
Abstract
Background:
Irreparable posterosuperior rotator cuff (PSRC) tears have been shown to result in shoulder pain and loss of function.
Purpose/Hypothesis:
The purpose of this study was to determine the effect of the loss of active or passive function of the PSRC on compensatory muscle loads in the deltoid and the remaining rotator cuff. It was hypothesized that both deactivation and resection of the PSRC would result in load increases in the remaining muscles and that resection of the PSRC would result in even higher required compensatory forces than would mere deactivation.
Study Design:
Controlled laboratory study.
Methods:
A total of 7 cadaveric shoulders were tested using a biomechanical shoulder simulator with 10 independently controlled actuators for various muscles (anterior, middle, and posterior deltoid; inferior and superior subscapularis; latissimus dorsi; pectoralis major; teres minor; supraspinatus; and infraspinatus) and 3-dimensional motion tracking. The muscle loads representing the latissimus dorsi and pectoralis major were each held constant, and the remaining muscle actuator forces required to abduct the arm in the scapular plane were determined. The actuator forces corresponding with arm elevation from 20° to 65° were compared at 5° increments for 3 testing conditions: (1) intact, active PSRC; (2) intact, deactivated PSRC; and (3) resected PSRC and shoulder capsule.
Results:
In both the deactivated and resected states, the teres minor showed a significant increase in required muscle forces through nearly the entire tested range of arm elevation compared to the active state. This was also the case for the subscapularis but only at higher elevation angles. The deltoid demonstrated increased muscle forces of at least 1 of its subunits between 25° and 55° of elevation when comparing the deactivated state or resected state to the active state. However, through nearly the entire range of elevation, no statistically significant differences were found between the deactivated and resected states for any of the actuator loads representing muscle forces.
Conclusion:
The loss of active function of the PSRC led to compensatory loads on the remaining rotator cuff and deltoid, regardless of the passive presence of the PSRC as a supposed subacromial spacer.
Clinical Relevance:
The findings encourage the exploration of treatment procedures that mimic the active function of the PSRC when the rotator cuff itself is irreparable.
Patients with massive rotator cuff tears but with intact force couples are still able to control the center of rotation to maintain overhead motion. In case of a loss of functioning force couples, a subset of patients are debilitated by pseudoparalysis of the shoulder with the inability to actively raise the arm. 1,2 The goal of surgical management is to restore the active function of the rotator cuff as completely as possible, which can successfully reverse pseudoparalysis, as reported by Denard et al 6 and Oh et al. 34 When chronic massive posterosuperior rotator cuff (PSRC) tears are technically irreparable, surgical preservation or restoration of joint control becomes challenging. In addition to partial repair, various surgical approaches have been reported for the treatment of irreparable PSRC tears, including (1) pain-reducing procedures such as arthroscopic debridement with tenotomy or tenodesis of the long head of the biceps tendon, (2) active procedures such as tendon transfer and interposition grafting, and (3) passive spacing procedures such as superior capsule reconstruction and balloon spacers. 3,13,15,22,26,27,33,40 None of these procedures is considered optimal for the treatment of irreparable PSRC tears because all alternatives to complete repair have shown inferior clinical outcomes and higher rates of postoperative complications over time. 4,21,22,24,26
The active and passive roles of the PSRC in shoulder kinematics are not yet fully understood. While the necessity of compensatory muscle forces to maintain arm motion in different rotator cuff tear conditions has been shown before in biomechanical studies and computer simulations, 7,19 the differentiation between active (contraction) and passive (spacing) effects of the PSRC on compensatory muscle forces is of interest when evaluating biomechanics of the aforementioned active and passive surgical procedures.
The purpose of this study was to determine the effect of the loss of active or passive function of the PSRC on compensatory muscle loads in the deltoid and the remaining rotator cuff when re-creating PSRC-deficient conditions in a cadaveric model. We hypothesized that both deactivation and resection of the PSRC would result in an increase in loads in the remaining rotator cuff muscles and deltoid to raise the arm compared to the intact, active state. We further hypothesized that resection of the PSRC would result in even higher required compensatory forces than would mere deactivation.
Methods
Biomechanical Setup
A total of 7 human cadaveric left-sided shoulders (donor mean age, 55 years) were biomechanically evaluated using the newest implementation of a previously published shoulder simulator 16,17,36,39 across 2 testing facilities (Figure 1). All specimens were donated to a tissue bank (Science Care) for medical research and then purchased by our institution.

