
Abstract
Cellular immunity plays important roles in clearing intracellular pathogens or tumor cells. It is of significance to develop an adjuvant able to efficaciously induce cellular immunity, but such an approved adjuvant is not currently available. Klebsiella pneumoniae (K. pneumoniae) possesses lipopolysaccharide and capsular polysaccharide, both of which have been demonstrated to be able to induce humoral immunity. In this study, we investigated the effect of K. pneumoniae on epidermal cellular immunity, in vitro. The effects of K. pneumoniae on the maturation of Langerhans cells (LCs), the capacity to induce T-cell proliferation, and the secretion of interferon gamma (IFN-γ) by T cells were examined. The results showed that K. pneumoniae induced significant upregulation of CD86 and human leukocyte antigen-deterodimer (HLA-DR) levels on LCs, but not CD40 and CD80. K. pneumoniae-loaded LCs induced significant CD4+ and CD8+ T-cell proliferation. Significant increases in extracellular IFN-γ secretion by CD4+ and CD8+ T cells stimulated with K. pneumoniae-pulsed LCs using enzyme-linked immunospot assay (ELISPOT) were demonstrated, while only a part of the subjects showed increases in intracellular secretion of IFN-γ using intracellular staining assay. In sum, K. pneumoniae has the potential to enhance epidermal cellular immunity and may act as a potential adjuvant in intradermal vaccines designed to enhance cellular immunity.
Keywords
Introduction
Cellular immunity plays important roles in clearing intracellular pathogens or tumor cells. 1 The adjuvants in licensed vaccines include aluminum salt, oil in water emulsion adjuvant such as MF59 and AS03, which mainly help to increase humoral immunity, but not cellular immunity. Although the compound adjuvant AS04 is able to induce cellular immunity, the homogeneity of vaccine preparation and high cost limit the wide usage of AS04. This urged us to develop new economic and easy to be obtained adjuvants able to enhance both humoral immunity and cellular immunity. Microbe-derived adjuvants acting like pathogen-associated molecular patterns (PAMPs) can trigger innate immune responses, activate antigen presenting cells (APCs), and initiate adaptive immune responses. Klebsiella pneumoniae (K. pneumoniae) is a member of the normal flora on the mucosal surfaces of mammals. 2 K. pneumoniae possesses surface saccharides in the form of lipopolysaccharide (LPS) and capsular polysaccharide (CPS), both of which have been demonstrated to be able to induce humoral immunity. 3 It is unclear whether K. pneumoniae lysate or the components from K. pneumoniae affect cellular immunity. In this study, we found that K. pneumoniae lysate was capable of enhancing cellular immunity. It is conceivable that K. pneumoniae lysate represents a promising ideal adjuvant for vaccines, because its components have been demonstrated to be capable of inducing humoral immunity as well because it is accessible and economic.
Materials and methods
Generation of monocytes-derived Langerhans cells
Fresh blood samples from healthy donors (n = 6) were obtained from Liaoning Province Blood Center, China. The subjects were numbered in chronological order. Peripheral blood mononuclear cells (PBMCs) were isolated and purified using ficoll lymphocyte separation medium (Axis-Shield-Lymphoprep, Norway). CD14+ monocytes were isolated from human PBMCs using CD14+ MicroBeads (Miltenyi Biotec, Auburn, USA) and the cells (1 × 106/mL) were cultured in RPMI-1640 (Hyclone, USA) medium containing 10% fetal calf serum (Clark, Australia), 1% penicillin/streptomycin (Genview, USA), rhGM-CSF (PeproTech, USA; 1000 IU/mL), rhIL-4 (PeproTech; 1000 IU/mL), and TGF-β1 (PeproTech; 10 ng/mL) to induce Langerhans cells (LCs) as described by Fahey et al. 4 Cytokines were replenished at equal concentration on the third day. After 7 days, LCs were successfully converted and the successful conversion to LCs was demonstrated by detecting CD1a (Miltenyi Biotec, Auburn, USA), Langerin (Miltenyi Biotec) using FACS Fortessa (China Medical University, The First Experimental Technology Center). The data were analyzed using Flowjo software (BD Biosciences, USA). The study was approved by the Institutional Review Board of the China Medical University, and written informed consents were obtained.
