
Abstract
Background
Wogonin is commonly used for the treatment of allergic diseases. However, neither its precise effect in preventing allergic rhinitis (AR) nor its mechanism of action are known.
Objectives
In this study, the effect of wogonin on allergic responses in ovalbumin (OVA) induced AR was investigated in mice.
Methods
BALB/c mice were sensitized with intraperitoneal (i.p.) OVA and then challenged intranasally with OVA. Wogonin (10 and 30 mg/kg) was given to the treatment groups, and the effect of wogonin on the release of allergic inflammatory mediators, specifically OVA-specific immunoglobulin E (IgE) and inflammatory cytokines, was explored. Eosinophil infiltration and the levels of interleukin (IL) 5 and IL-13 were measured by immunohistochemistry.
Results
In mice with AR, wogonin decreased OVA-specific IgE levels in serum, and the levels of the cytokines IL-4, IL-5, IL-13, eotaxin, and RANTES in nasal lavage fluid. Serum levels of IL-4, IL-5, and IL-13 were lower in both groups of wogonin-pretreated mice than in the OVA group. A reduction in eosinophil infiltration of the nasal mucosa and inhibition of the expression of IL-5 and IL-13 were also noted in the treated groups.
Conclusion
Wogonin induced antiallergic effects in a murine model of AR by decreasing the infiltration of eosinophils and levels of T-helper type 2 cytokines. Thus, wogonin merits consideration as a therapeutic agent for treating AR.
Allergic rhinitis (AR) is a disease with a prevalence in the general population worldwide of 3–19%, but this is increasing in frequency.1,2 The prevalence of AR in Korea ranges from 6 to 10%, and this is also increasing.3,4 The typical symptoms of AR are nasal congestion, rhinorrhea, nasal itching, conjunctivitis, and disturbed olfaction, all of which create difficulties in school or work life, and are a major risk factor for asthma exacerbation.5–7 AR is caused by allergy mediators such as histamine, inflammatory cytokines, eosinophils, and mast cells. 8 The AR reaction begins with the binding of mast cells to immunoglobulin E (IgE) receptors on the surface of basophils, which induces mast cell secretion of inflammatory cytokines and histamine. 9 As a result, inflammatory cells and eosinophils migrate to the involved tissues. 10
Current pharmacotherapy for AR includes corticosteroids, leukotriene receptor antagonists, mast cell stabilizers, and anticholinergics, but many patients seek complementary and alternative treatments, including the use of medicinal herbs, to decrease allergic inflammation.11,12 However, the precise effects and mechanisms of action of these approaches in reducing allergic asthma are unclear. Wogonin (5,7-dihydroxy-8-methoxyflavone) is a component of the dried root of Scutellaria baicalensis Georgi that has been used in China to treat liver disease, leukemia, and atherosclerosis.13,14 Antiangiogenesis, anticancer, and anti-inflammatory effects have also been described.15–17 In traditional Chinese medicine, wogonin is also used to treat allergies and inflammation, based on its toxin-removing and thermal cleansing properties, 18 Antioxidative effects have also been described. 19 In the present study, we examined the ability of wogonin to inhibit ovalbumin (OVA) induced allergic nasal symptoms and inflammatory cytokine release in a murine model of AR. We also investigated the mechanisms that underlie its effects.
METHODS
Animals
Six-week-old female BALB/c mice were purchased from Orient Bio Inc. (Seongnam, Korea). All animal experiments were carried out according to the Gachon University Animal Research Guidelines. The study was approved by Gachon University.
