
Abstract
Breast cancer is among the most common malignant tumors. It is the second leading cause of cancer mortality among women in the United States. Curcumin, an active derivative from turmeric, has been reported to have anticancer and chemoprevention effects on breast cancer. Curcumin exerts its anticancer effect through a complicated molecular signaling network, involving proliferation, estrogen receptor (ER), and human epidermal growth factor receptor 2 (HER2) pathways. Experimental evidence has shown that curcumin also regulates apoptosis and cell phase–related genes and microRNA in breast cancer cells. Herein, we review the recent research efforts in understanding the molecular targets and anticancer mechanisms of curcumin in breast cancer.
Introduction
Breast cancer is the most commonly diagnosed malignant tumor and the second leading cause of cancer mortality among women in the United States. 1 About 70% of breast cancers are classified as estrogen receptor (ER) positive and could be treated with antiestrogens.2,3 Recently, growing evidence has indicated that the application of a single molecule can hardly suppress the cross talk and negative feedback loops in the complex cancer cellular networks. 4 Therefore, drug combinations targeting different pathology signaling pathways have been considered as a major trend in drug design and discovery for killing endocrine-resistant breast cancer cells. 5 Yet, both the research and clinical applications are still in great need of effective candidates for better treatment of breast cancer.
Curcumin is a hydrophobic polyphenol derived from turmeric, a traditional Indian spice 6 ( Fig. 1 ). Curcumin has been used as an ethnic drug for the treatment of diverse diseases. Particularly, curcumin has been recognized as an effective anticancer agent that regulates multiple intracellular signaling pathways, including transcription factors (e.g., STAT3, NF-κB, and AP-1), receptors (e.g., IL-8, HER2, and CXCR4), kinases (e.g., EGFR, ERK, and JAK), cytokines (e.g., TNF, IL, and MIP), enzymes (e.g., MMP, iNOS, and GST), and growth factors (e.g., EGF, NGF, and HGF). 7 Yet, the discussions on curcumin’s anticancer effects have only been available in the last few decades. Sharma et al. 8 succeeded in treating colorectal cancer with curcumin in phase I, and Cruz-Correa et al. 9 used curcumin in the treatment of patients who were diagnosed with familial adenomatous polyposis. In an experimental investigation of mammary cancer induced by 7,12-dimethylbenzanthracene (DMBA), curcumin significantly decreased the initiation of mammary adenocarcinoma on the fourth day after DMBA administration. 10 Furthermore, a large number of studies on cancer prevention at different stages have indicated curcumin as a favorable agent for cancer chemoprevention, used both alone and in combination with other drugs.11-13
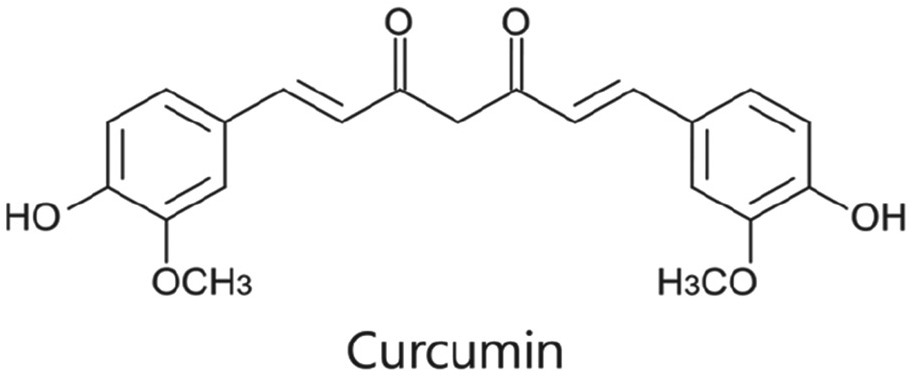
Chemical structure of curcumin.
Although healthy lifestyles, including balanced diets, regular exercise, alcohol reduction, smoking cessation, and stress management, have proved their benefits in preventing multiple kinds of cancer,14-18 the U.S. Food and Drug Administration (FDA) approval of tamoxifen, a selective estrogen receptor modulator (SERM), is considered to be a milestone in breast cancer chemoprevention. 19 When a potential drug compound is under investigation, the utmost importance should always be the safety issue. In this context, unlike synthetic compounds, natural products present in fruits and vegetables possess natural advantages. As such, the numerous effects of curcumin, including antioxidation and anti-inflammation, have called for extensive research efforts in recent years. In the following parts of this article, we review and summarize the recent publications in eight categories according to curcumin diversified functions on breast cancer.