Maturation analysis of LCs treated with K. pneumoniae
K. pneumoniae standard strain 13883 was kept in our laboratory. K. pneumoniae (10 μL) were cultured in 1 mL Luria-Bertani medium in a shaking incubator (185 r/min) at 37°C for 18 h. Then 50 μL logarithmic phase bacteria were cultured in 5 mL Luria-Bertani medium in a shaking incubator (185 r/min) at 37°C for 2 h. K. pneumoniae were pelleted by centrifugation (2000 g for 15 min at 4°C), washed twice with phosphate buffered saline (PBS), and resuspended in 5 mL PBS. K. pneumoniae lysate were obtained by breaking the bacteria for 15 min using ultrasonic cell disrupter (SM-900D, China) at 4°C and then heating them for 30 min at 95°C in a water bath. K. pneumoniae lysate were used to stimulate LCs at a ratio of 100:1 (bacteria/LCs). Tetanus toxoid (500 ng/mL; Calbiochem, USA) was used as positive control and media was used as negative control. After 48 h incubation, LCs were harvested and stained with antihuman CD40-phycoerythrin (PE; Miltenyi Biotec), CD80-allophycocyanin (APC; Miltenyi Biotec), CD86-fluorescein isothiocyanate (FITC; Miltenyi Biotec), and and human leukocyte antigen-deterodimer (HLA-DR)-peridinin chlorophyll protein (Percp; Miltenyi Biotec). The data were analyzed using Flowjo software (BD Biosciences, USA).
Analysis of T-cell proliferation induced by LCs treated with K. pneumoniae
On the ninth day of LCs conversion, CD4+ and CD8+ T cells were isolated from CD14− cells from the same subjects using CD4+ T Cell Isolation Kit II (Miltenyi Biotec) and CD8+ T Cell Isolation Kit II (Miltenyi Biotec) according to the manufacturer’s instructions. The effects of T-cell proliferation stimulated by LCs treated with K. pneumoniae were assessed using MTS assay. The assay was performed in six replicate wells by co-culturing T cells (1.5 × 106 cells/mL) with autologous LCs or K. pneumoniae-loaded LCs (4 × 104 cells/mL) in 100 μL of RPMI-1640 media (Hyclone, USA) in each well of a 96-well plate. Wells containing cells (T cells and LCs) and K. pneumoniae (10 μL), cells (T cells and K. pneumoniae-treated LCs) and K. pneumoniae (10 μL), cells (T cells and LCs) and RPMI-1640 media, and cells (T cells and LCs) and phytohemagglutinin (PHA; 2.5 μg/mL; DAKEWE, China) were set up. After incubation at 37°C for 7 days, cells were incubated for 4 h with 40 μL/well of CellTiter 96 Aqueous One solution (Promega, USA). The absorbency (ABS) at 490 nm was recorded with a Varioskan Flash Multimode Reader (Thermo Scientific, USA).
Extracellular interferon-γ secretion by T cells stimulated with K. pneumoniae-treated LCs using enzyme-linked immunospot assay
On the ninth day of LC conversion, CD4+ and CD8+ T cells were isolated from CD14− cells as described above. Enzyme-linked immunospot assay (ELISPOT) was performed in duplicate wells by co-culturing T cells (1.5 × 106 cells/mL) and autologous LCs or K. pneumoniae-treated LCs (4 × 104 cells/mL) in 100 μL of serum-free media (DAKEWE, China). Wells containing cells (T cells and K. pneumoniae-treated LCs) and K. pneumoniae (10 μL), cells (T cells and LCs) and K. pneumoniae (10 μL), cells (T cells and LCs) and media, and cells (T cells and LCs) and PHA (DAKEWE, China; 2.5 μg/mL) were set up. Clearly defined steps accorded to the human interferon gamma (IFN-γ) precoated ELISPOT kit’s instructions. After drying overnight at room temperature, spot forming units (SFUs) were counted using an automated image analysis system Bioreader 4000 (BioSys, Germany).
Intracellular IFN-γ secretion by T cells stimulated with K. pneumoniae-treated LCs using intracellular staining assay
On the ninth day of LCs conversion, CD4+ and CD8+ T cells were isolated from CD14− cells from the same subjects as described above. They were co-cultured with autologous K. pneumoniae-treated LCs at a ratio of 50:1 (T cells: LCs) in 1 mL of RPMI-1640 media in each well of a 12-well plate. K. pneumoniae (100 μL) and tetanus toxoid (500 ng/mL) as a positive control were added to stimulate cells. RPMI-1640 media alone was used as a negative control. After 6 days of incubation, phorbol 12-myristate 13-acetate (PMA, 200 nM, Sigma) and ionomycin (1 μg/mL, Biolegend, USA) were added for 4 h and Brefeldin A (10 μg/mL, Biolegend) was added for an additional 2 h. Cells were permeabilized/fixed and stained with antihuman IFN-γ APC and relevant isotype control. A total of 10,000 events were acquired using FACS Fortessa. The percentages of IFN-γ of T cells were analyzed and calculated using Flowjo software.