Group Classification and Development of an AR Model
The 6-week-old female BALB/c mice were divided into five groups (n=7 per group): (1) the normal control group, (2) OVA-sensitized group, (3) wogonin 10 mg/kg group, (4) wogonin 30 mg/kg group, and (5) dexamethasone 4 mg/kg group. After the mice had been allowed to stabilize for 1 week, they were sensitized to OVA on days 0, 7, and 14 by an intraperitoneal (i.p.) injection of 50 mg of OVA (chicken egg albumin; Sigma-Aldrich Corp., St. Louis, MO) in 100 μL of phosphate-buffered saline solution that contained 2 mg of aluminum hydroxide (alum; Sigma-Aldrich). One week after the last injection, on day 21, the mice were challenged with 20 μL of phosphate-buffered saline solution that contained 50 μg/mL of OVA into the bilateral nasal cavities. From day 21 to day 31, the mice in the control and OVA groups were perorally administered saline solution, whereas those in the dexamethasone group received 4 mg/kg of dexamethasone (Sigma-Aldrich), and the wogonin groups were each orally administered 10 and 30 mg/kg of wogonin 1 hour before intranasal challenge of OVA under the same conditions. Wogonin was purchased from Alexis Biochemicals (San Diego, CA) and dissolved in dimethylsulfoxide. Dexamethasone is a steroid medication used in the treatment of skin diseases, severe allergies, and asthma. 20 In this study, dexamethasone served as a positive control.
Collection of Serum and Nasal Lavage Fluid
Twenty-four hours after the last stimulation, the mice were euthanized while they were under anesthesia with isoflurane. Orbital puncture was used to obtain 300–400 μL of blood from the ophthalmic artery of each mouse. The blood samples were stored at 4°C for 2–4 hours and centrifuged, and the serum was collected. Nasal lavage fluid (NALF) was also collected in the posterior to anterior direction after irrigation with 1 mL of saline solution through an angiocatheter placed in the trachea. The lavage fluid was centrifuged (2000 rpm at 4°C), and 0.5 mL of the supernatant was stored at −70°C.
Extraction of Nasal Tissue Specimens
The decapitated head of each mouse was fixed in 10% paraformaldehyde for 24 hours. The extruded nasal tissue specimen was decalcified for 5 days and then sectioned from behind the upper incisor to 2 mm in front of the incisive papilla of the hard palate. After a paraffin block was prepared, slices of 4-μm thickness were obtained.
Measurement of OVA-Specific IgE in Serum
OVA-specific IgE levels were measured by using a mouse anti-OVA IgE enzyme-linked immunosorbent assay kit (Alpha Diagnostic International Inc., San Antonio, TX).
Measurement of Cytokine Levels in NALF and Serum
The levels of interleukin (IL) 4, IL-5, IL-13, eotaxin, and RANTES in NALF and serum were measured by using the respective enzyme-linked immunosorbent assay kits (BioSource International Inc., Camarillo, CA).
Hematoxylin and Eosin Staining
After deparaffinization of the samples, the nasal tissue specimens were stained with hematoxylin and eosin, and the numbers of inflammatory cells at five different sites were counted under ×400 magnification.
Immunohistochemical Staining
Nasal specimens were incubated with mouse monoclonal antihuman IL-4 antibody (BioVendor Laboratory Medicine Inc., Brno, Czech Republic) and purified antimouse/human IL-5 antibody (BioLegend, San Diego, CA), which was diluted 1:200, for 90 minutes at room temperature. The specimens were then incubated with goat-antimouse immunoglobulin (Santa Cruz Biotechnology, Santa Cruz, CA) for 1 hour at room temperature. Immunoreactive proteins were visualized by incubation with streptavidin-conjugated peroxidase for 15 minutes. After decolorization by using a 3,3-diaminobenzidine tetrahydrochloride solution, the samples were counterstained with hematoxylin. Control plates were processed similarly, but the primary antibody was omitted. The average ratio of the immunopositive area to the total area of epithelial cells in the nasal specimen was determined microscopically under ×100 magnification.
Statistical Analysis
The pathologic results regarding the nasal septum and cytokine levels in the NALF were analyzed statistically by using the Student’s t-test. A p value of <0.05 was considered to indicate statistical significance.
RESULTS
Effect of Wogonin on OVA-Specific IgE Levels in Serum of the AR Model
The mouse OVA-specific IgE level was 515.4±88.5 U/mL in the OVA group, which was significantly higher than that of the normal control group (50.1±U/mL). By contrast, mice administered either wogonin 30 mg/mL or dexamethasone had significantly lower OVA-specific IgE levels than the OVA group: 299.7±60.0 U/mL and 274.6±45.6 U/mL, respectively (Fig. 1).