Techniques for Extraction and Purification of Curcumin
Curcumin longa L. (turmeric) contains 2%–9% curcuminoids, including curcumin, demethoxycurcumin and bis-demethoxycurcumin, and cyclic curcumin. Among them, curcumin is considered the most important biomedical molecule. 20 Since 1815, when the first report of extraction and purification of curcumin was published, the curcumin extraction method has been continuously improving and advancing. Organic solvents, commonly reported for curcumin extraction, involve hexane, ethylacetate, acetone, and ethanol. Compared with zone-fining and dipping methods, ethanol extraction is a simpler process with a higher extraction ratio. A high extraction ratio also happens in Soxhlet extraction, ultrasonic extraction, and microwave extraction, which indicates that these techniques are suitable for industrial application.21-26 Moreover, some articles showed that pulse ultrasonic and microwave-assisted extraction lead to a high curcumin ratio. 24 Another common method uses acid-alkali, 27 the principle of which is to dissolve the phenolic hydroxyl group of curcumin with sodium hydroxide at pH 7.0. This method could also separate the starch in the crude extraction product. There are also a few reports on compound enzyme extraction where curcumin is treated with enzymes such as cellulose enzyme and pectinase, which could degrade the cytoderm and intercellular substance.28,29 However, due to the cost and complexity of the process, the enzyme-assisted method has not been commercially adopted.
Among all the protocols, the primary recommendation for laboratory research is supercritical CO2 fluid extraction (SFE). Fluid under the supercritical state with supercritical temperature and pressure is regarded as high density, with characteristics of both gas and liquid. The viscosity of supercritical fluid (SF) is similar to that of gas, while its diffusion coefficient is 10–100 times greater that of than liquid, which makes it an excellent choice of extractant. CO2 is more frequently used than ethylene and ethane hexane methyl methane.30,31 The normal operating condition for this process is at a pressure of 25 MPa and a temperature of 318K, which requires 4 h of static extraction and 5 h of dynamic extraction with cosolvent of 30% ethyl alcohol in 3.5 L/min CO2 flow. 32
Curcumin could be purified from crude curcumin (a mixture of curcuminoid) by column chromatography. It is a technology that uses different organics such as activated carbon, activated clay or silica with mixtures of solvent like dichloromethane/acetic acid or methanol/chloroform/dichloromethane, and ethanol/methanol mixtures as eluents to yield fractions.33-38 Among these, macroporous resin column chromatography is widely adopted. Macroporous resin was invented in 1964, with holes of 100-1000 nm distributed on the surface area. Combined with styrol and propionate, phenylethylene forms a porous polymer with a screening function. 