The shoulder motion simulator with a cadaveric specimen loaded for testing at the starting position for glenohumeral elevation (20° of elevation in the plane of the scapula, 50° of humeral axial external rotation in relation to the scapula). Optical trackers provided joint angle feedback, and cables routed from tendons of the insertion through pulleys (center right) to linear actuators (lower right) provided muscle tension forces to achieve desired joint positions.
This simulator has been shown to reproduce arm rotation movements about all 3 axes, accurate to within 1.5° of targeted values; replicate muscle lines of action and generate necessary muscle forces for up to 12 muscles; represent the scapulothoracic orientation by positioning the scapula with lockable multiaxis-positioning gimbals; and report biomechanical outputs such as muscle forces corresponding to 3-dimensional glenohumeral orientations during simulated arm movements.
Specimen Preparation
The arms were prepared via potting of the scapulae and rigidly pinning the elbows and wrists such that the elbow was held in full extension, the forearm was held in external rotation with the thumb aligned with the lateral epicondyle, and the wrist was held neutral. High-strength cables were attached to each humerus at the tendinous insertion sites of the pectoralis major; latissimus dorsi; teres minor; superior and inferior subscapularis; infraspinatus; supraspinatus; and anterior, middle, and posterior deltoid and were connected via a pulley system to linear actuators (N2 Series Electric Cylinder; Kollmorgen) equipped with inline load cells (Model SSM 500 lbf; Interface) operating under closed loop gain controls. Rigid body-tracking infrared light-emitting diode triads (Optotrak Certus; Northern Digital) were attached to the humerus, forearm, and scapula positioning frame. Throughout testing, the exposed soft tissues of the specimen were sprayed at regular intervals with 0.9% saline mixed with glycerin to maintain moisture and prevent tissue desiccation.
Biomechanical Testing
The scapula was placed in a normal position (10° of upward rotation and 10° of anterior tilt), 25 elevation of the arm was performed in the scapular plane (constant plane of elevation target of 0°), and axial rotation of the humerus relative to the scapula was maintained (constant humeral axial external rotation target of 50°). Motion of the humerus relative to the scapula (elevation angle, angle of the plane of elevation, axial humeral rotation angle) was actively controlled by the muscle actuators during the arm elevation movement. Testing was performed with the PSRC (infraspinatus and supraspinatus) in (1) intact and active, (2) intact but deactivated, and (3) resected states. In the deactivated state, the appropriate actuators were disconnected from the supraspinatus and infraspinatus connection points, but soft tissues were not disturbed. For the resected state, the 2 tendons were resected along with the underlying capsule. Otherwise, the testing conditions were identical for the deactivated and transected states.
The actuator control methodology avoided a priori muscle forces for angle control of the glenohumeral joint in favor of kinematic feedback of the glenohumeral joint, determining the direction and velocity in which the primary actuators moved. For testing, actuators were assigned functions in a muscle function matrix, corresponding to the following muscles in angle control: middle deltoid = positive elevation, anterior deltoid = positive elevation and positive plane of elevation (forward flexion), posterior deltoid = positive elevation and negative plane of elevation (extension), superior subscapularis = positive (internal) rotation, and infraspinatus = negative (external) rotation.
The actuator representing the inferior subscapularis was defined to act at 50% of the force of the superior subscapularis. Actuators for the latissimus dorsi and pectoralis major were each held at a constant force of 15 N to represent tonic tension and to assist with adduction/stabilization. 35 To avoid control redundancy, only 1 muscle was assigned to be the primary controller of each degree of freedom in each direction. Additionally, the control software incorporated gain settings to ensure a minimum muscle tension limit (tonic loading) and a maximum muscle tension limit (extraphysiological loading).
When the PSRC was active, the supraspinatus actuator acted as an elevator with a force always equal to 25% of the middle deltoid (the primary elevator), and the infraspinatus actuator acted in angle control to provide external rotation, with the teres minor defined to act at 30% of the infraspinatus. When the PSRC was deactivated or resected, the teres minor actuator acted in angle control alone to provide external rotation.
For each condition, the arm was elevated in the scapular plane from 20° to 65° in 5° increments and held long enough for the muscle forces to balance while maintaining the target plane of elevation and humeral axial rotation angles (Figure 2). When all control parameters were within a specific variance, data were recorded, and the system drove to the next position in the sequence. Overall, 5 trials for each test condition were performed, and the average of the various muscle forces for the last 3 trials was analyzed.