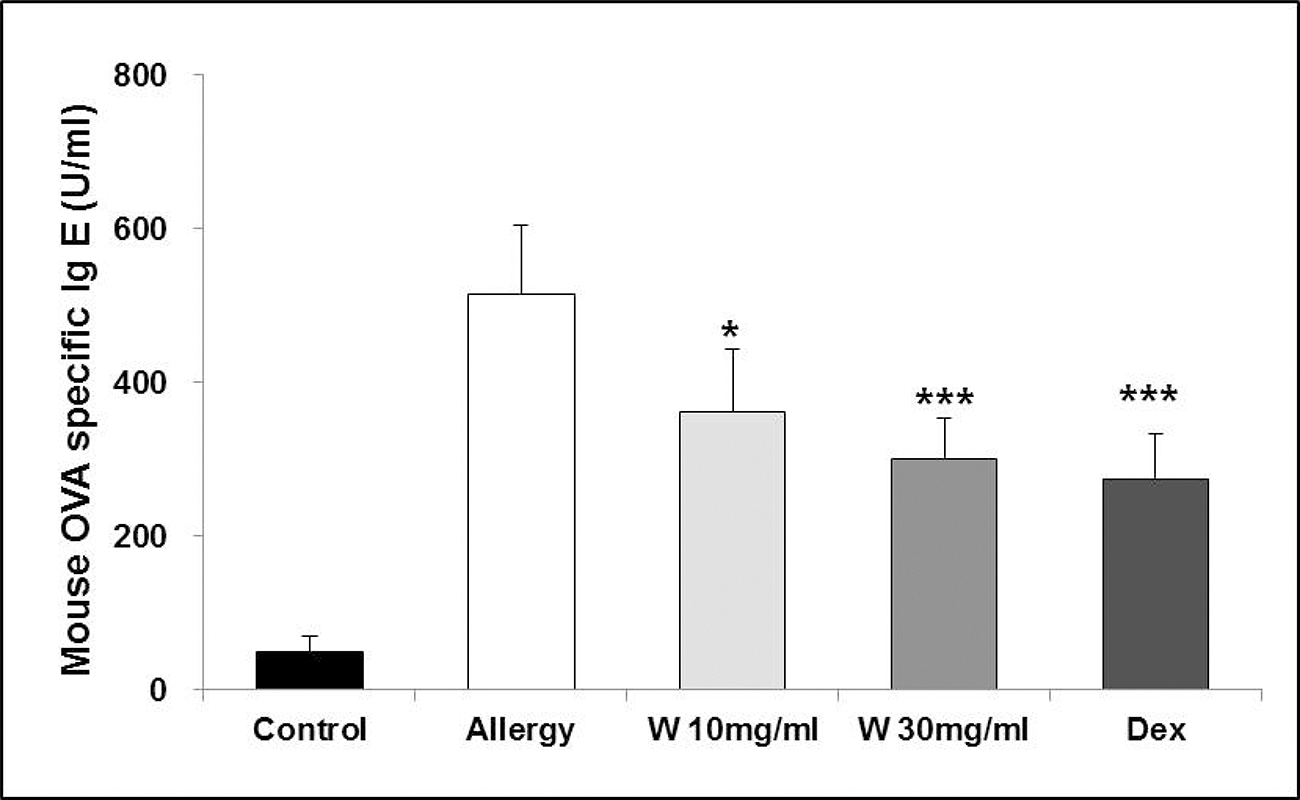
Effect of wogonin on OVA-specific IgE levels in serum of the allergic rhinitis (AR) mouse model. OVA-specific IgE levels were measured by using an enzyme-linked immunosorbent assay. Mice in the Dex and wogonin groups were treated orally with dexamethasone: dexamethasone, 4 mg/kg; and wogonin, 10 mg/mL; and wogonin 30 mg/mL. Control and OVA mice were treated with distilled water. Independent experiments were performed (n=7). The columns and error bars represent the mean±standard error, respectively. *p < 0.05 and ***p < 0.005, significantly different from the OVA group. IgE=immunoglobin E; OVA=ovalbumin; W=wogonin; Dex=dexamethasone.
