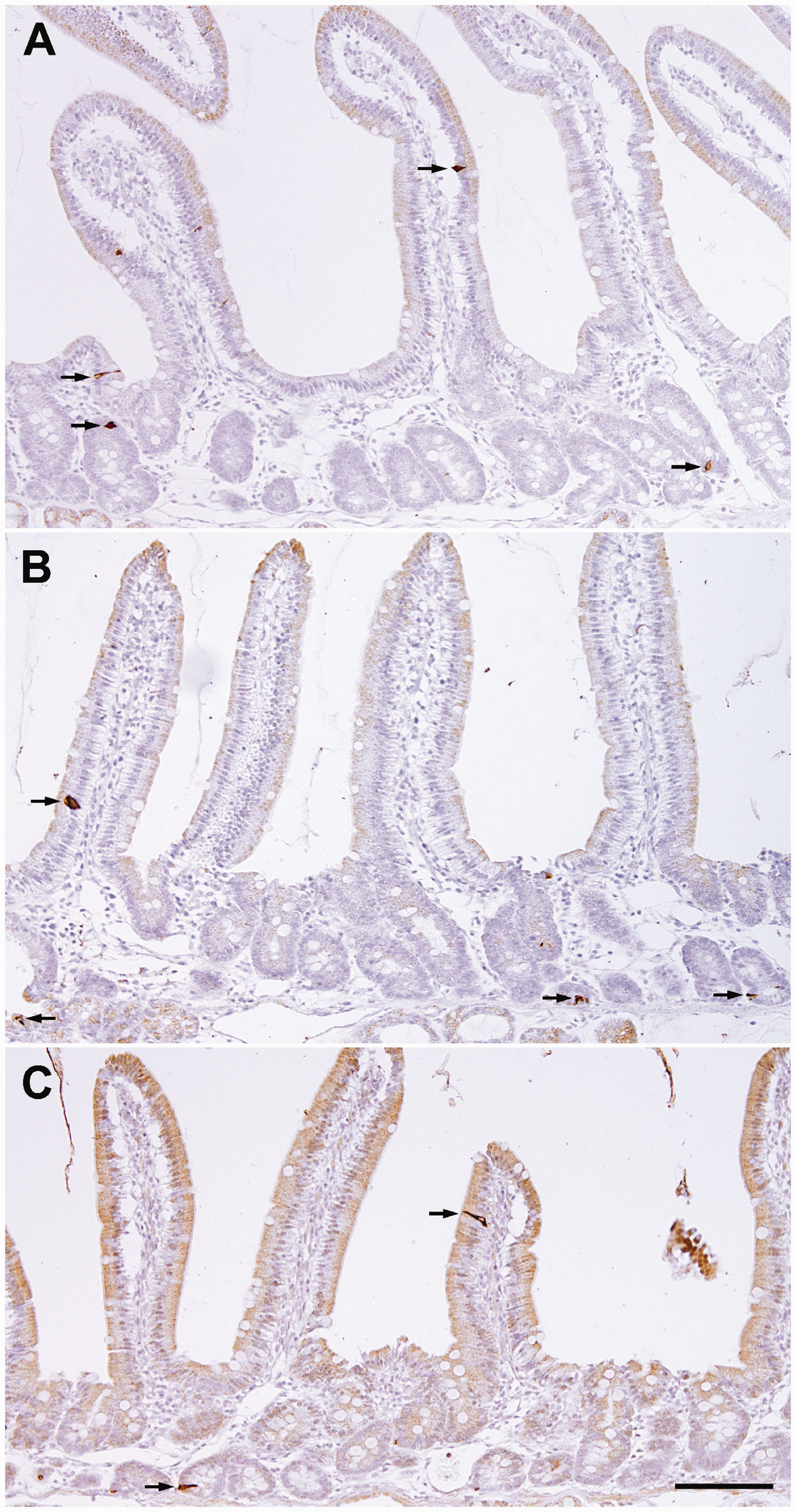
Discussion
The present study revealed the changes in the number of gastrin cells and CCK cells with the time after eating. The authors believe that the decreased number of immunoreactive cells in the present study may be due to the level of intracellular hormones being lower than that able to be detected by immunohistochemistry, but not due to the disappearance of cells. A previous electron microscopic study (Yamada et al., 1980) showing exocytotic images of secretory granules after eating supports this idea. The fact that the number of gastrin cells returned to baseline after eating may also confirm this idea.
It is well known that gastrin is secreted during the digestion process in the stomach and facilitates gastric acid secretion from the parietal cells in the fundic gland. After ingested food undergoes gastric digestion and moves into the small intestine, various food components such as peptides, fat, and acid stimulate secretion of CCK, a hormone known to contract the gallbladder and secrete pancreatic digestive enzymes. Therefore, it is estimated that the animals in the PC-10 group in the present study were just undergoing active gastric digestion and that those in the PC-60 group were undergoing intestinal digestion.
The number of gastrin cells was significantly decreased in the PC-10 group, and that in the PC-60 group returned to the number in the PC-0 group. It is likely that the decreased number of gastrin cells in the PC-10 group was due to the decreased content of gastrin following vigorous secretion, and that the LES at this time was contracted by gastrin (Allescher et al., 1995; Castell, 1975; Cohen & Lipshutz, 1971; Giles et al., 1969; Jennewein et al., 1973; Korn et al., 2011; Sturdevant & Kun, 1974). Gastrin can be replenished in the cells in the PC-60 group, resulting in the number being comparable to that in the PC-0 group. However, a significant decrease in the number of CCK cells was found in the PC-60 group in the present study. This also may be due to the decreased content of CCK following secretion. Therefore, it is conceivable that the LES was relaxed by CCK (Castell, 1975; Clavé et al., 1998; Hirsch et al., 2002; Korn et al., 2011; Resin et al., 1973; Sturdevant & Kun, 1974; Tonini et al., 2004) in the PC-60 group.
Previous observations showed that it takes more than 60 min to send most of the ingested meal from the stomach to the duodenum (George, 1968; Hunt & Spurrell, 1951). In the present study, residual contents were observed in the stomach of the rats, even in the PC-60 group, despite only one or two bites of chow for 3 min after fasting. At this time, the number of detectable CCK cells was decreased, presumably because of vigorous secretion, and the LES may have been relaxed by CCK. Therefore, there is a risk of GER even 60 min after eating. This indicates that patients should also be carefully monitored when they lie on their back for more than 60 min after having eaten.
The present data were obtained from healthy young adult rats (seven weeks old) 10 and 60 min after eating normal laboratory chow. However, GER is frequently noted in older patients as well as in patients with weakened muscles, neurological disorders, or anatomical abnormalities. Further studies are necessary to assess high-quality nursing care using various parameters that are considered to be involved with a high risk of GER, such as older age, food types (low-fat/high-fat, solid, or liquid), and time after eating. It is evident that nutrition-dense meals move to the duodenum slower than less dense meals (Hunt & Stubbs, 1975), and that solid food moves to the duodenum slower than liquid food (Christian, Moore, Sorenson, Coleman, & Weich, 1980). It is also necessary to take into consideration gastrointestinal hormones other than gastrin and CCK, such as secretin and glucagon for relaxing the LES and motilin, pancreatic polypeptide, and bombesin for contracting the LES (Castell, 1975; Jennewein et al., 1973; Korn et al., 2011).
Footnotes
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) received no financial support for the research, authorship, and/or publication of this article.