Statistical analysis
The statistical significance of differences between the experimental groups and control groups was determined using one-way analysis of variance (ANOVA). Significance was accepted at the P < 0.05 levels.
Results
Phenotypic maturation analysis of LCs
Successful conversion to LCs was demonstrated by the expression of CD1a, Langerin on LCs (data not shown). The mature effects of K. pneumoniae on LCs are shown in Figure 1. For CD86 and HLA-DR, mean fluorescence intensity (MFI) of LCs treated with K. pneumoniae was significantly higher than that of LCs treated with media (P < 0.05). Although the expression of CD40 on LCs in four samples and CD80 in all the examined five samples increased, overall no significant increases in the expression of CD40 and CD80 were observed in K. pneumoniae-treated LCs compared to untreated LCs after statistical analysis. It is unexpected that no significant increases were observed with LCs treated with tetanus toxoid over media for all these four surface molecules.
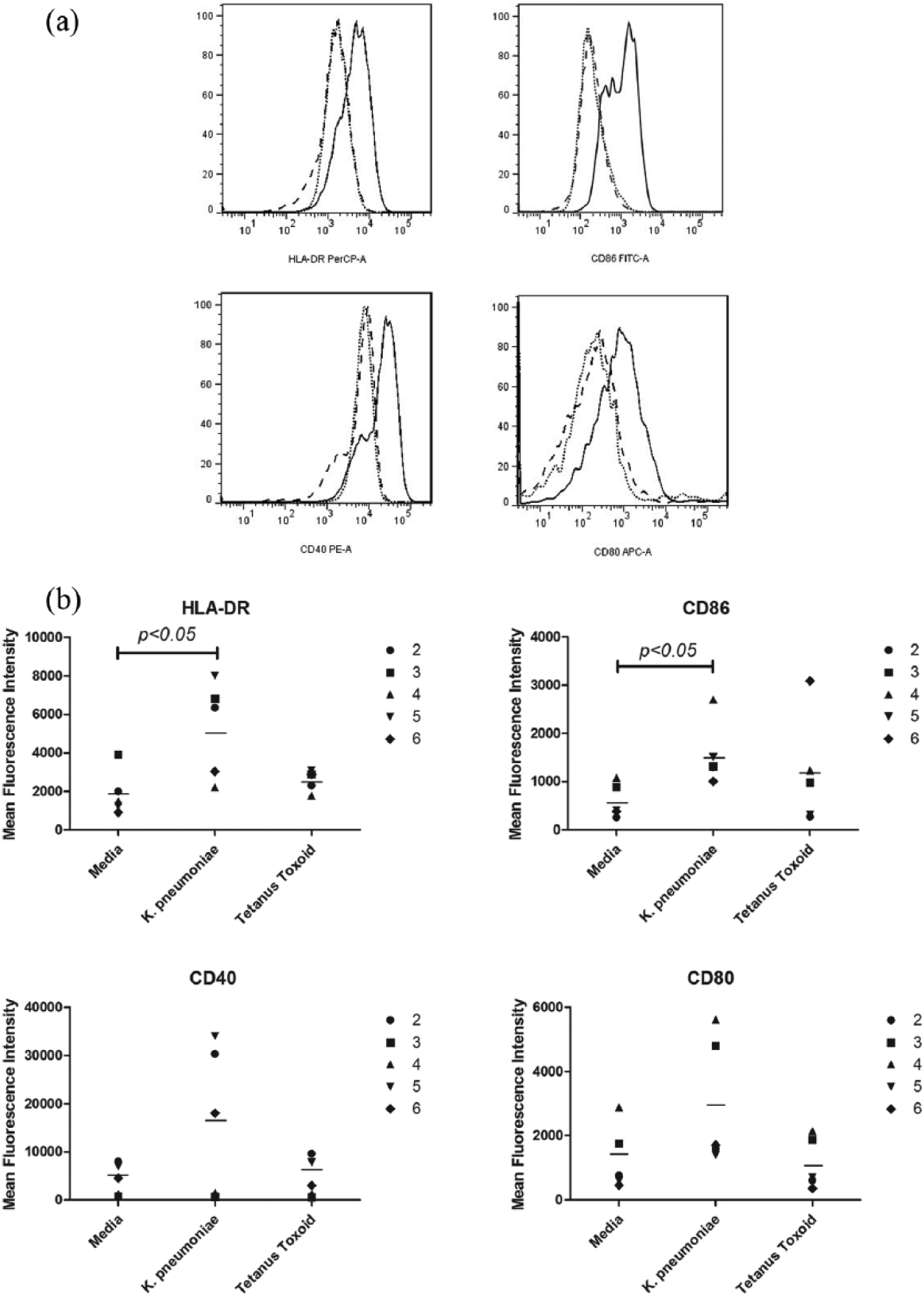
Maturation effects of K. pneumoniae on LCs detected by surface expression of HLA-DR, CD86, CD40, and CD80. (a) Representative FACS histograms from subject 2. The black dotted line, dashed line, and black solid line represent the media, tetanus toxoid, and K. pneumonia, respectively. (b) Summary of results from all subjects examined except subject 1. The bar represents the mean of the groups.