39 A recent paper introduced a novel method for curcumin preconcentration. The molecularly imprinted polymers (MIPs) based on magnetic multiwalled carbon nanotubes possessed excellent selectivity toward curcumin. Other advanced techniques based on functional comonomers, such as thermoresponsive magnetic molecularly imprinted polymers (TMMIPs), have also been developed with high reproducibility and stability for selective curcumin extraction.40-42
Meanwhile, high-performance liquid chromatography (HPLC) is widely used for the detection and estimation of curcumin. Absorption detectors with solvent containing acetonitrile/water and chloroform/methanol at the mobile phase could detect curcumin at wavelengths in the range of 350–370 nm.35-38 HPLC–diode array and fluorescence detection methods are used as another simple tool for detecting curcumin, with the most sensitive region in 390–450 nm. Some researchers have developed a high-performance thin-layer chromatography method, which is assisted with a phenyl column and acetonitrile/methanol/water as the solvent for curcumin detection and separation. 36 Microemulsion electrokinetic chromatography, employing oil droplets and surfactants, has proved useful in the extraction and estimation of curcumin. 43
Proliferation
In breast cancer cells, the survival signaling molecules, such as NF-κB, play a pivotal role in cell proliferation 44 ( Fig. 2 ). Liu et al. reported that curcumin was able to inhibit NF-κB expression 45 and toggled many downstream signaling pathways, which silenced inflammatory cytokines, such as CXCL1 and CXCL2, and mediated the expression of matrix metalloproteinase 9 (MMP-9), urokinase plasminogen activator (uPA), uPA receptor (uPAR), intercellular adhesion molecule 1 (ICAM-1), and chemokine receptor 4 (CXCR4). 46 Therefore, curcumin is believed to show its impact on cell growth and invasion of breast cancer partially through the downregulation of NF-κB signaling pathways. 47
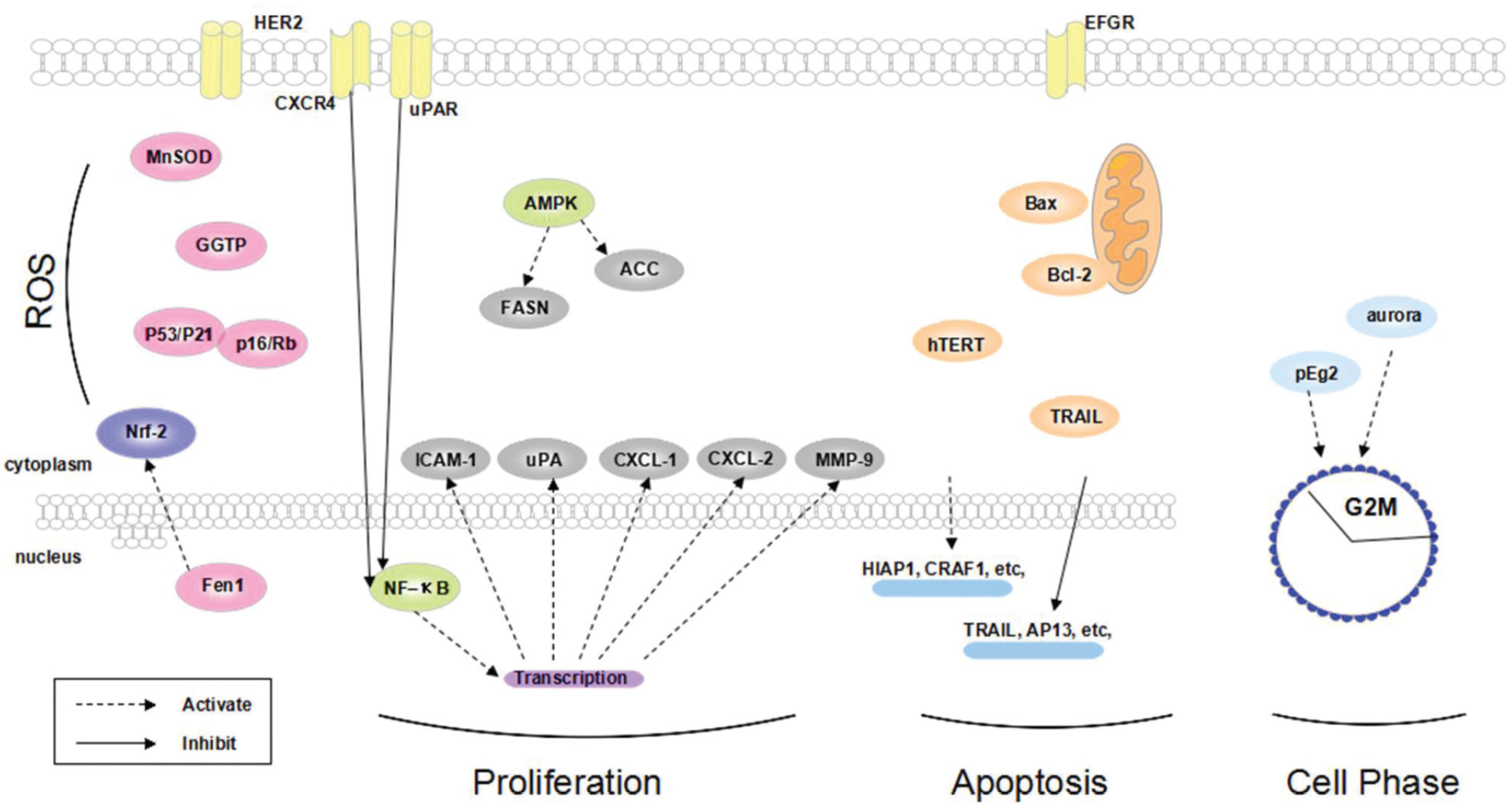
Molecular targets of curcumin on ROS, proliferation, apoptosis, and cell phase arrest. HER2, human epidermal growth factor receptor 2; MnSOD, manganese-dependent superoxide dismutase; GGTP, gamma-glutamyltranspeptidase; Nrf-2, nuclear factor 2–related factor; Fen1, Flap structure-specific endonuclease 1; CXCR4, chemokine receptor 4; uPAR, urokinase plasminogen activator receptor; FASN, fatty acid synthase; ACC, acetyl-CoA carboxylase; ICAM-1, intercellular adhesion molecule 1; uPA, urokinase plasminogen activator; MMP-9, matrix metalloproteinase 9; hTERT, telomerase reverse transcriptase; TRAIL, TNF-related apoptosis-inducing ligand.