The simulator achieving 65° of glenohumeral joint elevation while maintaining the scapular plane of elevation and humeral axial rotation targets under muscle actuator control.
Statistical Analysis
For each of the 7 tested specimens, muscle forces of the teres minor, superior subscapularis, anterior deltoid, middle deltoid, and posterior deltoid were calculated for each condition as the average actuator values for those muscle lines from the final 3 trials at each increment of arm elevation. The overall mean and standard deviation for the muscle forces for each condition were calculated as the mean and standard deviation of the individual specimen’s average muscle forces at each arm elevation increment.
After confirming the normality of the data, statistical analysis was conducted using 1-way analysis of variance and post hoc Tukey pairwise comparisons with corrections between conditions at each increment of arm elevation. The significance level was set at P < .05. This study was adequately powered to determine a 45-N difference between test groups. MiniTab 19 statistical software was used (MiniTab LLC, State College, PA USA).
Results
Effect of Deactivated and Transected PSRC on Deltoid Forces
The mean maximum anterior, middle, and posterior deltoid forces in the active, deactivated, and resected PSRC during glenohumeral abduction from 20° to 65° are illustrated in Figure 3.

Lines and whiskers showing mean and standard deviation forces (N) for the deltoid actuators to elevate the arm at 20° to 65° for the active, deactivated, and resected posterosuperior rotator cuff states.
Effect of Deactivated and Transected PSRC on Remaining Rotator Cuff Forces
The mean maximum forces for the subscapularis and teres minor in the active, deactivated, and resected PSRC during glenohumeral abduction from 20° to 65° are illustrated in Figure 4.
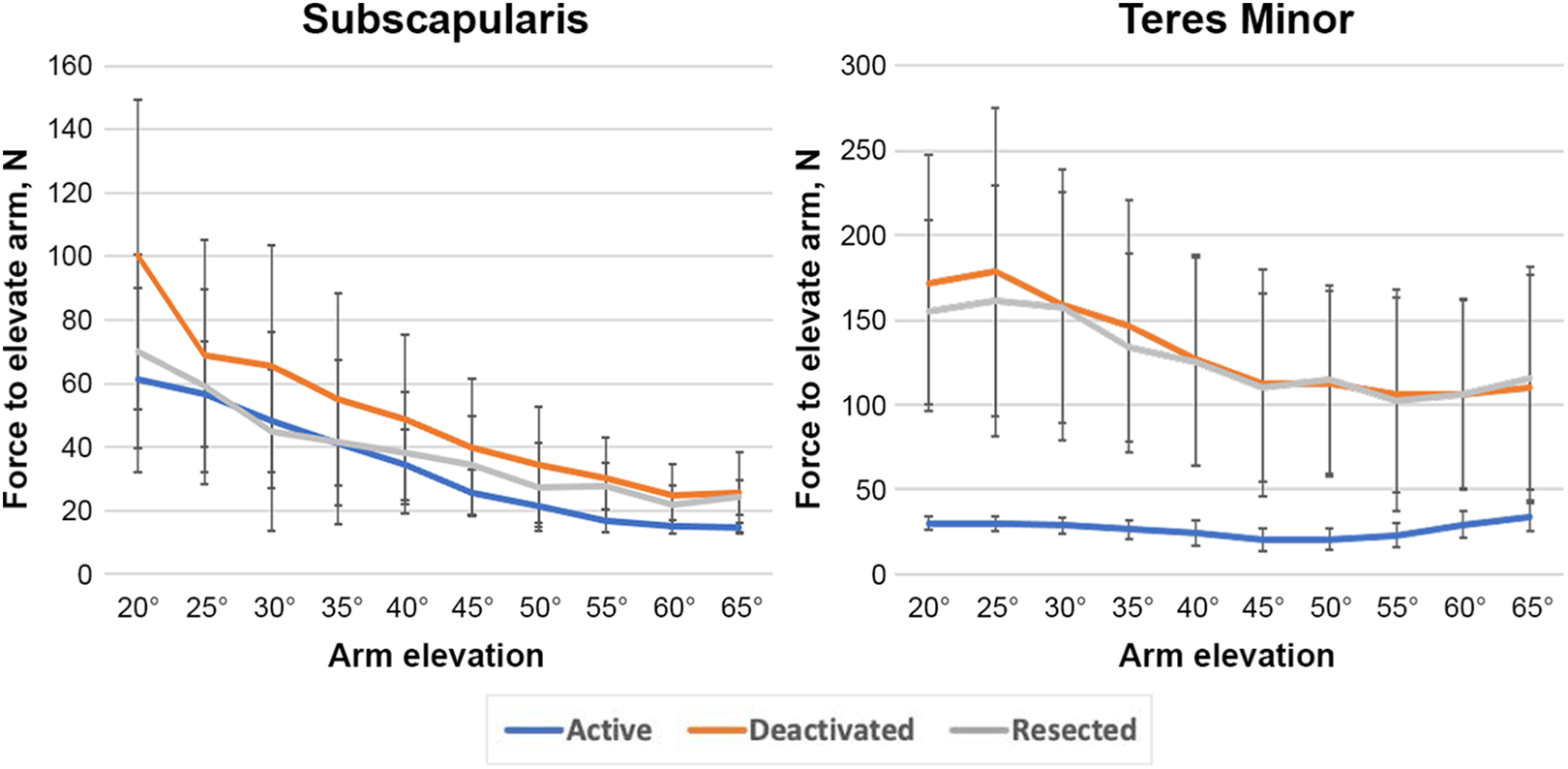
Lines and whiskers showing mean and standard deviation forces (N) for the superior subscapularis and teres minor actuators during elevation of the arm at 20° to 65° for the active, deactivated, and resected posterosuperior rotator cuff states.