Infiltration of Eosinophils in Nasal Mucosa
The number of infiltrated eosinophils was significantly higher in the OVA group than in the normal control group. In the wogonin and dexamethasone groups, eosinophil counts were significantly lower than in the OVA group (Fig. 2). These results indicated that treatment with wogonin effectively inhibited the infiltration of eosinophils.

Effects of wogonin on the filtration of eosinophils (arrows) into nasal mucosa of mice with allergic rhinitis (AR). (A) Eosinophil infiltration of the nasal mucosa. Representative hematoxylin-and-eosin–stained sections of the nasal mucosa tissues are shown at a magnification of ×400. The infiltration of inflammatory cells was markedly reduced in mice in the wogonin-treated groups. (B) The number of eosinophils was significantly lower in the wogonin groups than in the OVA group.
Effect of Wogonin on Serum Cytokine Levels in the AR Model
Because allergic diseases are caused by the secretion of a series of T-helper (Th) type 2 cytokines, particularly IL-4, IL-5, and IL-13, we examined whether their levels were changed in OVA+ wogonin-treated mice. The serum concentrations of IL-4, IL-5, and IL-13 were higher in the OVA group than in the control group (Fig. 3). Whereas the difference in serum IL-4 and IL-5 levels did not significantly differ between the wogonin and dexamethasone groups, IL-13 levels in the wogonin groups were significantly lower than in the OVA group.

Effects of wogonin on the serum levels of cytokines in a mouse model of allergic rhinitis (AR). The concentrations of (A) interleukin (IL) 4, (B) IL-5, and (C) IL-13 in the sera of the mice with AR were measured by using mouse cytokine enzyme-linked immunosorbent assay kits. Independent experiments were performed (n=7). The columns and error bars represent the mean±SE, respectively. *p < 0.05 and ***p < 0.005, significantly different from the OVA group.
Effect of Wogonin on Cytokine Levels of NALF in the AR Model
Next, we examined cytokine levels in the NALF of OVA+ wogonin-treated mice. IL-4 and IL-5 levels in NALF were not significantly changed in the ovalbumin induced group compared with the normal control group (Fig. 4, A and B). However, the difference in IL-4 and IL-5 levels in the groups administered wogonin differed significantly from those of normal control mice. As shown in Fig. 4 C , IL-13 levels were elevated in mice with allergy (112.8±14.5 pg/mL) compared with normal control mice (68.4±7.3 pg/mL) but significantly lower in mice treated with wogonin 10 mg/mL (67.3±7.0 pg/mL), wogonin 30 mg/mL (67.5±2.5 pg/mL), or dexamethasone (58.4±1.3 pg/mL). Eotaxin levels were higher in mice with allergy (80.0±41.1 pg/mL) than in normal control mice (29.3±2.9 pg/mL). By contrast, these levels were significantly lower in mice treated with wogonin at 10 and at 30 mg/mL (45.8±12.3 pg/mL and 59.4±2.7 pg/mL, respectively), and dexamethasone (45.1±14.6 pg/mL) than in mice with allergy (p <0.05) (Fig. 4 D). The level of RANTES was also significantly reduced in mice treated with either the lower or higher dose of wogonin (32.3±0.9 pg/mL and 31.2±0.2 pg/mL, respectively) or with dexamethasone (30.0±0.2 pg/mL) than in mice with allergy (p <0.05) (Fig. 4 E).

Effects of wogonin on cytokine levels in the nasal lavage fluid (NALF) of mice with allergic rhinitis (AR). The concentrations of (A) interleukin (IL) 4, (B) IL-5, (C) IL-13, (D) eotaxin, and (E) RANTES in the NALF of mice with AR were measured by using mouse cytokine enzyme-linked immunosorbent assay kits. Independent experiments were performed (n=7). The columns and error bars represent the mean±SE. *p < 0.05 and **p < 0.005, significantly different from the ovalbumin (OVA) group.