T-cell proliferation measured by MTS
The proliferation of CD4+ T cells treated with LCs or K. pneumoniae pre-pulsed LCs in the presence of K. pneumoniae increased significantly over media. For CD8+ T cells, absorbency of T cells treated with LCs or K. pneumoniae pre-pulsed LCs also increased significantly than media. PHA did not induce a significant increase in CD4+ and CD8+ T-cell proliferation though the increased T-cell proliferation in some subjects was observed (Figure 2.). In sum, K. pneumoniae-treated LC could induce T-cell proliferation effectively.

CD4+ and CD8+ T-cell proliferation measured using MTS. Summary of results from all subjects examined. The bar represents the mean ABS of the groups.
Extracellular IFN-γ secretion by ELISPOT
For CD4+ and CD8+ T cells, SFUs formed by T cells treated with LCs or K. pneumoniae pre-pulsed LCs in the presence of K. pneumoniae increased significantly higher than media. PHA also induced a significant increase of SFU (Figure 3). Since each spot represents a specific IFN-γ secreting T lymphocyte in ELISPOT, we can say K. pneumoniae can stimulate the IFN-γ secretion of CD4+ and CD8+ T cells effectively.
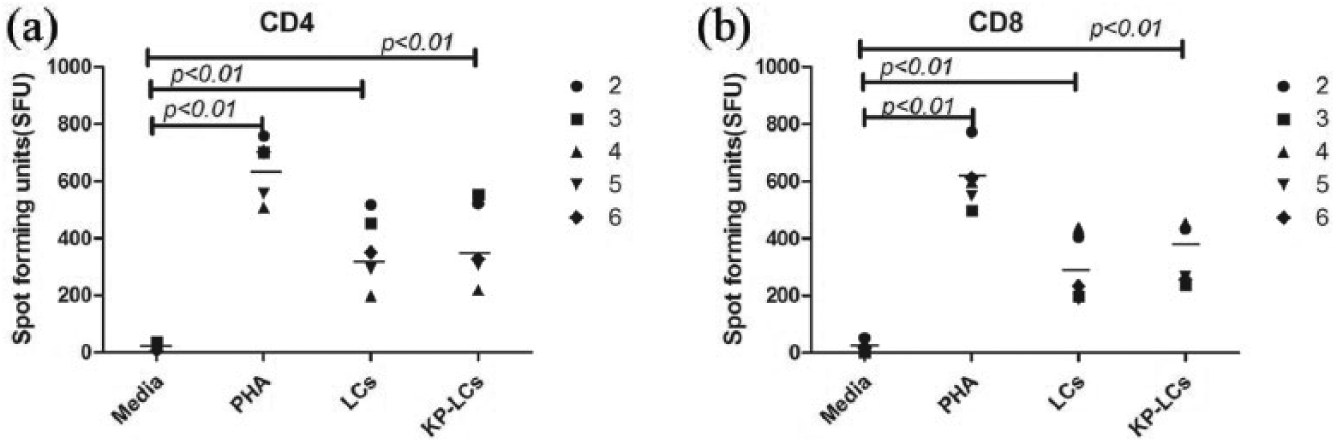
IFN-γ secretion by CD4+ and CD8+ T cells stimulated with K. pneumoniae using ELISPOT. Summary of results from all subjects examined except for subject 1. The bar represents the mean of the group.