Table 1 shows the results of comparisons in muscle forces between the active, deactivated, and resected states at arm elevation angles from 25° to 60°. Through nearly the entire range of arm elevation, the teres minor showed a significant increase in required muscle forces (P = .002-.031) when comparing the active versus deactivated state and the active versus resected state. For the subscapularis, this was only the case at higher elevation angles. The deltoid demonstrated increased muscle forces in its middle subunit at 20° and at 30° to 40° when comparing the active versus deactivated state and at 20° to 45° when comparing the active versus resected state. No statistically significant differences were found between the deactivated and resected states for any of the actuator loads representing muscle forces at any elevation angle studied.
P Values From Comparison of Muscle Forces Between Specimens With an Active, Deactivated, and Resected Posterosuperior Rotator Cuff During 20° to 65° of Arm Elevation a

a Bolded P values indicate a statistically significant difference between posterosuperior rotator cuff states (P < .05). DLTa, anterior deltoid muscle; DLTm, middle deltoid muscle; DLTp, posterior deltoid muscle; SSC, subscapularis; TM, teres minor.
Discussion
The main finding of the current study is that the passive stabilizing function of the PSRC and capsule seemed to be negligible in this biomechanical shoulder model with active muscle loading. Throughout the entire range of elevation studied, no significant differences between the deactivated and resected PSRC were detectable in terms of compensatory muscle forces needed during elevation of the arm for the remaining rotator cuff and middle deltoid.
When comparing the deactivated and resected states with the active state, the teres minor, subscapularis, and deltoid showed a significant increase in compensatory muscle forces at certain points throughout elevation. Therefore, the initial hypothesis that both deactivation and resection of the PSRC would result in an increase of loads to the remaining rotator cuff and deltoid was confirmed. The second hypothesis that resection of the PSRC would result in even higher required compensatory forces than would mere deactivation was not supported.