Hormone factors also contribute to the growth and proliferation of breast cancer cells. HER2 and estrogen are the typical targets belonging to this category. Yim-im et al. found that a combination of curcumin and its analogues AS-KTC006 and AS-KTC021 could inhibit one specific tyrosine kinase and work as a rival against HER2. 48 Furthermore, curcumin can enhance the doxorubicin (a chemotherapeutic drug) cytotoxicity by decreasing redundancy HER2 in breast cancer cells. 49 On the other hand, immunoliposome encapsulation is believed to enhance curcumin selectivity in breast cancer cells that overexpress HER2. 50 Recent research proved that curcumin, acting as a phytoestrogen, competitively inhibited endogenous estrogen, which also contributed to the suppression of breast cancer cell growth. 51
Furthermore, curcumin suppresses breast cancer proliferation partially due to other trophic signaling pathways. Curcumin showed its influence on fatty acid synthase (FASN) and acetyl-CoA carboxylase (ACC) through AMPK activation. 52 Curcumin also partially inhibited breast cancer cell growth by downregulating the insulin-like growth factor 1 (IGF-1) axis. 53 Results from Thulasiraman et al. 54 demonstrated that curcumin mediated the suppression of the fatty acid–binding protein 5 (FABP5)/peroxisome proliferator-activated receptor β/δ (PPARβ/δ) pathway by making breast cancer cells more sensitive to retinotic acid. The same situation is applicable to leptin and its receptor. 55
Apoptosis
Curcumin is reported to induce breast cancer apoptosis by regulating the expression of apoptosis related genes. Lvet al. 56 applied microarray hybridization of lone-tech apoptotic arrays with labeled first-strand probes of total RNA to analyze and characterize the genes, which were regulated by curcumin in human breast cancer cells. In MCF-7 cells, genes HIAP1, CRAF1, GADD45, HPRT, MCL-1, BCL2L2, NIP1, TRAP3, GSTP1, PIG11, DAXX, PIG3, RBP2, and JNK1 were upregulated, while genes TRAIL, AP13, TNFR, SARP3, TRAIL-R2, TNFRSF5, TNFb, and hTRIP were downregulated.