Intracellular secretion of IFN-γ by T cells stimulated with K. pneumoniae-pulsed LCs
CD4+ and CD8+ T cells stimulated with K. pneumoniae-pulsed LCs from six subjects were stained for intracellular secretion of IFN-γ. Intracellular secretion of IFN-γ by CD4+ and CD8+ T cells was not significantly increased, though increased IFN-γ secretion (>5%) was observed in CD4+ T cells exposed to K. pneumoniae-pulsed LCs over media in subjects 1, 2, and 4 (23.7%, 9.35%, and 10.17%). The increased IFN-γ secretion (>5%) was observed in CD8+ T cells in subjects 1, 2, and 5 (24.6%, 12.26% and 8.33%). Tetanus toxoid-pulsed LCs induced increased IFN-γ secretion (>5%) by both CD4+ and CD8+ T cells only in subject 1 (15.9% and 20%).
Discussion
The activation and maturation of APCs are essential for initiating immune response and influencing the width, magnitude, and type of the T-cell responses. 5 LCs as a special subset of APCs reside in the epidermis and mucous membrane. The induction of activation and maturation of LCs is important for efficiency of intradermally inoculated vaccines. To investigate the potentiality of K. pneumoniae as an adjuvant in subcutaneous vaccines, we evaluated the impact of K. pneumoniae on the maturation of LCs by examining the expression of HLA-DR, CD86, CD40, and CD80. The results showed K. pneumoniae induced significant increase in the expression of HLA-DR and CD86 but not CD40 and CD80, which is similar to as well as different from those reported by others. Evrard et al. 6 demonstrated that ultraviolet (UV)-killed K. pneumoniae increased significantly the expression of CD86 on DCs. Spisek et al. 7 examined the effect of Biostim®, K. pneumoniae glycoprotein on phenotypic changes of DCs and they found Biostim led to increasing expression of CD80, CD86, CD40, and HLA-DR on DCs. But for monocytes, Biostim significantly enhanced only the expression of HLA-DR and CD80, 8 but not CD40 and CD86. The reason for the differences between our results and other results may be the different types of cells, though they all are APCs, and the different stimulants though they all are from K. pneumoniae. Although no significantly increased expression of CD40 and CD80 was demonstrated, our data showed the increased expression of CD40 in four subjects among all the five examined subjects and the increased expression of CD80 in all the five examined subjects.
The activation of naïve T cells induced by efficacious adjuvants was followed by their proliferation.9 –11 Spisek et al. 7 demonstrated that Biostim-treated DC was able to induce the proliferation of allogeneic CD4+ T cells and Biostim-loaded DC induced more T-cell proliferation than immature DC. Our results showed K. pneumoniae-treated LCs induced significant proliferation of autologous CD4+ and CD8+ T cells, which was similar to that described above. IFN-γ primarily produced by activated T cells and natural killer (NK) is critical for innate and adaptive immunity against viral and some bacterial and protozoal infections.12,13 Braat et al. 14 demonstrated live K. pneumoniae-treated DC induced increased percentage of IFN-γ producing CD4+ T cells. Cabezón et al. 15 determined the percentage of IFN-γ producing allogeneic CD4+ T cells stimulated by heat-killed K. pneumoniae-treated immature DCs and mature DCs. In our study, CD4+ and CD8+ T cells stimulated with K. pneumoniae-treated LCs was demonstrated to secrete significantly more IFN-γ than media using ELISPOT. From these results, we can say K. pneumoniae-treated LCs may induce strong Th1 and CD8+ T cell immune responses, and K. pneumoniae-lysate has the potential to increase cellular immunity, though we do not know the exact immunopotential components. The limitation of our study is the small number of subjects. In the following research, we plan to increase the number of subjects and determine the specific component of K. pneumoniae as a potential adjuvant enhancing cellular immunity in vitro and vivo. In sum, K. pneumoniae has the potential to enhance epidermal cellular immunity and may act as a potential adjuvant in intradermal vaccines designed to enhance cellular immunity.
Footnotes
Declaration of conflicting interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
This work was supported by the National Natural Science Foundation of China (No. 81472439).