Immunohistochemistry of the Nasal Mucosa
IL-5 and IL-13 expression in the OVA-induced AR mouse model was measured by using immunohisto-chemistry. IL-5 expression in the nasal septum of the wogonin group was significantly lower than in the OVA group (Fig. 5). The decreased expression of IL-13 was relatively weak.
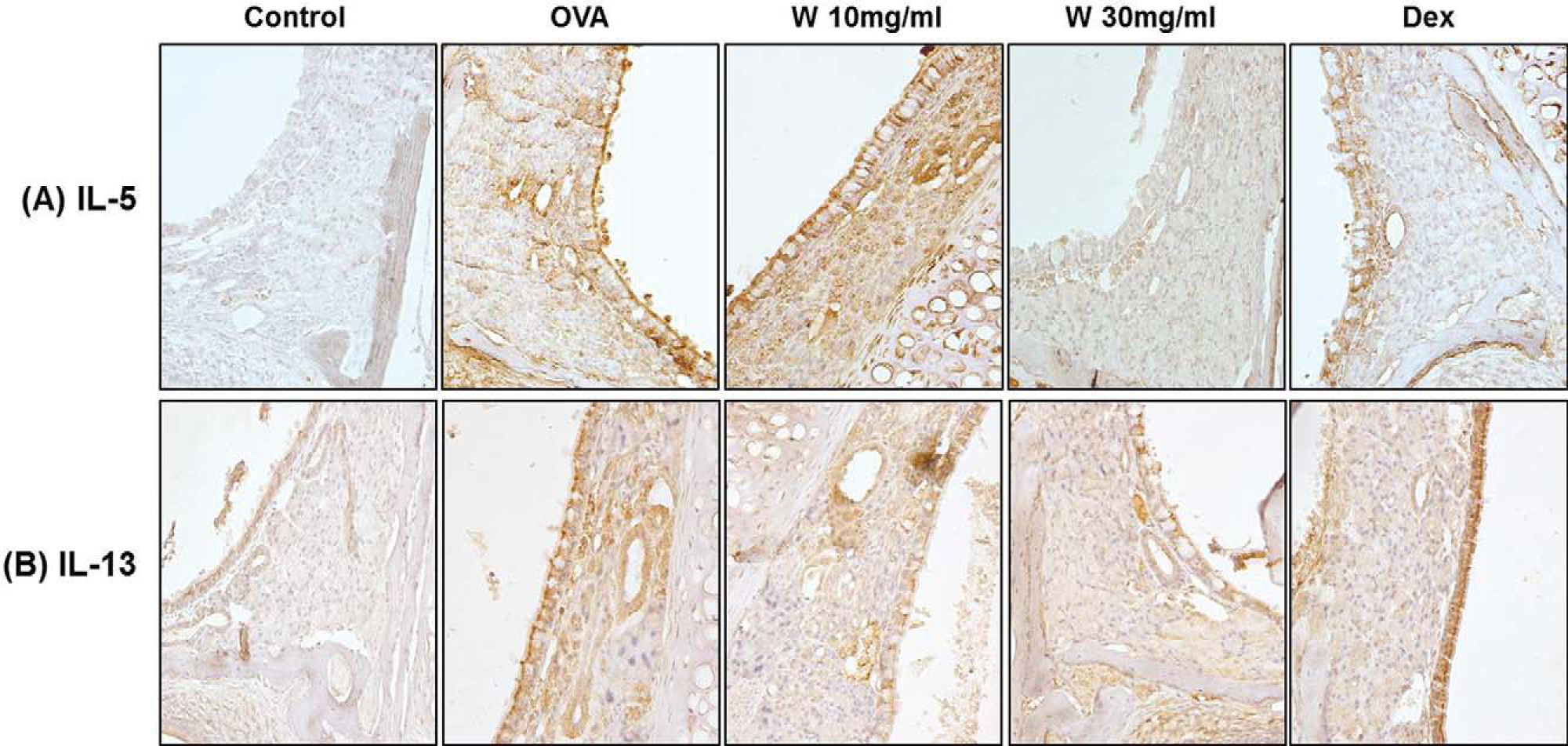
Effect of wogonin on interleukin (IL) 5 and IL-13 expression in the nasal mucosa of mice with allergic rhinitis (AR). Nasal mucosa tissues were processed for immunohistochemical staining for (A) IL-5 and (B) IL-13. The photographs are representative images taken at a magnification of ×400.