Curcumin was also reported to regulate the apoptosis-related proteins. Exerting the antiapoptotic action by blocking pro-apoptotic counterparts, Bcl-2 inhibited the extrinsic apoptosis pathway. 57 Induced by curcumin, the antiapoptotic protein Bcl-2 increased, while the pro-apoptotic protein Bax decreased, leading to an elevated Bax/Bcl-2 ratio.58,59 Sun et al. 60 studied the curcumin treatment on triple-negative breast cancer (TNBC) and found that these cells significantly inhibited the phosphorylation levels of endothelial growth factor receptor (EGFR) and downstream signaling molecules, such as ERK1/2. Recent studies reported that curcumin enhanced the TNF-related apoptosis-inducing ligand (TRAIL)–induced apoptosis even in TRAIL-resistant breast cancer cells.61,62 Moreover, telomerase activity was inhibited by curcumin through downregulating the expression of hTERT (telomerase reverse transcriptase). 63
Cell Phase Arrest
Curcumin could also affect human breast cancer cells through inducing cell cycle arrest at the G2M phase and late S phase in MCF-7 cells. Curcumin led to an obvious increase of the G2M phase fraction. 64 Furthermore, Ke et al. 65 indicated that the percentage of cells arrested in the G2 phase and late S phase increased along with an increase of curcumin dose. Curcumin also acted similarly to Aurora-A SiRNA (small interfering RNA), leading to monopolar spindle formation, S and G2/M arrest, and cell division reduction in MCF-7 cells. 66
Curcumin induces cancer cell phase arrest through regulating spindle-related signaling pathways. A profound effect on mitotic spindle organization was exerted by curcumin directly, and the monopolar spindles were formed. Consequently, chromosomes were not able to be segregated normally and cells were arrested in the M phase. 67 Alternatively, curcumin also affected the expression levels of spindle motor protein indirectly, including the Xenopus laevis/Aurora/Ip11p-Related Kinase pEg2. Phosphorylation of Eg5 regulated the activity of the spindle motor protein, and downregulation of pEg2 resulted in monopolar spindles. 68
Modulation of MicroRNA