Dyrna et al 7 studied required deltoid muscle forces during dynamic shoulder abduction in rotator cuff tear progression. In their cadaveric biomechanical analysis of 12 shoulders, they demonstrated that shoulders with rotator cuff tears require more compensatory deltoid forces to prevent the loss of abduction motion. While this study investigated the compensatory role of the deltoid muscle in a rotator cuff tear model via static muscle loading, 7 the current study aimed to closer reflect in vivo conditions via actively controlled loading of the deltoid and (remaining) rotator cuff during elevation. While the findings of Dyrna et al 7 were confirmed, the current study results further highlighted the compensatory role of the remaining rotator cuff as well. Indeed, the remaining rotator cuff muscles showed an increase in compensatory loads in the case of a deactivated or resected PSRC. It seems reasonable that a significant increase in muscle forces was observed for the teres minor throughout almost the entire range of elevation as the only remaining external rotator, which has a line of action to compensate for the loss of the active infraspinatus as the major external rotator. The subscapularis showed increased loads only at higher angles of elevation. These findings are in line with previous biomechanical and clinical investigations on different rotator cuff tear patterns, which showed that the remaining rotator cuff needs to compensate for the torn tendons. 12
Previous studies have indicated that the passive spacer 23 or tuberoplasty 30 effect seen in superior capsule reconstruction 22,27 –29,37 or balloon spacers 5,13,18,20,24 is beneficial to improve clinical outcomes possibly by recentering the humeral head (decrease superior translation) and decreasing muscle loads to the remaining rotator cuff and deltoid in superior rotator cuff–insufficient shoulders. According to Werner et al, 42 deactivation of the PSRC in vivo via a suprascapular nerve block has no effect on superior translation of the humeral head, suggesting that the deactivated PSRC acts as a subacromial spacer to prevent the humeral head from upward migration during abduction. However, in a second study, Werner et al 41 showed that deactivation of the PSRC in vivo does not increase subacromial pressure during active movements including abduction, indicating that it is not the spacing effect of the PSRC that keeps the humeral head centered; otherwise, increases in subacromial pressure during abduction would have been observed. The present study showed that both deactivation and resection of the PSRC lead to a significant increase in compensatory loads on the remaining muscles during elevation. Because the passive presence of an intact but deactivated PSRC alone biomechanically did not differ from the resected PSRC state in terms of compensatory muscle loading, the role of the PSRC as a subacromial spacer is further put into question. Based on these in vivo and in vitro findings, it seems that the compensatory loads on the remaining rotator cuff muscles and deltoid play a major role in keeping the humeral head centered and enabling arm elevation. On the other hand, the mere passive presence of the PSRC as a subacromial spacer does not seem to be of importance, as compensatory loads on the remaining muscles increase as soon as the active function of the PSRC is lost. In terms of therapeutic options available to treat irreparable PSRC tears, these findings suggest that only restoration of the active function of the PSRC is able to relieve the remaining rotator cuff and deltoid from excessive compensatory loads, thus supporting the role of partial rotator cuff repair and encouraging the further exploration of tendon transfer or interposition grafting, which aim to actively mimic the function of the PSRC. 8 –11,14,15,31 –33,38
Limitations
While this study revealed interesting findings, there were limitations. First, shoulder motion was performed only in the scapular plane rather than in multiple planes, which represent a greater range of in vivo conditions. 25 Second, the limited number of tested specimens likely did not reveal all the observable biomechanical differences as statistically significant. Nonetheless, the paired within-specimen statistical approach was able to identify several significant differences, which were sufficient to test both our hypotheses even with this limited sample size. Third, the subscapularis showed a definite trend (although not significant) toward lower compensatory loads with a resected PSRC than with a deactivated PSRC; the explanation for this observation remains speculative. Despite these limitations, this study provides valuable information to clinicians treating technically irreparable PSRC tears.
Conclusion
This biomechanical study demonstrated that the loss of active function of the PSRC led to compensatory loads on the remaining rotator cuff and deltoid, regardless of the passive presence of the PSRC as a supposed subacromial spacer. Clinically, the study findings encourage the exploration of treatment procedures that mimic the active function of the PSRC when the rotator cuff itself is irreparable.
Footnotes
Acknowledgment
The authors greatly thank Gerome Miller and Aniruddh Nayak for their mechanical testing contributions.
Final revision submitted January 23, 2022; accepted March 2, 2022.
One or more of the authors has declared the following potential conflict of interest or source of funding: Specimens used in this study were donated by DePuy Synthes Mitek. P.M. has received consulting fees from Arthrex, DePuy Synthes Mitek, and NCS Lab. A.C. has received consulting and speaking fees from Arthrex, DePuy Synthes Mitek, Wright Medical, and Zimmer Biomet. S.M.G., M.Z.S., and D.B.S. are employees of DePuy Synthes Mitek Sports Medicine and stockholders of Johnson & Johnson. J.M.T. has received education payments from Elite Orthopaedics, Goode Surgical, and Impact Ortho; consulting fees from Arthrex, Medical Device Business Services, and Zimmer Biomet; and speaking fees from Arthrex. AOSSM checks author disclosures against the Open Payments Database (OPD). AOSSM has not conducted an independent investigation on the OPD and disclaims any liability or responsibility relating thereto.
Ethical approval was not sought for the present study.