DISCUSSION
In this study, we showed that wogonin inhibited OVA-induced inflammation in a murine model of AR. Specifically, wogonin inhibited the levels of Th2-type inflammatory cytokines and reduced the number of eosinophils in the nasal mucosa. It also decreased OVA-specific IgE, IL-4, IL-5, and IL-13 levels in NALF as well as OVA-induced eotaxin and RANTES levels. AR is an inflammatory disease of the nasal mucosa characterized by the symptoms of nasal obstruction, sneezing, and rhinorrhea.21,22 OVA sensitization and challenge in animal models lead to an increase in the OVA-specific IgE and infiltration of inflammatory cells in the epithelium of the nasal mucosa. 23
The symptoms of AR are the product of an IgE-mediated inflammation of the nasal mucosa, which involves inflammatory cells. Eosinophils are the major inflammatory cell type that contributes to AR. They release proinflammatory mediators, such as Leukotriene C 4 and platelet activating factor, which increase the permeability of mucosal cells and are potent chemotactic factors that stimulate further eosinophil infiltration. Eosinophils also produce large amounts of proteins that are toxic to respiratory epithelial cells. 24 Th2 cells release cytokines, including IL-4, IL-5, and IL-13, all of which play important roles in allergic diseases. 25 IL-4 and IL-13 promote IgE production by B cells and mucus production in the airways. IL-4 also induces the activation of T cells and their differentiation into the Th2 subtype.26,27 IL-5 plays a critical role in the generation and release of eosinophils in the bone marrow, 28 whereas eotaxin promotes the selective recruitment of eosinophils into the inflamed area. 29 As shown in Fig. 4, A–C, wogonin reduced Th2 cytokine production (IL-4, IL-5, and IL-13) in NALF and thus demonstrated antiallergic effects.
Wogonin is one of several compounds from S. baicalensis with a proven anti-inflammatory effect. 30 A recent study of a murine model of allergic asthma showed that the potent activity of wogonin involves the regulation of the IL-4/ signal transducer and activator of transcription 6 signaling pathway. 31 In this study, wogonin treatment inhibited Th2 responses, thereby reducing IL-4 and IL-5, and attenuated allergic inflammation. Airway epithelial cells release high levels of eotaxin when stimulated with IL-4 or IL-13.32,33 IgE produced by B cells stimulated with IL-4 and IL-13 binds to mast cell receptors. The subsequently activated mast cells produce a variety of mediators and cytokines 34 that contribute to the characteristic allergic responses, 35 including the release of chemokines, cytokines, and growth factors by epithelial cells. This response promotes persistent late-stage allergic reactions and progressive allergic inflammation.36,37 Our results, obtained in a murine model of AR, indicated that the reduction of allergic inflammation by wogonin was related to a decrease in Th2 cytokine production, tissue eosinophilia, and production of OVA-specific IgE. These findings indicated the potential anti-inflammatory role of wogonin as an inhibitor of OVA-specific allergic responses in the nasal mucosa.
CONCLUSION
Wogonin suppressed allergic cytokines in a murine model of AR. Its anti-inflammatory activity is mediated by a decrease in eosinophil migration into the nasal mucosa and inhibition of serum IgE and of nasal IL-4, IL-5, and IL-13 levels. Our study demonstrated the antiallergic effect of wogonin in an in vivo model of AR, and, thus, by extension, the potential therapeutic utility of wogonin in treating AR in humans.
ETHICAL APPROVAL
This study was approved by the Institutional Review Board of Gachon University Gil Medical Center (approval number: GCIRB2014-362).
STATEMENT OF HUMAN AND ANIMAL RIGHTS
This article does not contain any studies with human or animal subjects.
STATEMENT OF INFORMED CONSENT
There are no human subjects in this article and informed consent is not applicable.