Curcumin suppresses breast cancer oncogenesis partially due to modulation of microRNA (miR), DNA, histone, and mitochrondria. The expression of miR-15a, miR-16, and miR-34a was upregulated in curcumin-treated cell models, and that of Bcl-2 and Bmi-1 was downregulated.69,70 Besides, recent evidence demonstrated that curcumin upregulated miR-22, which was conjectured to target estrogen receptor α (ERα) and the transcription factor Sp1, while miR-196 was dramatically downregulated. 71 Also, curcumin mediated the downexpression of two cytokines, CXCL-1 and CXCL-2, by steering miR181b. 72 Several papers revealed that DNA methyltransferase 1 (DNMT1), which epigenetically silenced multiple genes, decreased with curcumin treatment singly or in combination with other nature compounds.73,74 The same modulation happened upon acetylation of different histones, including H3K18, H4K16, and p53K120. 75
Immune and Metastasis Factors
Curcumin suppresses the growth of breast cancer partially by mediating the immune system. Recent research has indicated that curcumin could prevent the loss of T cells and inhibit immune suppressive cytokines, including transforming growth factor beta (TGF-β) and interleukin 10 (IL-10) in carcinogenesis.76,77 Particularly, tumor exosome-mediated inhibition of nature killer cell augmentation was partially reversed by curcumin through the impairment of a ubiquitin–proteasome system. 78 Moreover, curcumin acted as an inhibitor of the MEK/ERK signaling pathway, thus preventing tumor-dependent TGF-β-induced T–regulatory cell proliferation. 79
Curcumin also suppresses the growth of breast cancer by affecting metastasis factors in two ways ( Fig. 3 ). First, curcumin inhibited angiogenesis factors, such as vascular endothelial growth factor (VEGF) and basic fibroblast growth factor (bFGF), in ER-negative breast cancer cells.80,81 Meanwhile, Carroll et al. 82 reported that curcumin could suppress the secretion of angiogenesis factors, as one complication of medroxyprogesterone acetate. Second, curcumin also affects metastasis factors through invasion. Curcumin not only limited the expression of adhesion molecules (such as integrin α6β4) and the cytoskeleton protein (such as matrix metalloproteinase 9 and intercellular adhesion molecule 1), but also blocked recepteur d’origine nantais (RON) tyrosine kinase-mediated invasion.82-85 Moreover, it is certified that curcumin prevents hematogenous metastases in immunodeficient models. 86
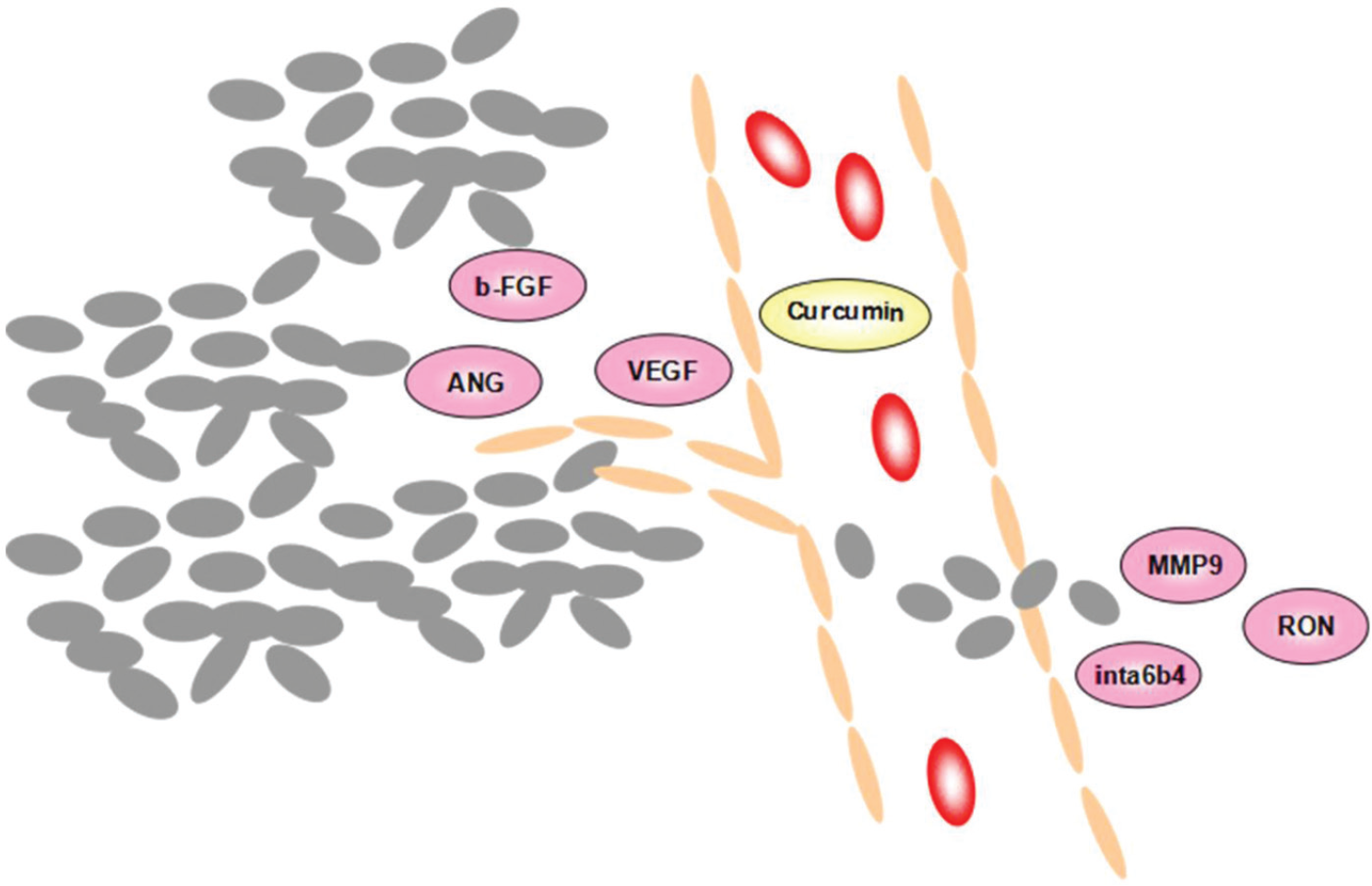
Molecular targets of curcumin on metastasis factors. VEGF, vascular endothelial growth factor; bFGF, basic fibroblast growth factor; ANG, angiopoietin; integrin α6β4, adhesion molecules; MMP-9, matrix metalloproteinase 9; RON, recepteur d’origine nantais.
Reactive Oxidative Stress
Reactive oxidative stress (ROS) expresses double-sided effects on carcinogenesis. On the one hand, ROS triggers cancer by DNA mutations. On the other hand, ROS has also been reported in some literature to kill tumor cells. Minimizing DNA damage resulting by ROS was always a vital consideration in the treatment of neoplasm. Curcumin has shown its effect on downregulating Flap endonuclease 1 (Fen1) expression, the overexpression of which was involved in the development of breast cancer, by interfering with the level of nuclear factor 2–related factor (Nrf-2), thus preventing the growth of breast cancer.87-89 Furthermore, curcumin could potentially increase the scavenger elements, such as manganese superoxide dismutase (MnSOD) and catalase. 90
Curcumin could also induce breast cancer cell apoptosis by promoting ROS. Curcumin-induced ROS accumulation leads to P53/p21- and p16/Rb-mediated breast cancer inhibition. 91 Moreover, Quiroga et al. detected a lower level of gamma-glutamyltranspeptidase (GGTP) activity even in an oxidation-resistant cell line treated with curcumin. 92
Oncogene and Tumor Suppressor Genes
In certain circumstances, oncogenes are activated and reprogram normal cells to cancer cells. Recently, Hong et al. 93 reported that the erbB2/neu (HER2) gene–encoded p185neu tyrosine kinase, the overexpression of which was assured to cause breast cancer, could be inhibited by curcumin. On the other hand, tumor suppressor genes prevent normal cells from being designated into cancer cells. Treatment of curcumin resulted in a high level of tumor suppressor genes, such as maspin and p16INK4A in invasive ductal carcinoma and cancer-associated myofibroblasts, respectively.93,94
Current Trend in Curcumin Pharmacy
In spite of the anticancer and antiantigenic properties of curcumin, its low potency and poor absorption limit the clinical application. 95 Analogues are designed and synthesized to improve the water solution.
Literature showed that analogues of curcumin have an anticarcinogenic effect on breast cancer due to mechanisms similar to those of curcumin. Hutzen et al. 96 compared the effect of GO-Y030 and curcumin, and their data showed that GO-Y030 had a more potent effect on inhibiting STAT3, especially in HER2-positive breast cancer cells. Another two analogues, FLLL11 and FLLL12, suppressed the growth of breast carcinoma cells by activating transcription 3 phosphorylation. Lin et al.’s results 97 showed that FLLL11 and FLLL12 were stronger inhibitors of STAT3 and AKT compared to curcumin. Moreover, analogues EF24 and EF31 blocked the nuclear translocation of NF-κB with a lower dosage, and PAC had a stronger effect on delaying the G2M phase.98,99 Furthermore, analogues had more cytotoxic effects through stimulating the cell phase 1 and phase 2 dox system, thus inducing stronger apoptosis of breast cancer cells. 100
Curcumin as Potent Chemosensitizer
Curcumin could serve as a chemosensitizer for other anticancer agents in treating human breast cancer cells, especially multi-drug-resistant (MDR) breast cancer cells. Limtrakul et al.101-104 investigated the effects of tetrahydrocurcumin on three ATP-binding cassette (ABC) drug transporters, including P-glycoprotein (ABCB1/P-gp), multidrug resistance protein 1 (ABCC1), and mitoxantrone resistance protein (ABCG2/MXR). The results illustrated that in a concentration-dependent manner, P-gp ATPase activity is stimulated and [(125)I]-iodoarylazidoprazosin (IAAP) is inhibited. Thus, the binding of MDR with IAAP leads to the efflux of mitoxantrone and the inhibition of pheophorbide. Curcumin also exerts early modification of relevant gene transcription, such as downregulation of Bcl-2 and inhibition of apoptosis proteins (IAPs) in MCF-7R cells with overexpression of P-gp. 105 In 5-FU-resistant cells, curcumin acts as a chemosensitizer for 5-FU by silencing the enzyme thymidylate synthase (TS) in breast cancer cells. 106 TS, an upstream molecule of NF-κB, can be inhibited by 5-FU. However, prolonged exposure of 5-FU reversely increases the expression of TS and activates TS-dependent NF-κB, which causes 5-FU resistance in breast cancer cells. Curcumin can silence TS and suppress NF-κB, and thus chemosensitize the 5-FU anticancer capability. A similar mechanism can be found in doxorubicin-resistant breast cancer cells. Meiyanto et al.’s study 107 suggested that curcumin inactivated NF-κB and reduced the expression of HER2 in doxorubicin-resistant breast cancer cells.
Footnotes
Declaration of Conflicting Interests
The authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The authors disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This paper has been supported by the National Natural Science Foundation of China (81301293) and National Science and Technology Major Projects for “Major New Drugs Innovation and Development” (2014ZX09507008).